
Abstract
Genetic variation of cultivated tomato can be enhanced by induced mutagenesis. Multivariate analysis based on 10 important fruit yield components grouped 21 tomato genotypes into 10 clusters indicating the presence of ample diversity among the genotypes. The genotypes Alisa Craig Aft and CLN-B showed best performance. Seven doses of gamma radiation was employed by imposing 100 Gy–700 Gy radiation with 100 Gy interval to the dry seeds of 5 diverse genotypes selected from different clusters which included the best performers Alisa Craig Aft, CLN-B, two wild relatives Solanum pimpinellifolium L., Solanum lycopersicum var. cerasiformae (Dunal) Spooner and an induced mutant Pmut-5. Seedling emergence and shoot length of these genotypes were recorded to determine the median lethal dose (LD50) and median growth reduction dose (GR50). Probit analysis suggested linear decrease in seed germination and shoot length with the increase in the dose of gamma irradiation for all the five genotypes. The LD50 and GR50 values varied widely among the genotypes, being very high for the wild relatives S. lycopersicum var. cerasiformae (298 Gy and 300 Gy) and S. pimpinellifolium (288 Gy and 295 Gy) followed by 240 Gy 251 Gy for Pmut-5, 179 Gy and 186 Gy for Alisa Craig Aft and 173 Gy 182 Gy for CLN-B. The study suggested the optimum gamma radiation dose ranged between 150 and 300 Gy. This optimum dose can be used to induce a higher rate of effective mutation creating maximum favorable genetic diversity with little biological damage in M2 and succeeding generations of tomato.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Tomato (Solanum lycopersicum L.) is considered one of the most widely cultivated vegetables in the world. It ranks second after potato in terms of consumption and contribute a great deal to the agro-based industries globally. Tomato production has increased worldwide due to its nutritive value and diverse utility as raw, cooked and processed food (Chaudhary et al. 2019). However, genetic diversity of cultivated tomato is low as a consequence of inbreeding during domestication (Foolad 2007; Henareh et al. 2015). Mutations occurring spontaneously results in heritable variation. However, frequency of natural mutation is very low which varies from 10−5 to 10−8 per loci in higher plants (Jiang and Ramachandran 2010). Induced mutagenesis utilizing physical and chemical mutagens create new genetic variation and mutants with desirable characters can be isolated (Mba et al. 2010; Wolabu and Tadege 2016). Induced mutagenesis acts as a viable alternative to conventional breeding methods in induction of desirable changes in various traits without altering the overall superior agronomic base. Mutagenesis maintains the genetic status quo and does not involve introduction of alien genes, unlike in genetically modified organisms (Gruère and Rosegrant, 2008; Davison and Ammann 2017).
Gamma irradiation induces mutation majorly by means structural abberations of chromosome in the form of deletion and translocation (Shu et al. 2012). Gamma rays have been favored over other mutagenic agents because of its easy availability, relatively higher degree of plant tissue penetration, reproducibility and greater mutation frequency (Matsukura et al. 2007; Gruère and Rosegrant 2008; Laskar et al. 2018). Different studies suggested it as the most effective electromagnetic radiation for improvement of different crops including tomato (Norfadzrin et al. 2007; Sikder et al. 2013; Im et al. 2018). Gamma irradiation has been used to extend the shelf life as well as improve some quality attributes of tomato (Desai and Joshi 2018; Munir et al. 2018; Loro et al. 2018; Gyimah et al. 2020). Induced mutagenesis along with innovative breeding approaches like speed breeding and single seed descent selection method can develop cultivars with desired characters in a relatively shorter breeding cycle (Watson et al. 2018; Chiurugwi et al. 2019). Success of induced mutagenesis is dependent upon the use of optimum dose of mutagenic agents (Ernest et al. 2020; Hazra et al. 2021). Optimum dose of mutagen is estimated by calculating the median lethal (LD50) dose, which kills 50% of the total population, and median growth reduction (GR50) dose, which reduces growth by 50% (Gruère and Rosegrant 2008; Mudibu et al. 2012). The principle of dose optimization underlies in the concept that lower doses of mutagenic treatment do not impact the genome significantly as a result of which no change in the phenotype is expected while, higher doses increase the mutation frequency, but the chances of obtaining desirable and utilizable mutants decreases. This study is aimed to assess the radio-sensitivity of five divergent tomato genotypes for gamma irradiation with respect to median lethal and median growth reduction dose for further mutation breeding programs.
2 Materials and methods
Genetic materials for the commencement of the investigation comprised of 21 genotypes of tomato consisting of elite variety (Berika, Ratan, H-24, Alisa Craig), adaptable genotype of West Bengal (Patharkutchi), breeding line (BCT-53,CLN-B, BCT-110,BCT-82, EC 620,176), near isogenic lines of different spontaneous mutant (Alisa CraigAft, Alisa Craighp-1, Alisa Craig ogc, Alisa Craig fulgens), other spontaneous mutant (BCT-111rin,BCT 115 dg), induced mutant (Pmut-5) and four wild relatives (Solanum pimpinellifolium L., Solanum chilense (Dunal) Reiche, Solanum lycopersicum var. cerasiformae (Duran) Spooner, Solanum peruvianum L.) maintained at the Department of Vegetable Science, Bidhan Chandra Krishi Viswavidyalaya, West Bengal, India. These genotypes were evaluated in 2018–19 following randomized block design with 3 replications at Central Research Station, Bidhan Chandra Krishi Viswavidyalaya lying at 23oN latitude, 89E longitude and at 9.75 m elevation above mean sea level in field condition during autumn–winter season (October to March) under the average day and night temperature ranging between 24.8 to 33.4 °C and 10.2–25.1 °C for 10 fruit yield components and yield viz., plant height (cm), days to first flower, flower cluster per plant, flower per cluster, fruits per plant, fruit weight (g), pericarp thickness (mm), locule number/fruit, seeds/fruit and fruit yield per plant(Kg). Each genotype was grown in 2 rows of 6.0 m long with a spacing of 70 × 70 cm following all recommended agronomic practices. Observations on 10 characters were recorded on 5 randomly selected plants of each entry in a replication.
Five widely divergent genotypes selected from different clusters to study the radio-sensitivity for gamma irradiation were Solanum pimpinellifolium and S. lycopersicum var. cerasiformae (wild relatives), best performing lines based on the 10 quantitative characters studied Alisa Craig Aft (isogenic line of Alisa Craig with introgressed Anthocyanin fruit (Aft) gene) and CLN-B (heat tolerant breeding line from AVRDC, Taiwan) and induced mutant Pmut-5 (dark green fruit mutant isolated from Patharkuchi by 150 Gy gamma irradiation). Dry seeds (9% moisture content) of these five genotypes were irradiated with 7 doses of gamma irradiation viz., 100 Gy, 200 Gy, 300 Gy, 400 Gy, 500 Gy, 600 Gy and 700 Gy at Regional Nuclear Agricultural Research Centre, Bidhan Chandra Krishi Viswavidyalaya, Mohanpur, in a Cobalt-60 Gamma cell 5000 Irradiator, Board of Radiation and Isotope Technology, Mumbai, India, at 40.9 Gy∙min−1. Dosimetry was carried out by cerric-cerrous dosimeters calibrated with Fricke’s dosimeter. The M1 seeds (seeds referred to after irradiation) were sown in finely prepared nursery bed. The soil of the seed bed was loam in texture and constituted sufficient organic matter (0.78%). For this reason, after light pre-sowing sprinkling of water, it retained sufficient moisture and light sprinkling of water twice was needed during the course of the experiment. The irradiated seeds were sown in three replications following Randomized Block Design with 100 seeds per replication. Data on seedling emergence and shoot length was taken after 15 days of sowing.
Statistical analysis The data emanated from the evaluation of 21 genotypes were subjected to analysis of variance (Gomez and Gomez, 1984). Genetic divergence among the genotypes was determined by the Mahalanobis’ generalized distance (Mahalanobis 1936) as per Rao (1952) using Genres software version 7.01. Arc-sin transformation was performed for variation in seedling emergence of the irradiated seeds. The data were subjected to analysis of variance and the mean value for seedling emergence and shoot length due to different gamma irradiation dose were subjected to Tuckey’s test using STAR software (Statistical Tool for Agricultural Research, R-packages, ver. 1.5 STAR 2.0.1, International Rice Research Institute, Los Banos, Phillippines). The median lethal dose LD50 and median growth reduction dose GR50 were calculated from survival rate of the seedlings using probit analysis (a type of regression analysis used for the analysis of binomial response variables and transforms the sigmoid dose–response curve to a straight line) according to the equation proposed by Busvine (1971).
3 Results
Analysis of variance suggested significant variation in the 21 genotypes for all the 10 quantitative characters (Table 1). Based on the tomato descriptors and yield per plant, the genotypes Alisa Craig Aft, CLN-B were best performing. The genotypes could be grouped into 10 clusters (Table 2) as per D2 analysis and biological distance depicted by dendrogram (Fig. 1). It suggested the presence of ample divergence among the genotypes. Cluster I and cluster IX accommodated three genotypes each; single genotype came in cluster X while two genotypes each was grouped in the 7 other clusters. Minimum intra-cluster distance was recorded in cluster II (5.903) followed by cluster III (6.532) while it was zero for cluster X being mono-genetic cluster (Fig. 2). Maximum intra-cluster distance was recorded in cluster IX (99.087) followed by cluster I (64.908). Inter-cluster distance was minimum between cluster II and cluster V (8.3), followed by 10.4 between cluster III and cluster V and 10.9 between cluster IV and cluster V. Maximum inter-cluster distance was recorded between cluster IX and cluster X (163), followed by 162.5 between cluster IV and cluster IX and 160.2 between cluster III and cluster IX. The inter-cluster distance between cluster IX and the other clusters were also very high. Of the 10 quantitative characters recorded in the present investigation, fruits per plant contributed the maximum of 34.3% towards total divergence (Fig. 3) followed by fruit yield per plant (31.4%), fruit weight (16.0%) and seeds/fruit (10.0%). Five divergent genotypes which included the best performers Alisa Craig Aft and CLN-B, wild relatives S. pimpinellifolium and Solanum lycopersicum var. cerasiformae and the induced mutant Pmut-5 belonging to 5 different clusters and having conspicuously different genetic background were employed for the study of genotypic response towards median lethal dose (LD50) for gamma irradiation.

Dendrogram depicting biological distance of 21 genotypes

Inter cluster and intra cluster distance (not to scale)

Percentage contribution of the characters towards divergence
Degree of hindrance and tolerance to gamma irradiation for seedling emergence and shoot length of the five genotypes was documented in the M1 generation. Analysis of variance (Table 3) depicted significant difference for seedling emergence and shoot length of the five genotypes due to exposure of the seeds to gamma irradiation. Mean values for both the parameters of the genotypes indicated no beneficial effects of different gamma radiation doses on growth of the tomato genotypes. However, a significant decrease in seedling emergence and shoot length with the increase in the dose was recorded (Table 4; Fig. 4).

Effect of gamma radiation on seedling emergence in M1 generation. Lines labelled with different lowercase letters indicate significantly different shoot length (cm) across the various gamma radiation doses. Bars labelled with different uppercase letters are similarly significantly different for seedling emergence percentage according to Tukey’s honest significant difference test. The significance level of seedling emergence (%) for all the five varieties being similar at each mutagen dose, a single upper case letter has been provided for each bunch of columns
Probit analysis (Table 5; Fig. 5) suggested linear decrease in seedling emergence with the increase in the dose of gamma irradiation for all the five genotypes. The median lethal dose (LD50) varied markedly with the genotypes. The highest LD50 value was recorded for the wild relative S. lycopersicum var. cerasiformae (298 Gy) which was closely followed by the other wild relative S. pimpinellifolium (288 Gy), Pmut-5 (240 Gy), Alisa Craig Aft (179 Gy) and CLN-B (173 Gy). The R2 or coefficient of determination was very high for the genotypes being 0.908, 0.945, 0.908, 0.919 and 0.942 for Solanum pimpinellifolium, S. lycopersicum var. cerasiformae, Alisa Craig Aft, CLN-B and Pmut-5, respectively.

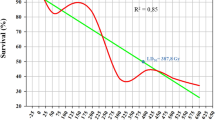
Probit analysis on effect of gamma radiation on seedling emergence in M1 generation. The point of intersection of horizontal and vertical lines indicate the logarithmic value of the median lethal dose, and by taking the antilog of that value the median lethal dose is determined
Probit analysis for shoot length (Table 6; Fig. 6) indicated similar trend as observed in probit analysis for seedling emergence. The R2 values for the genotypes were recorded to be very high being 0.9293, 0.9565, 0.9392, 0.9483 and 0.9162 for S. pimpinellifolium, S. lycopersicum var. cerasiformae, Alisa Craig Aft, CLN-B and Pmut-5, respectively. Similar to seedling emergence the highest GR50 value was observed for S. lycopersicum var. cerasiformae (300 Gy) followed by S. pimpinellifolium (295 Gy), Pmut-5 (251 Gy), Alisa Craig Aft (186 Gy) and CLN-B (182 Gy).

Probit analysis on effect of gamma radiation on shoot length in M1 generation. The point of intersection of horizontal and vertical lines indicate the logarithmic value of the median growth reduction dose, and by taking the antilog of that value the median growth reduction dose is determined
4 Discussion
Significant variation among the genotypes for all the quantitative characters and subsequently grouping of the genotypes in 10 clusters indicated wide divergence among the genotypes which was the prerequisite for the present study. The biological distance among the genotypes and their clustering pattern suggested no parallelism between genetic diversity and geographical origin which was also suggested in earlier reports (Kumar et al. 2013; Prakash et al. 2019; Debnath et al. 2020). Grouping of the genotypes of the same geographical origin into different clusters might be due to change in certain characters as a result of natural or artificial selection as suggested by Narayan et al. (2018). Natural or artificial selection and genetic drift resulted in creation of more genetic diversity rather than genetic distance (Meena and Bahadur 2015). The characters like fruits per plant and fruit yield per plant contributing maximum towards the total divergence may offer a good scope of improvement through selection as suggested by Anuradha et al. (2020) in tomato (S. lycopersicum L.) and Kumar et al. (2020) in okra (Abelmoschus esculentus L. Monech). Laskar et al. (2018) studying in lentil (Lens culinaris Medik) suggested that such characters contributing towards divergence can be improved and evaluated through induced mutagenesis.
Decrease in seedling emergence and shoot length with the increase in mutagen dose was documented in all the five genotypes indicating negative effect of gamma irradiation on seedling emergence as recorded earlier by Norfadzrin et al. (2007). The inhibitory effect on seedling emergence and growth parameters as the result of exposure to gamma irradiation might be due to several reasons as gamma rays imparts highest photon energy and has deep penetrating properties. It results in severe alterations including production of reactive oxygen species which interacts with DNA and other cellular components resulting in functional changes in the overall metabolic activities of the exposed seed materials (Majeed et al. 2018). Earlier reports on other crops suggested that inhibitory effects of gamma rays on seed germination might be the effect of cytochrome oxidase on respiration as reported in wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.) (Swaminathan et al., 1962), altered enzyme activity and/or block in the formation of cellular DNA causing the plant growth to slow or stop in mungbean [Vigna radiate (L.) Wilczek] (Khan and Goyal 2009; Roslim et al. 2015), nature and extent of chromosomal damage in garden bean (Lablab purpureus L.) (Monica and Seetharaman 2016). The differences in germination among the five genotypes due to different gamma irradiation doses might be the result of the metabolic events that take place during germination differ in their timing among the genotypes as suggested earlier by Norfadzrin et al. (2007).
The probit analysis indicating a linear decrease in seedling emergence and shoot length with the increase in gamma radiation doses was supported by the R2 co-efficient which fitted well for all the five genotypes. Coefficient of determination (R2) is similar to the correlation coefficient which indicates how strong of a linear relationship there is between the two variables. The LD50 and GR50 values for the five genotypes corresponded closely with each other proving the reliability of the experiment.
The pre-requisite to any mutation breeding programme is dose optimization, since radio-sensitivity is species, variety and even genotype specific as reported in ground-nut, Arachis hypogaea L. (Tshilenge-Lukanda et al. 2012) and tomato, Solanum lycopersicon L. (Sikder et al. 2013). Calculation of LD50 and GR50 as a method of determining the optimum dose range has been suggested from several earlier studies (Norfadzrin et al. 2007; Sikder et al. 2013; Hazra et al. 2021).
The median lethal and median growth reduction dose of two wild relatives of tomato viz., Solanum pimpinellifolium and Solanum lycopersicum var. cerasiformae, corresponded closely. This might have happened because biological distance between these two genotypes was low although, they were grouped into different clusters. This result also suggested that the wild relatives were less liable to radiation damage than the cultivated tomato genotypes. However, wide variation was recorded for the LD50 and GR50 values of the cultivated tomato genotypes. LD50 and GR50 values for CLN-B and Alisa Craig Aft were much lesser but close to each other suggesting their high sensitivity to gamma irradiation. The median lethal dose and median growth reduction dose of the induced mutant Pmut-5 was comparatively high being 240 Gy and 250 Gy, respectively. It corresponded closely to median lethal dose for the genotype Patharkuchi (303 Gy) from which it was isolated earlier by gamma irradiation (Sikder et al. 2013).
The median lethal dose (LD50) and median growth reduction dose (GR50) varied widely for the five genotypes indicating differences in radio-sensitivity among the genotypes. The optimum gamma radiation dose ranged between 150 and 300 Gy which can be used for further tomato mutation breeding programme using gamma irradiation for creating maximum favorable genetic diversity in M2 and succeeding generations and isolate desirable and utilizable mutants of tomato.
Determination of optimum mutagen dose is critical for any mutation breeding program to isolate mutants with desired agronomic traits. Results from the present investigation suggested differential radio-sensitivity among the five diverse genotypes since the LD50 and GR50 values varied widely between them. Based on the radio-sensitivity results the optimum dose range for gamma irradiation for tomato was between 150 and 300 Gy. Since diverse genotypes were taken into consideration during this investigation this optimum range of gamma irradiation dose might hold good for most of the tomato genotypes. The optimum dose for gamma irradiation determined in this investigation could be useful in future tomato mutation breeding program to isolate mutants with improved agronomic traits.
References
Anuradha B, Reddy PSKR, Sudini H, Geetha A (2020) Genetic divergence for yield and yield attributes in tomato (Solanum lycopersicum L.). Green Farming 11:293–298
Busvine JR (1971) A critical review of the techniques for testing insecticides, 2nd edn. Commonwealth Institute of Entomology, London
Chaudhary J, Alisha A, Bhatt V, Chandanshive S, Kumar N, Mir Z, Kumar A, Yadav SK, Shivaraj SM, Sonah H, Deshmukh R (2019) Mutation breeding in tomato: advances, applicability and challenges. Plants 8:128. https://doi.org/10.3390/plants8050128
Chiurugwi T, Kemp S, Powell W, Hickey LT (2019) Speed breeding orphan crops. Theor Appl Genet 132:607–616
Davison J, Ammann K (2017) New GMO regulations for old: determining a new future for EU crop biotechnology. GM Crops Food 8:13–34
Debnath A, Kumar R, Prasad SK, Sharma N, Kushwah JK (2020) Multivariate analysis for genetic diversity estimation among tomato (Solanum lycopersicum L.) Genotypes. Eco Env Cons 26:1208–1211
Desai DB, Joshi DC (2018) Effect of gamma irradiation on physico-chemical properties and shelf life of tomato (Solanum lycopersicum cv Anand tomato-3) Int J Chem Stud 6: 63–71
Ernest FP, Noëlle MAH, Godswill NN, Thiruvengadam M, Simon OA, Bille NH, Martin BJ, Rebezov M, Shariati MA (2020) Radiosensitivity of two varieties of watermelon (Citrullus lanatus) to different doses of gamma irradiation. Braz J Bot 43:897–905
Foolad MR (2007) Genome mapping and molecular breeding of tomato. Int J Plant Genomics 2007:1–52
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research, 2nd edn. John wiley and sons, NewYork
Gruère GP, Rosegrant MW (2008) Assessing the implementation effects of the biosafety protocol’s proposed stringent information requirements for genetically modified commodities in countries of the Asia Pacific economic cooperation. Rev Agric Econ 30:214–232
Gyimah LA, Amoatey HM, Boatin R, Appiah V, Odai BT (2020) The impact of gamma irradiation and storage on the physicochemical properties of tomato fruits in Ghana. Food Qual Saf 4:151–157
Hazra S, Gorai S, Kumar VU, Bhattacharya S, Maji A, Jambhulkar S, Ali N, Chattopadhyay A (2021) Optimization of gamma radiation dose for induction of mutations in okra. Int J Veg Sci 27:574–584. https://doi.org/10.1080/19315260.2021.1894626
Henareh M, Dursun A, Mandoulakani BA (2015) Genetic diversity in tomato landraces collected from Turkey and Iran revealed by morphological characters. Acta Sci Pol Hortorum Cultus 14:87–96
Im J, Ryu J, Kim WJ, Kim SH, Kang SY, Ha BK (2018) Growth characteristics and biological responses of Korean elite soybean (Glycine max L. Merr.) cultivars exposed to gamma-rays. Plant Breed Biotech 6:109–118
Jiang SY, Ramachandran S (2010) Natural and artificial mutants as valuable resources for functional genomics and molecular breeding. Int J Biol Sci 6:228–251
Khan S, Goyal S (2009) Improvement of mung bean varieties through induced mutations. Afr J Plt Sci 3:174–180
Kumar A, Kumar M, Kumar V, Sharma VR, Chaudhary V (2020) Genetic divergence studies for yield and quality traits in Okra [Abelmoschus esculentus (L.) Moench.]. Int J Agricult Stat Sci 16:355–360
Kumar M, Buckseth T, Thakur MS, Thakur KS (2013) Genetic divergence and cluster analysis in Tomato (Solanum lycopersicum). Progr Agric 13:114–117
Laskar RA, Laskar AA, Raina A, Khan S, Younus H (2018) Induced mutation analysis with biochemical and molecular characterization of high yielding lentil mutant lines. Int J Biol Macromol 109:167–179
Loro AC, Botteon VW, Spoto MHF (2018) Quality parameters of tomatoes submitted to different doses of gamma radiation. Braz J Food Technol 21:e2017168. https://doi.org/10.1590/1981-6723.16817
Mahalanobis PC (1936) On the generalized distance in statistics. Natl Inst Sci India 2:49–55
Majeed A, Muhammad Z, Ullah R, Ali H (2018) Gamma irradiation i: effect on germination and general growth characteristics of plants–a review. Pak J Bot 50:2449–2453
Matsukura C, Yamaguchi I, Inamura M, Ban Y, Kobayashi Y, Yin YG, Saito T, Kuwata C, Imanishi S, Nishimura S (2007) Generation of gamma irradiation-induced mutant lines of the miniature tomato (Solanum lycopersicum L.) cultivar ‘Micro-Tom.’ Plant Biotechnol 24:39–44
Mba C, Afza R, Bado S, Jain SM (2010) Induced mutagenesis in plants using physical and chemical agents. In: Davey MR, Anthony P (eds) Plant Cell Culture: Essential Methods. John Wiley and Sons, Chichester, UK
Meena OP, Bahadur V (2015) Breeding potential of indeterminate tomato (Solanum lycopersicum L.) accessions using D2 analysis. SABRAO J Breed Genet 47:49–59
Monica S, Seetharaman N (2016) Effect of gamma irradiation and ethyl methane sulphonate (EMS) mutagenesis in early generation of garden bean [L. purpurens (L.) Sweet var. typicus]. Int J Adv Sci Tech Res 6:398–410
Mudibu J, Nkongolo KCK, Kalonji-Mbuyi A, Kizungu VR (2012) Effect of gamma irradiation on morpho-agronomic characteristics of soybeans (Glycine max L.). Am J Plant Sci 3:331–337
Munir N, Manzoor A, Haq R, Naz S (2018) Influence of gamma irradiation on shelf life and proximate analysis of fresh tomatoes (Solanum lycopersicum). J Anim Plant Sci 28:1385–1390
Narayan R, Mer MS, Singh DB, Jeena AS, Kishor A (2018) Genetic divergence in tomato (Solanum lycopersicum L.) based on morpho-chemical traits under Polyhouse in Uttarakhand. Biotech Today: an Int J Biol Sci 8:80–84
Norfadzrin F, Ahmed OH, Shaharudin S, Rahman DA (2007) A preliminary study on gamma radiosensitivity of tomato (Lycopersicon esculentum) and okra (Abelmoschus esculentus). Int J Agric Res 2:620–625
Prakash O, Choudhary S, Kumar S, Godara A (2019) Genetic divergence studies in tomato (Solanum lycopersicum L.). J Pharm Phyto 8:4486–4488
Rao CR (1952) Advanced statistical methods in biometrical research. John Wiley and Sons Inc, New York, USA
Roslim DI, Herman A, Fiatin I (2015) Lethal dose 50 (LD50) of mungbean (Vigna radiata L. Wilczek) cultivar Kampar. Soc. Advance Breed. Rsch Asia Oceania J Breed Genet 47:510–516
Shu QY, Forster BP, Nakagawa H (eds) (2012) Plant mutation breeding and biotechnology. CABI, Wallingford. https://doi.org/10.1079/9781780640853.0000
Sikder S, Biswas P, Hazra P, Akhter S, Chattopadhyay A, Badigannavar AM, D’Souza SF (2013) Induction of mutation in tomato (Solanum lycopersicon L.) by gamma irradiation, EMS and their combinations. Indian J Genet 73:392–399
Swaminathan MS, Chopra VL, Bhaskaran S (1962) Chromosome aberrations and the frequency and spectrum of mutations induced by ethyl methane sulphonate in barley and wheat. Indian J Genet 22:192–207
Tshilenge-Lukanda L (2012) Radio-sensitivity of some groundnut (Arachis hypogaea L.) genotypes to gamma irradiation: indices for use as improvement. Br Biotechnol J 2:169–178. https://doi.org/10.9734/BBJ/2012/1459
Watson A, Ghosh S, Williams MJ, Cuddy WS, Simmonds J, Rey MD, Hatta MAM, Hinchli A, Steed A, Reynolds D (2018) Speed breeding is a powerful tool to accelerate crop research and breeding. Nat Plants 4:23–29
Wolabu TW, Tadege M (2016) Photoperiod response and floral transition in sorghum. Plant Signal Behav 11:e1261232
Acknowledgements
We thank the Regional Nuclear Agricultural Research Centre (RNARC), Bidhan Chandra Krishi Viswavidyalaya, Mohanpur, West Bengal, India, for letting us use the facility for this investigation.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
All authors contributed significantly towards the final make-up of the paper.
Conceptualisation (PH and SH); Data curation (SH and SB); Formal analysis (SG and SB); Investigation and methodology (SH and SG); Supervision (PH and AC); Writing-original draft (SH and PH); Writing-reviewing and editing (AM and AC).
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Human or Animal Rights
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hazra, S., Gorai, S., Bhattacharya, S. et al. Radio-sensitivity of diverse tomato genotypes with respect to optimization of gamma irradiation dose. Braz. J. Bot 45, 917–927 (2022). https://doi.org/10.1007/s40415-022-00823-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40415-022-00823-2