
Abstract
Both the impact of habitat modification on the food preferences of species and its impact on ecosystem functioning are poorly understood. In this study, we analyzed food selection by dung beetles in 80 tropical forest fragments and their adjacent cattle pastures in the Los Tuxtlas Biosphere Reserve, Mexico. Ten pitfall traps were placed at each site, half baited with human dung and the other half with fish carrion. We assessed dung beetle food selection and classified any specialization in resource use quantitatively using a multinomial classification model. We collected 15,445 beetles belonging to 42 species, 8747 beetles (38 species) in forest fragments and 6698 beetles (29 species) in cattle pastures. Twenty-five species were present in both habitats. Of all the beetles captured, 76% were caught in dung traps (11,727 individuals) and 24% in carrion traps (3718 individuals). We found 21 species of dung specialists, 7 carrion specialists, 8 generalists, and 6 species too rare to classify. The bait most frequently selected by beetles in this study was dung in both forests and pastures. Specialists tended to remain specialists in both habitats, while generalists tended to change their selection of bait type depending on the habitat. In summary, our results show that replacing forests with cattle pastures modifies the patterns of resource selection by dung beetles and this could affect ecosystem functioning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The diversification of dung beetles on Earth seems to have followed the increase in mammal dung types during the Cenozoic, which would have promoted their high degree of specialization as coprophages (Hanski & Cambefort 1991). However, to feed and reproduce, dung beetles use not only excrement from different mammals and other vertebrates (such as monkeys, tapirs, rodents, tortoises, and birds) but also several other resources such as carrion, eggs decaying, rotten fruit, leaf litter, fungi, and even live animals (Halffter & Matthews 1966, Gill 1991, Pfrommer & Krell 2004). In Neotropical forests, coprophagy is the predominant feeding habit of dung beetles, followed by necrophagy (Halffter & Matthews 1966, Gill 1991, Scholtz & Kryger 2009).
Dung beetles are a diverse and abundant group of insects, with up to 60 species and more than 2000 individuals per hectare (Estrada & Coates-Estrada 2002). They perform many important ecological functions in both natural areas and those modified by human activity, particularly cattle pastures (Nichols et al 2008). One of the main ecosystem services provided by these insects is the removal of excrement, an activity that contributes to nutrient recycling (incorporating nitrogen and phosphorus into the soil) and accelerates mineralization (Nichols et al 2008). Other ecosystem services provided by dung beetles include the aeration of the upper soil layer by large species, the control of parasites such as cattle worms, protozoa and the pest flies that grow on dung (Nichols & Gómez 2014), and secondary seed dispersal that decreases seed predation and contributes to forest regeneration (Andressen 2002).
The rapid loss and degradation of forests throughout the tropics has led to the expansion of fragmented landscapes immersed in agricultural systems, where the long-term persistence of biodiversity may be endangered (Melo et al 2013). Forest fragmentation and matrix homogenization are considered the main causes of species loss and change in the structure and composition of biological communities (Fahrig et al 2011). These changes are also reflected in the structure of the feeding guild and the food chain, both of which have direct effects on many of the ecological processes that are key to ecosystem functioning, such as pollination, decomposition, and predation (Hooper et al 2005). The impact of habitat change on species’ food preference and its impact on ecosystem functioning are still poorly understood.
For several decades, dung beetles have been used to evaluate the effects of habitat loss and fragmentation (Howden & Nealis 1975, Nichols & Gardner 2011). Most studies have focused on evaluating how these changes affect the diversity and structure of the community, but little is known about the impact of the modification of tropical forest on the food preferences of dung beetles and the potential effects of these preferences on ecosystem functioning (Nichols et al 2008). In this study, we asked whether land use conversion from tropical forest to cattle pastures influences food selection by dung beetles. This was assessed by quantifying how many dung beetles fell into pitfall traps containing two different types of bait, human dung and rotten fish. We evaluated how constant or changeable food selection specificity by dung beetles was in forest fragments and cattle pastures. Given that carrion is more abundant in forest and dung is more abundant in pastures, we expected differences in beetle abundance by bait and habitat, with more beetles in carrion traps set in the forests and more beetles in dung traps set in the pastures. Also, we predicted that dung beetles would have the plasticity required to switch food preference as resource availability varied, represented by the two habitats studied. If this is correct, then species categorized as carrion specialists or generalists in the forest would be recognized as dung specialists in pastures.
Material and Methods
Study site
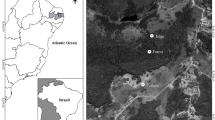
The region of Los Tuxtlas is located in the state of Veracruz in central-eastern Mexico, on the coastal plain of the Gulf of Mexico. It is delimited by a mountain range located between latitudes 18°05′N–18°45′N and longitudes 94°35′W–95°30′W (Fig 1). This plain is 3300 km2 in area and was created by the activity of about 300 volcanoes and sedimentation. There is a strong elevation gradient that rises from sea level to 1680 m above sea level (asl) at the top of its two volcanoes (San Martin Tuxtla, Santa Marta). Los Tuxtlas represents the northern limit of tropical forest in Mexico (Rzedowski 1963). The climate is warm and humid, with a mean annual temperature of 25°C and annual rainfall between 3000 and 4600 mm (Soto 2004).

Location of the Los Tuxtlas Biosphere Reserve, Mexico. The dots represent sampling sites. Each sampled site corresponds to a fragment of forest and a pasture adjacent.
The study was conducted in the Los Tuxtlas Biosphere Reserve (∼1552 km2), which has been highly impacted by human activities (Dirzo & García 1992, Flint Hughes et al 2000). The original dominant vegetation type (below 700 m asl) was tropical rainforest, but the area has been heavily deforested and fragmented since the 1970s, and the remaining rainforest is surrounded by a matrix of pastures and crops. In 2004, only 17.2% of the original vegetation remained in the region. By that time, the land use was cattle pasture (72.7%), secondary vegetation (4.2%), crops (4.6%; sugarcane, tobacco, banana, avocado, coffee, and maize), and human settlements (1.3%) (Castillo-Campos & Laborde 2004).
Beetle surveys
We sampled beetle ensembles (sensu Fauth et al 1996) from July to August 2003 during the rainy season, which is the most active season for dung beetles due to temperature and humidity conditions (Gill 1991). Within the reserve, we collected beetles from 80 forest patches (size 0.92–244 ha) and 80 adjacent pastures. All sampling sites were located within the two coastal landscapes of the reserve (∼50 km2), from 0 to 200 m asl (Fig 1). Ten pitfall traps were set up along a transect at each site, for a total of 1600 traps in the entire study. Traps were separated by 50 m, which has been shown to prevent any interference between effective sampling areas for this group and to provide a good sample of the dung beetle species in a given area (Larsen & Forsyth 2005). We baited half of the traps with decomposing fish (∼50 g) and the other half with ∼30 g of human dung. We exposed the traps once, for 48 h, before collecting the beetles. For beetle identification, we used keys and descriptions of the Neotropical genera and species (e.g., Morón 1979, 2003, Génier 1996, 2009, Kohlmann & Solís 1997, 2006, Rivera-Cervantes & Halffter 1999, Delgado & Kohlmann 2007, Edmonds & Zidek 2010, 2012, González-Alvarado & Vaz-De-Mello 2014) and deposited specimens in the reference collection housed in Federico Escobar’s laboratory at the Instituto de Ecología A. C., in Xalapa, Veracruz, Mexico.
Data analysis
Species were classified based on their bait selection using the multinomial classification model proposed by Chazdon et al (2011). This analysis is based on the relative abundance of individuals, correcting for the fact that sampling tends to overestimate common species and underestimate rare ones. The method permits a robust statistical classification of habit (specialists and generalists), without excluding rare species a priori (Chazdon et al 2011). Given our design and interests, this analysis allowed us to place species into the following four categories: dung specialists, carrion specialists, generalists, and beetles too rare to classify. First, we performed a general multinomial classification model without taking into account habitat type, and then, we ran the analysis for the forest patches and pastures independently. All analyses were run in CLAM software (Chao & Lin 2011). An important parameter of the multinomial model is K, which refers to the cutoff point for classifying species according to their feeding preferences (Chazdon et al 2011). The analytical procedure performs one-sided statistical tests to classify species at a specified significance level, P. Thus K and P are inputs to the multinomial model. We used a cutoff point of K = 0.5 for a simple majority rule or liberal threshold and a conservative P value of 0.005. We used a conservative approach because our goal was to analyze the whole community, rather than a particular species (Chazdon et al 2011).
To determine whether abundance was a function of habitat and bait, we applied a two-way analysis of variance using a generalized linear model, assuming a Poisson distribution of errors (link function = log; Crawley 2007). Additionally, to test if the preferences of shared species between habitats changed, we used a Wilcoxon signed-rank test on the results of the classification analysis.
Finally, we reviewed the literature to compare the feeding preferences we recorded with those reported previously (Halffter & Matthews 1966, Kohlmann 1984, Morón et al 1985, Gill 1991, Edmonds 1994, 2000, Génier 1996, 2009, Zunino & Halffter 1997, Kohlmann & Solís 1997, 2001, 2004, 2006, Solís & Kohlmann 2002, Morón 2003, Edmonds & Zidek 2004, Delgado & Kohlmann 2007, Escobar et al 2008, Price & May 2009).
Results
We collected 15,445 beetles belonging to 42 species (Table 1), 38 species in remnants of tropical rainforest and 29 in cattle pastures. Of the total species, four account for 53% of all individuals captured, three of which were captured in dung—Bdelyropsis newtoni Howden, Onthophagus batesi Howden & Cartwright, and Uroxys boneti Pereira & Halffter—and the other in carrion—Deltochilum pseudoparilae Paulian. Twenty-one species specialized on dung, seven on carrion, eight were generalists, and six species were too rare to classify (Table 1).
We found a significant effect of habitat on the number of beetles captured with each type of bait (two-way ANOVA, interaction effect χ 2 = 11.698, df = 1, P = 0.0006). There were differences in the number of beetles captured in carrion between forest fragments (mean ± 95% CI = 35.4 ± 19.42) and cattle pastures (12.4 ± 4.15), while the number of beetles captured in dung was similar in both habitats (forest = 78.0 ± 35.44; cattle pasture = 71.5 ± 33.23).
In forest fragments, two thirds of the beetles were captured in dung traps (6014 of 8747), and of these, 18 were dung specialists, 4 were carrion specialists, 5 were generalists, and 11 were too rare to classify. In cattle pastures, 85% of the beetles were found in dung traps (5713 of 6698), and of these, 3 species were dung specialists, 6 carrion specialists, 9 generalists, and 11 too rare to classify (Table 2). There were fewer dung specialists, more generalists, and fewer carrion specialists in pastures than in forests (Table 2). Only O. batesi was a dung specialist in both habitats. Of the carrion specialists, B. newtoni was only found in forests and three species (Canthidium laetum Harold, Coprophanaeus pluto Harold, and Deltochilum sublaeve Bates) were only found in cattle pastures.
Of the 42 species collected, 25 were present in both habitats (Table 1). By examining this set of species, we found that their food selection changed between habitats (V = 161, p = 0.007). Of all the species shared between habitats, none changed from one specialization to another and no generalist species remained a generalist (Fig 2). Some dung specialists in forest fragments changed to generalists in cattle pastures (Canthon euryscellis Bates, Copris laeviceps Harold, Copris lugubris Boheman, Dichotomius amplicollis Harold) or remained dung specialists (Dichotomius colonicus Say and O. batesi). Carrion specialists in forest fragments remained carrion specialists in pastures (Canthon cyanellus LeConte, Coprophaneus telamon Erichson, D. pseudoparile) and one generalist in forest fragments changed to a carrion specialist in cattle pastures (D. sublaeve).

Food selection by dung beetle species shared between forest fragments and cattle pastures. Line width represents the number of species that changed from their preferred bait to another, ranging from 1 to 4. Numbers correspond to species number for each type of feeding preference according to the CLAM analysis.
Discussion
This study shows that there is a change in bait selection by species shared between forest remnants and cattle pastures. In general, specialist species tended to continue to be specialists in both habitats while generalists tended to modify their selection of bait type between the two habitats, supporting the notion that generalist species in their feeding habits may occupy different habitats, making them more tolerant to the changes in food availability that result from forest modification. Although the availability of food is one of the factors determining the distribution of dung beetles, in Neotropical rainforests, it does not seem to be a limiting factor. Despite the abundance of cattle dung in pastures, many of the beetles that inhabit the forest cannot occupy these environments due to changes in canopy cover (i.e., more light), microclimate (i.e., high temperature), and soil characteristics (i.e., compaction), as reported in various studies (Escobar et al 2008, Barragán et al 2014).
Coprophagy was the most common specialization found in this study, particularly in forest fragments. This is considered the primitive feeding mode in dung beetles and one which has determined their development, behavior, distribution, and morphology (Halffter & Matthews 1966). Some of our dung specialists have only been reported in the literature on dung, C.euryscellis, Canthon femoralis Chevrolat, Canthon subhyalinus Harold, D. colonicus, and Sulcophanaeus chryseicollis Harold (see Table 2). These species were almost exclusively collected on dung in our study. Other species have been reported to exhibit a strong preference for dung, O. batesi, Eurysternus mexicanus Harold, and Dichotomius satanas Harold (Halffter & Matthews 1966, Zunino & Halffter 1997, Kohlmann & Solís 2001, Morón 2003, Escobar et al 2008, Génier 2009). All of our carrion specialist species have previously been collected on carrion, and the following three species were reported with a strong preference for carrion: C. telamon, D. sublaeve, and D. pseudoparile (Halffter & Matthews 1966, Morón 2003, Escobar et al 2008).
Previous studies have shown that even if most dung beetles are opportunistic, dung from omnivorous mammals, such as humans, attracts more species and more individuals (Silva et al 2010, Bogoni & Hernández 2014) and that dung beetles prefer dung from small- and medium-sized mammals (Estrada et al 1993, Larsen et al 2006). Dung beetles select omnivore dung because it is more nutritious than the dung of carnivores and herbivores (Bogoni & Hernández 2014). The amount of dung that beetles eat or use differs according to the type of dung available in a given habitat. For example, they remove about two times more monkey dung (Ateles geoffroyi) than cow dung in Los Tuxtlas (Amézquita & Favila 2010), which could explain the higher beetle abundance in forest fragments compared to pastures and may also explain the increased necrophagy and generalization in cattle pastures.
We only found two species of dung specialist in both pastures and forests, D. colonicus and O. batesi. The first mainly inhabits pastures but is highly invasive as soon as forest is cleared (Morón 2003), and the second was previously found in Los Tuxtlas both in pastures surrounded by tropical forest and edges of those forests (Lobo & Montes de Oca 1994). Although there are more dung specialists species than carrion specialists in both forests and cattle pastures, there were more species of carrion specialists in cattle pastures.
Cadaverine is the main chemical compound that attracts necrophagous dung beetles (Favila et al 2012), and this is why rotten fish is widely used as bait for dung beetles in Neotropical regions (Yamanaka et al 1989). Necrophagy in the Scarabaeinae is considered particularly important in the Neotropics, where the occurrence of dung from large mammals is low (Halffter & Matthews 1966, Gill 1991). More non-dung-feeding dung beetles are found in the Neotropics than in any other region (Scholtz & Kryger 2009). Dung beetles in tropical America have been seen eating the carcasses of vertebrates but also Atta ants, millipedes, and other invertebrates (Gill 1991, Escobar 2003). Canthon, Coprophanaeus, and Deltochilum have many necrophagous species (Scholtz & Kryger 2009). Canthon cyanellus is known for its necrophagous habit (Halffter & Matthews 1966). Other genera that we found in the necrotraps have also been found on carrion, Ateuchus and Deltochilum (Halffter & Matthews 1966). Ateuchus illaesum Harold, classified as a carrion specialist in our study, has also been found on rotten fruit in the Lesser Antilles and on dung in Martinique (Halffter & Matthews 1966).
Resource distribution is much more aggregated in cattle pastures than in forests. Cows produce dung in big patches, up to 30 kg a day of dung per individual (Favila 2014), while howler monkeys (Alouatta palliata) produce an average of only 50 g a day of dung per individual (Estrada et al 1993). Resource aggregation in pastures can make competition more intense, and this could explain the loss of dung specialists and increase in generalist species that may be less sensitive to competition and more tolerant to open environments. Dung beetle flexibility in food selection could have genetic basis, in terms of the beetles’ capacity to adapt to different temperatures, dung availability, and soil conditions (Simmons & Ridsdill-Smith 2011). Some species are only found in pastures, such as Digitonthophagus gazella Fabricius, a species introduced in California and Texas around 1972 to recycle cattle dung more efficiently (Barbero & Guerrero 1992). This species quickly expanded its distribution in tropical America, taking advantage of a novel resource—cattle dung—when cattle were brought to the America during the Spanish Conquest and one that is underexploited by tropical native beetles in pastures because of their harsh environmental conditions (Favila 2014). In our study, D. gazella was not captured in the forest but was found in both carrion and human dung in pastures. However, in a concurrent sampling effort, we searched for beetles on cow dung and found this species to be abundant (more than 100 individuals in 10 cow pats, F. Escobar, data unpublished).
When it comes to making nests and feeding offspring, selection is much stricter than in adult feeding (Philips 2011), and this could explain shifts in preference by the more flexible species. The reproductive rates of some beetles can be affected by the quality of dung they feed on (Giller & Doube 1994). For example, omnivores produce higher-quality dung, so the species that evolved with them usually produce nests with more chambers and more progeny than dung beetles that evolved with big herbivores (Doube 1991). A more direct evaluation of dung beetle food selection could be made in controlled and field experiments, using the dung and carrion that are typically available in these habitats, such as monkey, coati, and cattle dung (Estrada et al 1993), and the carrion of native and invasive invertebrates and vertebrates, such as rats (Whipple & Hoback 2012).
The change in habitat from forests to cattle pastures modifies the microclimate for beetles in terms of humidity, soil temperature, and vegetation density, leading to a shift in their diversity and species composition (Nichols et al 2008). Three quarters of the species were present in both habitats, but only 10 species could be assessed for food preference based on the analysis we used. Only 3 species of dung specialists were found in cattle pastures, compared to 18 species in forests. With the loss of habitat, many species of dung specialists are lost, and dung specialists may be more sensitive to a reduction in or the disappearance of forest at the landscape scale. Dung processing on cattle pastures is then mostly left to generalists and to dung specialists that are mostly specialized on open environments such as pastures.
The impact of species loss on ecosystems is still not completely clear for invertebrates (Bikhofer et al 2015). It has been suggested that eliminating so-called redundant species would have a negative effect on ecosystem functioning, in part because the many of the ecosystems would become less resilient (Rosenfeld 2002) and in part because some of their benefits to humans could be affected, such as carbon sequestration, soil stability, dung degradation, and pest control, when there’s a change in land use (Wolters 2001). If there is more dung available and fewer dung specialists in cattle pastures, the dung could remain on the ground undegraded in cattle pastures longer than in forests. This could lead not only to slower nutrient cycling, which could limit the recovery of forests, but also to bigger fly infestations and the spread of disease among cattle and domesticated animals. The replacement of forest by grazing areas could be modifying the patterns of resource selection by dung beetles with an unknown impact on the functional integrity of the ecosystem.
References
Amézquita S, Favila ME (2010) Removal rates of native and exotic dung by dung beetles (Scarabaeidae: Scarabaeinae) in a fragmented tropical rain forest. Environ Entomol 39:328–336
Andressen E (2002) Dung beetles in a central Amazonian rainforest and their ecological role as secondary seed dispersers. Ecol Entomol 27:257–270
Barbero E, Guerrero Y (1992) Some considerations on the dispersal power of Digitonthophagus gazella (Fabricius 1787) in the new world (Coleoptera, Scarabaeidae, Scarabaeinae). Trop Zool 5:115–120
Barragán F, Moreno CE, Escobar F, Bueno-Villegas J, Halffter G (2014) The impact of grazing on dung beetle diversity depends on both biogeographical and ecological context. J Biogeogr 41:1991–2002
Bikhofer K, Smith HG, Weisser WW, Wolters V, Gossner MM (2015) Land-use effects on the functional distinctness of arthropod communities. Ecography 38:001–012
Bogoni JA, Hernandez MIM (2014) Attractiveness of native mammal’s feces of different trophic guilds to dung beetles (Coleoptera: Scarabaeinae). J Insect Sci 14:1–7
Castillo-Campos G, Laborde J (2004) La vegetación. In: Guevara S, Laborde J, Sánches-Ríos G (eds) Los Tuxtlas. El paisaje de la sierra. Instituto de Ecología, A. C., and European Union, Xalapa, pp 231–265
Chao A, Lin SY (2011) Program CLAM (Classification Method) program and user’s guide. Available from: http://www.purl.oclc.org/clam. Accessed 20 Apr 2015
Chazdon RL, Chao A, Colwell RK, Lin SK, Norden N, Letcher SG, Clark DB, Finegan B, Arroyo JP (2011) A novel statistical method for classifying habitat generalists and specialists. Ecology 92:1332–1343
Crawley MJ (2007) The R book. 2nd ed. John Wiley & Sons. Chichester, p 942
Delgado L, Kohlmann B (2007) Revisión de las especies del genero Uroxys Westwood de México y Guatemala. Folia Entomol Mex 46:1–35
Dirzo R, García MC (1992) Rates of deforestation in Los Tuxtlas, a Neotropical area in Southeast Mexico. Conserv Biol 6:84–90
Doube BM (1991) Dung beetles of southern Africa. In: Hanski I, Cambefort Y (eds) Dung beetle ecology. Princeton University Press, Princeton, pp 133–155
Edmonds WD (1994) Revision of Phanaeus Macleay, a new world genus of Scarabaeinae dung beetles (Coleoptera: Scarabaeidae, Scarabaeinae). Contrib Sci 443:1–105
Edmonds WD (2000) Revision of the Neotropical dung beetle genus Sulcophanaeus (Coleoptera: Scarabeidae: Scarabaeinae). Folia Heyrovskyana Suppl 6:1–60
Edmonds WD, Zidek J (2004) Revision of the Neotropical dung beetle genus Sulcophaphaeus (Coleoptera: Scarabeidae: Scarabaeinae). Folia Heyrovskyana Suppl 11:1–58
Edmonds WD, Zidek J (2010) A taxonomic review of the Neotropical genus Coprophanaeus Olsoufieff, 1924 (Coleoptera: Scarabaeidae, Scarabaeinae). Insecta Mundi 0129:1–111
Edmonds WD, Zidek J (2012) Taxonomy of Phanaeus revisited: revised keys to and comments on species of the new world dung beetle genus Phanaeus MacLeay, 1819 Coleoptera: Scarabaeidae: Scarabaeinae: Phanaeini). Insecta Mundi 0274:1–108
Escobar F (2003) Feeding habits and distributional records of 11 species of Neotropical Scarabaeinae (Coleoptera: Scarabaeidae). Coleopt Bull 57:131–132
Escobar F, Halffter G, Solís A, Halffter V, Navarrete D (2008) Temporal shifts in dung beetle community structure within a protected area of tropical wet forest: a 35-year study and its implications for long-term conservation. J Appl Ecol 45:1584–1592
Estrada A, Coates-Estrada R (2002) Dung beetles in continuous forest, forest fragments and in an agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodivers Conserv 11:1903–1918
Estrada A, Halffter G, Coates-Estrada R, Meritt DA Jr (1993) Dung beetles attracted to mammalian herbivore (Alouatta palliata) and omnivore (Nasua narica) dung in the tropical rain forest of Los Tuxtlas, Mexico. J Trop Ecol 9:45–54
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin JL (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14:101–112
Fauth JE, Bernardo J, Camara M, Resetarits WJ Jr, Van Buskirk J, McCollum SA (1996) Simplifying the jargon of community ecology: a conceptual approach. Am Nat 147:282–286
Favila ME (2014) Historical, biogeographical and ecological factors explain the success of some native dung beetles after the introduction of cattle in Mexico. Pastos 42:161–181
Favila ME, Ortiz-Domínguez M, Chamorro-Florescano I, Cortez-Gallardo V (2012) Comunicación química y comportamiento reproductor de los escarabajos rodadores de estiércol (Scarabaeinae: Scarabaeini): aspectos ecológicos y evolutivos, y sus posibles aplicaciones. In: Rojas JC, Malo EA (eds) Temas selectos en ecología química de insectos. El Colegio de la Frontera Sur, Chiapas, pp 141–164
Flint Hughes R, Kauffman JB, Jaramillo VJ (2000) Ecosystem-scale impacts of deforestation and land use in a humid tropical region of Mexico. Ecol Appl 10:515–527
Génier F (1996) A revision of the Neotropical genus Ontherus. Mem Entomol Soc Can 170:3–170
Génier F (2009) Le genre Eurysternus Dalman, 1824 révision taxonomique et clés de détermination illustrées. Pensoft Series Faunistica 85:1–430
Gill BD (1991) Dung beetles in tropical American forests. In: Hanski I, Cambefort Y (eds) Dung beetle ecology. Princeton University Press, Princeton, pp 211–229
Giller PS, Doube BM (1994) Spatial and temporal co-occurrence of competitors in southern African dung beetle communities. J Anim Ecol 63:629–643
González-Alvarado A, Vaz-de-Mello FZ (2014) Taxonomic review of the subgenus Hybomidium Shipp 1897 (Coleoptera: Scarabaeidae: Scarabaeinae: Deltochilum). Ann Soc Entomol France (NS) 50:431–447
Halffter G, Matthews EG (1966) The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera, Scarabaeidae). Folia Entomol Mex 12–14:3–312
Hanski I, Cambefort Y (1991) Dung beetle ecology. Princeton University Press, Princeton, p 481
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, La Seta H, Ymstad AJS, Andermeer JV, Wardle DA (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
Howden HF, Nealis VG (1975) Effects of clearing in a tropical rain forest on the composition of the coprophagous scarab beetle fauna (Coleoptera). Biotropica 7:77–83
Kohlmann B (1984) Biosistemática de las especies norteamericanas del género Ateuchus (Coleoptera: Scarabaeidae: Scarabaeinae). Folia Entomol Mex 60:3–81
Kohlmann B, Solís A (1997) El género Dichotomius (Coleoptera: Scarabaeidae) en Costa Rica. G Ital Entomol 8:343–382
Kohlmann B, Solís A (2001) El género Onthophagus (Coleoptera: Scarabaeidae) en Costa Rica. G Ital Entomol 49:159–261
Kohlmann B, Solís A (2004) El género Canthidium (Coleoptera: Scarabaeidae) en Costa Rica. G Ital Entomol 52:1–73
Kohlmann B, Solís A (2006) El género Canthidium (Coleoptera: Scarabaeidae) en Norteamerica. G Ital Entomol 53:235–295
Larsen TH, Forsyth A (2005) Trap spacing and transect design for dung beetle biodiversity studies. Biotropica 37:322–325
Larsen TH, Lopera A, Forsyth A (2006) Extreme trophic and habitat specialization by Peruvian dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt Bull 60:315–324
Lobo J, Montes de Oca E (1994) Local distribution and coexistence of Digitonthophagus gazella (Fabricius, 1787) and Onthophagus batesi Howden and Cartwright 1963 (Coleoptera: Scarabaeidae). Elytron 8:117–127
Melo FPL, Arroyo-Rodríguez V, Fahrig L, Martínez-Ramos M, Tabarelli M (2013) On the hope for biodiversity-friendly tropical landscapes. Trends Ecol Evol 28:461–468
Morón MA (1979) Fauna de coleópteros lamelicornios de la estación de biología tropical “Los Tuxtlas”, Veracruz, UNAM. México. An Esc Nac Cienc Biol Mex. Ser Zool 1:375–454
Morón MA (2003) Atlas de los escarabajos de México Coleoptera: Lamellicornia Vol. 11 Familias Scarabaeidae, Trogidae, Passalidae y Lucanidae. Argania editio, Barcelona, p 227
Morón MA, Villalobos FJ, Deloya C (1985) Fauna de coleópteros lamelicornios de Boca del Chajul, Chiapas, México. Folia Entomol Mex 66:57–118
Nichols ES, Gardner TA (2011) Dung beetles as a candidate study taxon in applied biodiversity conservation research. In: Simmons LW, Ridsdill-Smith TJ (eds) Ecology and evolution of dung beetles. Blackwell Publishing, Sussex, pp 267–292
Nichols E, Gómez A (2014) Dung beetles and fecal helminth transmission: patterns, mechanisms and questions. Parasitology 141:614–623
Nichols E, Spector S, Louzada, Larsen JT, Amézquita S, Favila ME (2008) Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol Conser 141:1461–1474
Pfrommer A, Krell FT (2004) Who steals the eggs? Coprophanaeus telamon (Erichson) buries decomposing eggs in western Amazonian rain forest (Coleoptera: Scarabaeidae). Coleopt Bull 58:21–27
Philips TK (2011) The evolutionary history of diversification of dung beetles. In: Simmons LW, Ridsdill-Smith TJ (eds) Ecology and evolution of dung beetles. Blackwell Publishing, Sussex, pp 21–47
Price DL, May ML (2009) Behavioral ecology of Phanaeus dung beetles (Coleoptera: Scarabaeidae): review and new observations. Acta Zool Mex 25:211–238
Rivera-Cervantes LE, Halffter G (1999) Monografía de las especies mexicanas de Canthon del subgenero Glaphyrocanthon. Acta Zool Mex 77:23–150
Rosenfeld JS (2002) Functional redundancy in ecology and conservation. Oikos 98:156–162
Rzedowski J (1963) El extremo boreal del bosque tropical siempre verde en Norteamérica continental. Vegetatio 11:173–198
Scholtz CDA, Kryger U (2009) Evolutionary biology and conservation of dung beetles. Pensoft Publishers, Sofia, p 587
Silva FAB, Costa CMQ, Moura RC, Faria AI (2010) Study of the dung beetle coleoptera (Scarabaeidae) community at two sites: Atlantic forest and clear-cut, Pernambuco, Brazil. Environ Entomol 39:359–367
Simmons LW, Ridsdill-Smith TJ (2011) Reproductive competition and its impact on the evolution and ecology of dung beetles. In: Simmons LW, Ridsdill-Smith TJ (eds) Ecology and evolution of dung beetles. Blackwell Publishing, Sussex, pp 1–20
Solís A, Kohlmann B (2002) El género Canthon (Coleoptera: Scarabaeidae) en Costa Rica. G Ital Entomol 50:1–68
Soto A (2004) El clima. In: Guevara S, Laborde J, Sánches-Ríos G (eds) Los Tuxtlas. El paisaje de la sierra. Instituto de Ecología, A. C., and European Union, Xalapa, pp 195–198
Whipple SD, Hoback WW (2012) A comparison of dung beetle (Coleoptera: Scarabaeidae) attraction to native and exotic mammal dung. Environ Entomol 41:238–244
Wolters V (2001) Biodiversity of soil animals and its function. Eur J Soil Biol 37:221–227
Yamanaka H, Shiomi K, Kikuchi T (1989) Cadaverine as a potential index for decomposition of salmonoid fishes. Food Hyg Saf Sci 30:170–174
Zunino M, Halffter G (1997) Sobre Onthophagus Latreille, 1802 americanos (Coleoptera: Scarabaeidae: Scarabaeinae). Elytron 11:157–178
Acknowledgments
We would like to thank Fernando Escobar Hernández for confirming our dung beetle identifications, Susan G. Letcher for the assistance with the CLAM analysis, Juan José Von Thaden for the help with the maps, Andrés J. Quesada and Bianca Delfosse for the style correction of the manuscript, and two anonymous reviewers for the comments that greatly improved the manuscript. This research was funded by the Consejo Nacional de Ciencia y Tecnología de México (CONACYT scholarship 477922 and CONACYT project 37514-V).
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by Marcelo N Rossi – Unifesp
Rights and permissions
About this article

Cite this article
Bourg, A., Escobar, F., MacGregor-Fors, I. et al. Got Dung? Resource Selection by Dung Beetles in Neotropical Forest Fragments and Cattle Pastures. Neotrop Entomol 45, 490–498 (2016). https://doi.org/10.1007/s13744-016-0397-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-016-0397-7