
Abstract
Purpose of Review
In the modern obesogenic environment, food cues play a crucial role in the development of obesity by disrupting hormone and energy balance mechanisms. Thus, it is critical to understand the neurobiology of feeding behaviors and obesity in the context of ubiquitous food cues. The current paper reviews the physiology of feeding, hormonal regulation of energy balance, and food cue responses and discusses their contributions to obesity.
Recent Findings
Food cues have strong impact on human physiology. Obese individuals have altered food cue-elicited responses in the brain and periphery, overpowering hormone and energy balance regulation. Disrupted homeostasis during food cue exposure leads to continued food intake, unsuccessful weight management, and poor treatment outcomes, which further contributes to obesity epidemic.
Summary
Findings from the review emphasize the crucial role of food cues in obesity epidemic, which necessitates multidimensional approaches to the prevention and treatment of obesity, including psychosocial interventions to reduce food cue reactivity, along with conventional treatment.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Obesity Today
A marked steady increase in the prevalence of obesity has been observed in the end of last century. Population weight status has been tracked by the National Health and Nutrition Examination Survey (NHANES) from the National Center for Health Statistics since 1960, and a sharp increase was noted in the 1970s (Fig. 1) [1]. In 2000, more than two-thirds of the US adult population was found to be overweight or obese [1], reaching an epidemic status [2]. Several risk factors have been identified to promote weight again, including environment and genetic factors [3,4,5,6]. Among them, the modern obesogenic environment has been implicated as one of the main drivers in the epidemic of obesity [7, 8], associated with sedentarism [9, 10] and increased consumption of highly palatable and calorie-rich processed foods [8, 11, 12, 13•]. In Western wealthy societies, food became cheaper, widely available, and offered in large portions [4, 14]. Increased consumption of calorie-dense food unopposed by an equal amount of energy expenditure (physical activity) may be playing a major role to the observed progressive increase in the population’s body weight.

Prevalence of overweight and obesity (BMI > 25), adapted from CDC: “Trends in overweight, obesity, and extreme obesity aged 20–74 years: United States, 1960–1962 through 2009–2010.” Sources: National Center for Health Statistics, National Health Examination Survey, and National Health and Nutrition Examination Survey
Food availability and palatability appears to influence food intake, but it cannot solely explain the obesity epidemic. Not all rich Western countries present with this dramatic increase in the prevalence of obesity [3], and in the USA, obesity is more prevalent in lower social economical classes [7]. Therefore, societal, cultural, biological, and genetic factors must be influencing weight gain. During this same period that the prevalence of obesity increased exponentially, major changes in how food is processed and presented for consumption were observed [4, 15, 16•]. Processed foods have been precisely tailored to meet the optimal taste and presentation, which is usually achieved by the expenses of adding a significant amount of sugar and fat [16•, 17, 18]. This flavorful and appetizing diet may influence our eating habits by decreasing satiety [19, 20•] and increasing desire and craving for food [21]. In addition, food cues are ubiquitous [22], and they appear to work as a reinforcer for less healthier food choices [16•, 23]. These findings suggest that food cues have significant impact on human physiology.
The obesity epidemic thus suggests that the modern obesogenic environment is overpowering the body’s innate ability to regulate food intake and energy balance in the context of ubiquitous food cues. The current paper will review the physiology of feeding and energy balance and neurophysiology of food cue response and discuss their contributions to obesity.
Hormone and Energy Balance
Physiology of Feeding
The feeding process starts even before actual food ingestion. Environmental cues, such as time of the day and food availability, together with homeostatic signals relaying information about the body’s energy status are integrated by the central nervous system (CNS) to regulate eating behavior [24, 25]. Sight and/or smell of a food trigger feeding-related reactions such as increased salivation in preparation for the actual feeding [26, 27]. As food ingestion starts, signals from the oral cavity (food texture and taste) will further influence palatability and desire to eat [28]. Subsequently, mechanical and neural signals from food being swallowed, digested, and absorbed in conjunction with rising circulating levels of nutrients, gastrointestinal peptides, and hormones will be integrated by the brain to help define meal size and when to stop eating (satiety) [29, 30]. These signals can reach the brain both directly, through receptors in the hindbrain and hypothalamus [31], and indirectly via the afferent vagus nerve [32].
Several hormones have been shown to modulate satiety via peripheral nervous system as well as by direct effects in the CNS in both animal and in humans. (See recent reviews for hormonal regulation of feeding [30, 33, 34••]).
Between meals, the interaction of environmental and homeostatic signals plays an important but underappreciated role in defining when and how much to eat [33]. Prior to and in preparation to receive the next meal, a brain-initiated hormonal release, called pre-meal cephalic hormonal release, was observed in both animals [35, 36] and humans [37, 38]. Animals habituated to eat at a certain time of day demonstrate brain-induced release of insulin, GLP-1, and ghrelin 15 min to an hour before actual meal consumption [39]. This pre-meal hormonal release may work as a signal to control hunger prior to eating and may be a mechanism to help limit food intake [33].
While food is being digested and absorbed, feeding-related hormones are released, influencing satiation and termination of food intake. Among them, insulin is the primary hormone which serves to regulate glucose metabolism [40]. Insulin is secreted in response to a meal [41], in particular to glucose ingestion. This post-prandial rise in insulin levels appears to be a satiety signal [42]. Insulin can cross the blood-brain barrier [43, 44] and insulin receptors can be found throughout the brain [45]. In animal studies, intracerebroventricular injection of insulin inhibits food intake [42, 46,47,48], and within the brain, the arcuate nucleus of the hypothalamus appears to be the major center regulating feeding behavior and energy balance [34••]. Proopiomelanocortin (POMC) and agouti-related peptide and neuropeptide Y (AgRP/NPY) neurons from the arcuate nucleus can control food intake, energy expenditure, and glucose metabolism [31, 34••]. POMC and AgRP/NPY neurons have insulin receptors [49] and are activated by insulin [50]. In men, but not in women, intranasal insulin decreases food intake [51]. Insulin levels in the periphery correlate with body weight [52] and may work as a peripheral signal informing the brain about the body’s energy status.
Leptin is secreted by the adipose tissue and leptin levels correlate with body weight [53]. Similar to insulin, intracerebroventricular injection of leptin reduces food intake [54], and activation of leptin receptors in POMC and AgRP/NPY neurons modulates food intake and energy expenditure [33, 34••]. In humans, leptin deficiency causes severe obesity [55], confirming a role of leptin as a satiety hormone in both animals and humans. GLP-1 are secreted by L-cells in the intestine in response to a meal [56]. The increase in GLP-1 levels stimulates the pancreas to secrete insulin and to decrease glucagon, and it is important for regulating glucose levels in the post-prandial period [57]. GLP-1 receptors are also found in the brain [58]. In humans, GLP-1 agonist can improve glucose control in patient with type 2 diabetes and promote weight loss [59]. Ghrelin, on the other hand, is an orexigenic hormone predominantly secreted by the stomach [60]. Ghrelin levels increase prior to a meal and decrease to its baseline levels after eating [61]. Intraperitoneal administration of ghrelin increases food intake and weight gain in mice [62] and increases hunger and food intake in humans [63].
In addition to hormones, nutrients also play a role in brain control of food intake [31, 64]. Among them, glucose [65, 66] and FFA [67] have been shown to affect brain control of food intake. Glucose levels have been described to regulate hunger/satiety and to promote food intake [31, 68, 69]. In the 1950s, Meyer proposed the glucostatic theory where the post-prandial rise in glucose levels could be sensed by the hypothalamus, which would relay the information to other brain regions to regulate food intake [70]. Indeed, a drop in glucose levels has been shown to precede a meal and may be important to initiate feeding [68, 69]. In addition, both centrally [71] or peripherally [72, 73] induced hypoglycemia can increase hunger and promote food intake.
In general, food intake and energy expenditure are very tightly regulated, with the brain playing a central role in integrating homeostatic, environmental, and hedonic signals to modulate food intake and energy expenditure. However, under the current epidemic of obesity, a shift towards increased energy storage has occurred, disrupting the fine-tuning of brain control of energy balance.
Hormonal Dysregulation in Obesity
Obesity results from an imbalance between food intake and energy expenditure. A sustained positive energy balance promotes energy storage in the form of fat and weight gain. However, it is not exactly known how the body adapts to this new higher body weight status. Indeed, body weight (even in obese individuals) tends to remain constant throughout the years [74]. Therefore, homeostatic control of food intake may adapt to this new increased energy storage level. Obese individuals, in comparison to normal-weight individuals, have elevated insulin [52] and leptin levels [53] and decreased ghrelin levels [75]. Insulin and leptin levels parallel to body weight status [52, 53] and insulin and leptin resistance have been proposed to play an important role in the pathogenesis of obesity [29, 34••]. Insulin and leptin resistance at the level of the arcuate nucleus decreases activation of POMC neurons and increases activation of AgRP/NPY neurons resulting in decreased satiety, increased hunger, and lower energy expenditure. Recent work has pointed out that fatty acid-induced inflammation and increased production of mitochondrial reactive oxygen species in POMC and AgRP/NPY neurons may play a role in the development of central insulin and leptin resistance in obesity (see review [34••]).
Hormonal Dysregulation in Weight Management
As described above, obese individuals demonstrate a distinct hormonal profile in comparison to lean individuals; which may influence energy balance and help maintain a higher weight. When this energy balance equilibrium is disrupted by diet-induced weight loss, hormonal and metabolic changes occur [76] in order to increase energy intake and decrease energy expenditure to eventually restore the previous energy equilibrium [77]. Weight loss from decreased calorie intake leads to decreased insulin, leptin, and ghrelin levels, and also decreased resting metabolic rate. However, neither weight loss nor long-term weight maintenance can be explained by these hormonal and metabolic changes [76, 78]. In addition, failures to weight management or regaining weight after weight loss are often observed in obese dieters, which suggests the presence of additional mechanisms underlying obesity other than hormonal regulation.
Food Cue and Feeding Behaviors
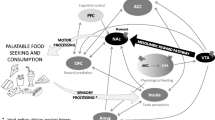
In addition to hormone and energy balance mechanisms, food cues in the environment play a crucial role in the development of obesity by altering physiological and brain responses. In modern society with abundant food cues, brain and physiological factors interact with environmental stimuli in modulating feeding behaviors. These environmental factors serve as triggers to arouse food craving and subsequent food intake. Therefore, the neurobiology of obesity and feeding behaviors under food cue exposure is complex. In the face of emotional challenges from abundant food cue environment, vulnerable individuals might have difficulties controlling high food craving and urge for food intake, increasing obesity risk.
Food Cue
Food cues include viewing or smelling of food stimuli, advertisements, or any cues or situations associated with food-related memories. These cues serve as conditioned stimuli that elicit food-related responses and subsequent food intake [79]. These cues accompany psychological (e.g., craving, urge) and physiological changes (salivation, hormone secretion) along with brain reactivity [80] that promote subsequent feeding behaviors [81]. These food cue-induced changes mobilize the appetitive system and increase motivation towards food consumption. As a result, highly palatable food cues often facilitate food-seeking even in a state of satiety and are regarded as an important risk factor for obesity. A meta-analysis study reporting the results from 45 studies showed the significant effect of food cue reactivity and craving on feeding behaviors [82•], suggesting that food cue substantially influences eating-related outcomes. To understand food-related behaviors and obesity risk, relevant neurobiological response to food cues should be clarified in the context of individual differences.
Food Cue and Obesity
Food or food-associated cues are risk factors for the development of obesity. A study showed that food cues, rather than hunger, are stronger stimulators of unhealthy eating behaviors in overweight individuals [83]. Obese individuals also show greater psychological and physiological responses to food cues. After the viewing and smelling of food cue (pizza), overweight individuals displayed greater salivation and enhanced desire for food compared to lean individuals [84]. Another study found that after repeated exposure to food cues involving gustatory presentation of food, obese women showed delayed decline of salivation response, suggesting their difficulties with the extinction of physiological response to food cues [85]. Furthermore, obese subjects showed increased attention to food images even in a satiated state [86]. Consistent with obese adults, obese adolescents displayed greater food cue responses compared to lean children. Overweight children are more susceptible to food-related triggers and have greater cue-related salivation flow, which was associated with increased food consumption [87]. In addition, children with higher BMI and waist scores demonstrated lower satiety and increased reactivity to food cues, even after controlling for age, sex, and parental education and BMI [88]. These studies point to a strong association between food cue reactivity and obesity risk.
In addition to peripheral responses, neuroimaging studies have provided compelling evidence on the effects of food cues on brain response. A review from 60 studies that were published in 2014 indicates that obese individuals consistently showed increased activation in regions involved in emotion and reward modulation including the insula (emotion/interoception) and orbitofrontal cortex (OFC; emotion/reward regulation) during the viewing of food stimuli [89]. Another study found that increased activity in the neural circuit of reward modulation was elicited in response to high-calorie versus low-calorie food pictures in obese individuals including the medial and lateral OFC, amygdala (emotion), ventral striatum (reward), and insula (interoception), and this difference was larger than lean individuals [90]. Furthermore, women having bulimia nervosa were found to have greater reactivity in reward regions in response to food images including the striatum and insula [91]. Across different studies, researchers have indicated that the food-conditioned responses stimulate the dopamine system in the ventral striatum, which plays a crucial role in the development of obesity, food-related addiction, and failure to weight management after treatment [92, 93].
Multimodal neuroimaging studies reported concurrent changes in both brain and hormone responses during the presentation of food cues. A combined fMRI study with concurrent hormone monitoring compared 25 obese with 15 lean adolescents during the viewing of food-related visual stimuli. The study found increased striatal-limbic response to high-calorie food pictures in obese adolescents, which was also associated with circulating leptin levels. This finding suggests food cue-related impairment in leptin signaling and brain functions in emotion and reward regions in adolescent obesity [94], emphasizing the significant impact of food cues on brain and endocrine functions. This finding is consistent with a study with 21 obese adults and 23 controls, reporting increased activity in the ventral striatum in response to visual food cue, which was associated with plasma leptin levels and BMI [95]. Taken together, neuroendocrine studies indicate altered brain and hormone responses to food cues, especially in obese individuals, suggesting that food cues increase appetitive response and motivation towards high-calorie foods and increase vulnerability to obesity.
Food Cue, Self-Control, and Weight Management
As food cues, including the smell, taste, and sight of highly palatable foods, arouse strong physiological responses and motivation towards eating behaviors, food cue-elicited responses can serve as a marker of identifying those who are not successful in weight management. It has been suggested that controlling food craving and intake is associated with an ability to control urges to eat in response to food cues [81]. Increased physiological response to food-related reward cues has shown to be a predictor of future weight gain. For example, obese individuals, who failed to lose weight during treatment, displayed increased activity in brain regions of reward modulation including the ventral striatum, insula, and anterior cingulate cortex in response to high-calorie food pictures. Increased activity in the ventral striatum during the presentation of food images was also associated with subsequent weight gain after 6 months follow-up [96]. Another study reported that increased response to food cues in emotion and reward regions (striatum and insula) predicted failure in weight management over a 9-month follow-up period [97].
Likewise, decreased responses to food cues (e.g., reduced salivary response) have predicted successful regulation of food intake and weight control in obese individuals [98]. In a Food Stroop task measuring cognitive control, individuals who were successful in weight loss maintenance showed slower reaction time in response to high-calorie foods, indicating less cognitive interference from food cues compared to normal-weight or obese individuals [99]. In addition, those who were successful in weight management showed decreased activity in reward brain regions and increased activity in regions of inhibitory control in response to food cues. Consistent with this, individuals who succeeded in weight loss showed greater activity in regions engaged in inhibitory control such as the left superior frontal lobe compared to normal-weight or obese individuals [100]. A literature review of brain imaging studies during visual food images indicated that obese individuals were more reactive to food pictures even during satiety, and brain activity in regulatory regions such as cingulate gyrus and precuneus predicted weight loss [89]. Studies also indicated that successful treatment intervention might be beneficial in assisting weight control by reducing physiological response to food cues. Individuals who received behavioral intervention were successful in weight loss and showed increased brain response to low-calorie food pictures at 6 months later [101]. In addition, decreased brain responses in mesolimbic reward regions to high calorie foods were found in 10 female patients after gastric bypass surgery [102]. Taken together, these studies indicate that food cues have strong impact on human physiology and feeding behaviors, and responses to food cues may serve as a marker of identifying those with increased vulnerability to obesity and be utilized as a predictor of treatment outcomes.
Homeostasis Overpowered by Food Cues
In the last few decades, significant scientific progress has been made in understanding the hormonal control of energy balance, and the importance of food cues as a contributing factor to weight gain. Food cues can overpower hormonal regulation of food intake in both healthy and obese subjects. In healthy non-obese individuals, an increase in plasma insulin levels via an intravenous insulin infusion (with the use of the hyperinsulinemic-euglycemic clamp technique) in comparison to a saline infusion did not affect brain responses to food pictures, and hunger levels increased in response to high-calorie food images [103•]. This suggests that brain activity and food-related behaviors during food cue exposure were not influenced by hyperinsulinemia or hormonal status. In addition, obese individuals, in comparison to normal weight, appear to be more sensitive to food cues both in the fasted and fed states [104, 105]. Brain responses to taste seem to influence weight gain [106••], and weight loss does not completely reverse brain responses to food cues [107]. These findings point to the significant impact of food cues on psychological and physiological responses. In particular, obesity-induced changes in homeostatic milieu will boost food cue-induced brain responses and elicit higher motivation towards environmental cues, which will promote food intake, help maintain weight at a higher threshold, and also regain weight after diet-induced weight loss.
Conclusions and Clinical Implications
Taken together, food cues elicit significant changes in brain and physiological functions, trigger feeding behaviors, and influence treatment outcomes. Overweight and obese individuals, in comparison with normal-weight individuals, have greater food cue-elicited responses in the brain and periphery, overpowering hormone and energy balance regulation. This dysregulated brain control or energy homeostasis facilitates continued food intake, resulting in unsuccessful weight management and poor treatment outcomes, which further contribute to obesity epidemic.
Therefore, in the setting of the current obesity epidemic, it is imperative to understand the pathogenesis of obesity in relation to food cues and to develop new ways to prevent and treat weight gain, and to avoid weight regain. In the last decades, anti-obesity drugs have been approved and are available in the market; however, their limited efficacy (~ 3–7% net weight loss) and side effects prevent their widespread use for the general population [59]. Bariatric surgeries, on the other hand, are more efficacious (21% weight loss 10 years after Roux-en-Y gastric bypass [108•]) but they are only indicated in more severe forms of obesity (BMI ≥ 40, or ≥ 35 and one or more severe obesity-related complications) [109].
In order to effectively contain the epidemic obesity, future research studies should consider the multiple factors contributing to weight gain, such as the environment (food cues) and the dysregulated homeostatic milieu, as a whole. Along with conventional treatments, behavioral interventions reducing food cue reactivity, craving and promoting life style for conscious awareness of food cue, and mindful eating might be helpful in reducing obesity. In addition, the current review supports policy-making initiatives for obesity prevention including regulating access to and advertisements of high caloric, unhealthy foods and promoting the public education of healthier diet.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
C D Fryar, M D Carroll and C L Ogden. Prevalence of overweight, obesity, and extreme obesity among adults: United States, trends 1960–1962 through 2009–2010. NCHS Health E-Stat. 2014.
Mitchell NS, Catenacci VA, Wyatt HR, Hill JO. Obesity: overview of an epidemic. Psychiatr Clin North Am. 2011;34(4):717–32.
Swinburn B, Sacks G, Ravussin E. Increased food energy supply is more than sufficient to explain the US epidemic of obesity. Am J Clin Nutr. 2009;90(6):1453–6.
Swinburn BA, Sacks G, Hall KD, McPherson K, Finegood DT, Moodie ML, et al. The global obesity pandemic: shaped by global drivers and local environments. Lancet. 2011;378(9793):804–14.
Yang W, Kelly T, He J. Genetic epidemiology of obesity. Epidemiol Rev. 2007;29:49–61.
Locke AE, Kahali B, Berndt SI, Justice AE, Pers TH, Day FR, et al. Genetic studies of body mass index yield new insights for obesity biology. Nature. 2015;518(7538):197–206.
Ogden CL, Yanovski SZ, Carroll MD, Flegal KM. The epidemiology of obesity. Gastroenterology. 2007;132(6):2087–102.
Hill JO, Peters JC. Environmental contributions to the obesity epidemic. Science. 1998;280(5368):1371–4.
King DM, Jacobson SH. What is driving obesity? A review on the connections between obesity and motorized transportation. Curr Obes Rep. 2017;6(1):3–9.
Gortmaker SL, Must A, Sobol AM, Peterson K, Colditz GA, Dietz WH. Television viewing as a cause of increasing obesity among children in the United States, 1986-1990. Arch Pediatr Adolesc Med. 1996;150(4):356–62.
Westerterp-Plantenga MS, Wijckmans-Duijsens NE, Verboeket-van de Venne WP, de Graaf K, van het Hof KH, Weststrate JA. Energy intake and body weight effects of six months reduced or full fat diets, as a function of dietary restraint. Int J Obes Relat Metab Disord. 1998;22(1):14–22.
Yao M, Roberts SB. Dietary energy density and weight regulation. Nutr Rev. 2001;59(8 Pt 1):247–58.
• Gittelsohn J, Trude A. Diabetes and obesity prevention: changing the food environment in low-income settings. Nutr Rev. 2017;75(suppl 1):62–9. A review of 10 community trials evaluating the impact of changes in the food environment in low-income minority settings.
Koplan JP, Dietz WH. Caloric imbalance and public health policy. JAMA. 1999;282(16):1579–81.
Cutler D, Glaeser E, Shapiro J. Why have Americans become more obese. J Econ Perspect. 2003;17(3):93–118.
• Louzada ML, Baraldi LG, Steele EM, Martins AP, Canella DS, Moubarac JC, et al. Consumption of ultra-processed foods and obesity in Brazilian adolescents and adults. Prev Med. 2015;81:9–15. A dietary survey from 30,243 individuals in Brazil demonstrated that ultra-processed food is associated with obesity and weight gain.
Putnam J, Allshouse J, Kantor LS. U.S. per capita food supply trends: more calories, refined carbohydrates, and fats. Food Rev: Mag Food Econ. 2002;25(3):2–15.
Ludwig DS. Technology, diet, and the burden of chronic disease. JAMA. 2011;305(13):1352–3.
Holt SH, Miller JC, Petocz P, Farmakalidis E. A satiety index of common foods. Eur J Clin Nutr. 1995;49(9):675–90.
• Fardet A. Minimally processed foods are more satiating and less hyperglycemic than ultra-processed foods: a preliminary study with 98 ready-to-eat foods. Food Funct. 2016;7(5):2338–46. Evaluation of the glycemic glucose equivalent (GGE) of ultraprocessed food on satiety and blood glucose levels. Ultraprocessed food was associated with higher glycemic response and decreased satiety.
Volkow ND, Wise RA. How can drug addiction help us understand obesity? Nat Neurosci. 2005;8:555–60.
Cohen DA. Obesity and the built environment: changes in environmental cues cause energy imbalances. Int J Obes. 2008;32(Suppl 7):S137–42.
Volkow ND, Wang GJ, Fowler JS, Tomasi D, Baler R. Food and drug reward: overlapping circuits in human obesity and addiction. In: Carter CS, Dalley JW, editors. Brain imaging in behavioral neuroscience. Berlin, Heidelberg: Springer Berlin Heidelberg; 2012. p. 1–24.
Carnell S, Gibson C, Benson L, Ochner CN, Geliebter A. Neuroimaging and obesity: current knowledge and future directions. Obes Rev. 2012;13(1):43–56.
Berthoud HR. Homeostatic and non-homeostatic pathways involved in the control of food intake and energy balance. Obesity (Silver Spring). 2006;14(Suppl 5):197S–200S.
Christensen CM, Navazesh M. Anticipatory salivary flow to the sight of different foods. Appetite. 1984;5(4):307–15.
Pangborn RM, Berggren B. Human parotid secretion in response to pleasant and unpleasant odorants. Psychophysiology. 1973;10(3):231–7.
Lindemann B. Receptors and transduction in taste. Nature. 2001;413(6852):219–25.
Morton GJ, Meek TH, Schwartz MW. Neurobiology of food intake in health and disease. Nat Rev Neurosci. 2014;15(6):367–78.
Kairupan TS, Amitani H, Cheng KC, Runtuwene J, Asakawa A, Inui A. Role of gastrointestinal hormones in feeding behavior and obesity treatment. J Gastroenterol. 2016;51(2):93–103.
Gao Q, Horvath TL. Neurobiology of feeding and energy expenditure. Annu Rev Neurosci. 2007;30:367–98.
Strader AD, Woods SC. Gastrointestinal hormones and food intake. Gastroenterology. 2005;128(1):175–91.
Begg DP, Woods SC. The endocrinology of food intake. Nat Rev Endocrinol. 2013;9(10):584–97.
•• Timper K, Bruning JC. Hypothalamic circuits regulating appetite and energy homeostasis: pathways to obesity. Dis Model Mech. 2017;10(6):679–89. A review with the latest information about the role of the hypothalamus on feeding and the pathogenesis of obesity.
Berthoud HR, Bereiter DA, Trimble ER, Siegel EG, Jeanrenaud B. Cephalic phase, reflex insulin secretion neuroanatomical and physiological characterization. Diabetologia. 1981;20(Suppl 1):393–401.
Drazen DL, Vahl TP, D’Alessio DA, Seeley RJ, Woods SC. Effects of a fixed meal pattern on ghrelin secretion: evidence for a learned response independent of nutrient status. Endocrinology. 2006;147(1):23–30.
Simon C, Schlienger JL, Sapin R, Imler M. Cephalic phase insulin secretion in relation to food presentation in normal and overweight subjects. Physiol Behav. 1986;36(3):465–9.
Schussler P, Kluge M, Yassouridis A, Dresler M, Uhr M, Steiger A. Ghrelin levels increase after pictures showing food. Obesity (Silver Spring). 2012;20(6):1212–7.
Vahl TP, Drazen DL, Seeley RJ, D’Alessio DA, Woods SC. Meal-anticipatory glucagon-like peptide-1 secretion in rats. Endocrinology. 2010;151(2):569–75.
Chen M, Porte D Jr. The effect of rate and dose of glucose infusion on the acute insulin response in man. J Clin Endocrinol Metab. 1976;42(6):1168–75.
Polonsky KS, Given BD, Van Cauter E. Twenty-four-hour profiles and pulsatile patterns of insulin secretion in normal and obese subjects. J Clin Invest. 1988;81(2):442–8.
Woods SC, Lotter EC, McKay LD, Porte D. Chronic intracerebroventricular infusion of insulin reduces food intake and body weight of baboons. Nature. 1979;282:503–5.
Baura GD, Foster DM, Porte D Jr, Kahn SE, Bergman RN, Cobelli C, et al. Saturable transport of insulin from plasma into the central nervous system of dogs in vivo. A mechanism for regulated insulin delivery to the brain. J Clin Invest. 1993;92(4):1824–30.
Banks WA, Jaspan JB, Kastin AJ. Selective, physiological transport of insulin across the blood-brain barrier: novel demonstration by species-specific radioimmunoassays. Peptides. 1997;18(8):1257–62.
Unger J, McNeill TH, Moxley RT 3rd, White M, Moss A, Livingston JN. Distribution of insulin receptor-like immunoreactivity in the rat forebrain. Neuroscience. 1989;31(1):143–57.
Florant GL, Singer L, Scheurink AJ, Park CR, Richardson RD, Woods SC. Intraventricular insulin reduces food intake and body weight of marmots during the summer feeding period. Physiol Behav. 1991;49(2):335–8.
Honda K, Kamisoyama H, Saneyasu T, Sugahara K, Hasegawa S. Central administration of insulin suppresses food intake in chicks. Neurosci Lett. 2007;423(2):153–7.
Brown LM, Clegg DJ, Benoit SC, Woods SC. Intraventricular insulin and leptin reduce food intake and body weight in C57BL/6J mice. Physiol Behav. 2006;89(5):687–91.
Konner AC, Janoschek R, Plum L, Jordan SD, Rother E, Ma X, et al. Insulin action in AgRP-expressing neurons is required for suppression of hepatic glucose production. Cell Metab. 2007;5(6):438–49.
Williams KW, Margatho LO, Lee CE, Choi M, Lee S, Scott MM, et al. Segregation of acute leptin and insulin effects in distinct populations of arcuate proopiomelanocortin neurons. J Neurosci. 2010;30(7):2472–9.
Hallschmid M, Benedict C, Schultes B, Fehm HL, Born J, Kern W. Intranasal insulin reduces body fat in men but not in women. Diabetes. 2004;53(11):3024–9.
Bagdade JD, Bierman EL, Porte D Jr. The significance of basal insulin levels in the evaluation of the insulin response to glucose in diabetic and nondiabetic subjects. J Clin Invest. 1967;46(10):1549–57.
Considine RV, Sinha MK, Heiman ML, Kriauciunas A, Stephens TW, Nyce MR, et al. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med. 1996;334(5):292–5.
Campfield LA, Smith FJ, Guisez Y, Devos R, Burn P. Recombinant mouse OB protein: evidence for a peripheral signal linking adiposity and central neural networks. Science. 1995;269(5223):546–9.
Montague CT, Farooqi IS, Whitehead JP, Soos MA, Rau H, Wareham NJ, et al. Congenital leptin deficiency is associated with severe early-onset obesity in humans. Nature. 1997;387(6636):903–8.
Brubaker PL, Anini Y. Direct and indirect mechanisms regulating secretion of glucagon-like peptide-1 and glucagon-like peptide-2. Can J Physiol Pharmacol. 2003;81(11):1005–12.
Willms B, Werner J, Holst JJ, Orskov C, Creutzfeldt W, Nauck MA. Gastric emptying, glucose responses, and insulin secretion after a liquid test meal: effects of exogenous glucagon-like peptide-1 (GLP-1)-(7-36) amide in type 2 (noninsulin-dependent) diabetic patients. J Clin Endocrinol Metab. 1996;81(1):327–32.
Campos RV, Lee YC, Drucker DJ. Divergent tissue-specific and developmental expression of receptors for glucagon and glucagon-like peptide-1 in the mouse. Endocrinology. 1994;134(5):2156–64.
Srivastava G, Apovian CM. Current pharmacotherapy for obesity. Nat Rev Endocrinol. 2018;14(1):12–24.
Date Y, Kojima M, Hosoda H, Sawaguchi A, Mondal MS, Suganuma T, et al. Ghrelin, a novel growth hormone-releasing acylated peptide, is synthesized in a distinct endocrine cell type in the gastrointestinal tracts of rats and humans. Endocrinology. 2000;141(11):4255–61.
Higgins SC, Gueorguiev M, Korbonits M. Ghrelin, the peripheral hunger hormone. Ann Med. 2007;39(2):116–36.
Asakawa A, Inui A, Kaga T, Yuzuriha H, Nagata T, Ueno N, et al. Ghrelin is an appetite-stimulatory signal from stomach with structural resemblance to motilin. Gastroenterology. 2001;120(2):337–45.
Wren AM, Seal LJ, Cohen MA, Brynes AE, Frost GS, Murphy KG, et al. Ghrelin enhances appetite and increases food intake in humans. J Clin Endocrinol Metab. 2001;86(12):5992.
Myers MG Jr, Olson DP. Central nervous system control of metabolism. Nature. 2012;491(7424):357–63.
Davis JD, Wirtshafter D, Asin KE, Brief D. Sustained intracerebroventricular infusion of brain fuels reduces body weight and food intake in rats. Science. 1981;212(4490):81–3.
Page KA, Chan O, Arora J, Belfort-Deaguiar R, Dzuira J, Roehmholdt B, et al. Effects of fructose vs glucose on regional cerebral blood flow in brain regions involved with appetite and reward pathways. JAMA. 2013;309(1):63–70.
Obici S, Feng Z, Morgan K, Stein D, Karkanias G, Rossetti L. Central administration of oleic acid inhibits glucose production and food intake. Diabetes. 2002;51(2):271–5.
Louis-Sylvestre J, Le Magnen J. Fall in blood glucose level precedes meal onset in free-feeding rats. Neurosci Biobehav Rev. 1980;4(Suppl 1):13–5.
Campfield LA, Smith FJ, Rosenbaum M, Hirsch J. Human eating: evidence for a physiological basis using a modified paradigm. Neurosci Biobehav Rev. 1996;20:133–7.
Mayer J. Glucostatic mechanism of regulation of food intake. N Engl J Med. 1953;249(1):13–6.
Slusser PG, Ritter RC. Increased feeding and hyperglycemia elicited by intracerebroventricular 5-thioglucose. Brain Res. 1980;202(2):474–8.
Lotter EC, Woods SC. Injections of insulin and changes of body weight. Physiol Behav. 1977;18(2):293–7.
Thompson DA, Campbell RG. Hunger in humans induced by 2-deoxy-D-glucose: glucoprivic control of taste preference and food intake. Science. 1977;198(4321):1065–8.
Flatt JP. What do we most need to learn about food intake regulation? Obes Res. 1998;6(4):307–10.
Tschop M, Weyer C, Tataranni PA, Devanarayan V, Ravussin E, Heiman ML. Circulating ghrelin levels are decreased in human obesity. Diabetes. 2001;50(4):707–9.
Fothergill E, Guo J, Howard L, Kerns JC, Knuth ND, Brychta R, et al. Persistent metabolic adaptation 6 years after “The Biggest Loser” competition. Obesity (Silver Spring). 2016;24(8):1612–9.
Sumithran P, Prendergast LA, Delbridge E, Purcell K, Shulkes A, Kriketos A, et al. Long-term persistence of hormonal adaptations to weight loss. N Engl J Med. 2011;365(17):1597–604.
Strohacker K, McCaffery JM, MacLean PS, Wing RR. Adaptations of leptin, ghrelin or insulin during weight loss as predictors of weight regain: a review of current literature. Int J Obes. 2014;38(3):388–96.
Martin-Soelch C, Linthicum J, Ernst M. Appetitive conditioning: neural bases and implications for psychopathology. Neurosci Biobehav Rev. 2007;31(3):426–40.
Yokum S, Ng J, Stice E. Attentional bias to food images associated with elevated weight and future weight gain: an fMRI study. Obesity. 2011;19(9):1775–83.
Jansen A. A learning model of binge eating: cue reactivity and cue exposure. Behav Res Ther. 1998;36(3):257–72.
• Boswell RG, Kober H. Food cue reactivity and craving predict eating and weight gain: a meta-analytic review. Obes Rev. 2016;17(2):159–77. Meta-analyses of 45 studies evaluating the effect of food cues on eating behavior identified that visual food cues were as effective as real food (and more that olfactory cues) in affecting eating behavior and weight gain.
Cleobury L, Tapper K. Reasons for eating ‘unhealthy’ snacks in overweight and obese males and females. J Hum Nutr Diet. 2014;27(4):333–41.
Ferriday D, Brunstrom J. ‘I just can’t help myself’: effects of food-cue exposure in overweight and lean individuals. Int J Obes. 2011;35(1):142–9.
Epstein LH, Paluch R, Coleman KJ. Differences in salivation to repeated food cues in obese and nonobese women. Psychosom Med. 1996;58(2):160–4.
Castellanos EH, Charboneau E, Dietrich MS, Park S, Bradley BP, Mogg K, et al. Obese adults have visual attention bias for food cue images: evidence for altered reward system function. Int J Obes. 2009;33(9):1063–73.
Jansen A, Theunissen N, Slechten K, Nederkoorn C, Boon B, Mulkens S, et al. Overweight children overeat after exposure to food cues. Eat Behav. 2003;4(2):197–209.
Carnell S, Wardle J. Appetite and adiposity in children: evidence for a behavioral susceptibility theory of obesity. Am J Clin Nutr. 2008;88(1):22–9.
Pursey KM, Stanwell P, Callister RJ, Brain K, Collins CE, Burrows TL. Neural responses to visual food cues according to weight status: a systematic review of functional magnetic resonance imaging studies. Front Nutr. 2014;1:7.
Stoeckel LE, Weller RE, Cook EW, Twieg DB, Knowlton RC, Cox JE. Widespread reward-system activation in obese women in response to pictures of high-calorie foods. NeuroImage. 2008;41:636–47.
Brooks SJ, Owen G, Uher R, Friederich H-C, Giampietro V, Brammer M, et al. Differential neural responses to food images in women with bulimia versus anorexia nervosa. PLoS One. 2011;6(7):e22259.
Berthoud H-R, Lenard NR, Shin AC. Food reward, hyperphagia, and obesity. Am J Phys Regul Integr Comp Phys. 2011;300(6):R1266–R77.
Volkow ND, Wang G-J, Fowler JS, Tomasi D. Addiction circuitry in the human brain. Annu Rev Pharmacol Toxicol. 2012;52:321–36.
Jastreboff AM, Lacadie C, Seo D, Kubat J, Van Name MA, Giannini C, et al. Leptin is associated with exaggerated brain reward and emotion responses to food images in adolescent obesity. Diabetes Care. 2014;37(11):3061–8.
Grosshans M, Vollmert C, Vollstadt-Klein S, Tost H, Leber S, Bach P, et al. Association of leptin with food cue-induced activation in human reward pathways. Arch Gen Psychiatry. 2012;69(5):529–37.
Demos KE, Heatherton TF, Kelley WM. Individual differences in nucleus accumbens activity to food and sexual images predict weight gain and sexual behavior. J Neurosci. 2012;32(16):5549–52.
Murdaugh DL, Cox JE, Cook EW, Weller RE. fMRI reactivity to high-calorie food pictures predicts short-and long-term outcome in a weight-loss program. NeuroImage. 2012;59(3):2709–21.
Jansen A, Stegerman S, Roefs A, Nederkoorn C, Havermans R. Decreased salivation to food cues in formerly obese successful dieters. Psychother Psychosom. 2010;79(4):257–8.
Phelan S, Hassenstab J, McCaffery JM, Sweet L, Raynor HA, Cohen RA, et al. Cognitive interference from food cues in weight loss maintainers, normal weight, and obese individuals. Obesity. 2011;19(1):69–73.
McCaffery JM, Haley AP, Sweet LH, Phelan S, Raynor HA, Del Parigi A, et al. Differential functional magnetic resonance imaging response to food pictures in successful weight-loss maintainers relative to normal-weight and obese controls. Am J Clin Nutr. 2009;90(4):928–34.
Deckersbach T, Das SK, Urban LE, Salinardi T, Batra P, Rodman AM, et al. Pilot randomized trial demonstrating reversal of obesity-related abnormalities in reward system responsivity to food cues with a behavioral intervention. Nutr Diab. 2014;4:e129.
Ochner CN, Kwok Y, Conceição E, Pantazatos SP, Puma LM, Carnell S, et al. Selective reduction in neural responses to high calorie foods following gastric bypass surgery. Ann Surg. 2011;253:502–7.
• Belfort-De Aguiar R, Seo D, Naik S, Hwang J, Lacadie C, Schmidt C, et al. Food image-induced brain activation is not diminished by insulin infusion. Int J Obes (Lond). 2016;40(11):1679–86. In this study, brain responses to food cues were not affected by an intravenous insulin infusion, in comparison to a saline infusion.
Dimitropoulos A, Tkach J, Ho A, Kennedy J. Greater corticolimbic activation to high-calorie food cues after eating in obese vs. normal-weight adults. Appetite. 2012;58(1):303–12.
Le DS, Pannacciulli N, Chen K, Del Parigi A, Salbe AD, Reiman EM, et al. Less activation of the left dorsolateral prefrontal cortex in response to a meal: a feature of obesity. Am J Clin Nutr. 2006;84:725–31.
•• Sun X, Kroemer NB, Veldhuizen MG, Babbs AE, de Araujo IE, Gitelman DR, et al. Basolateral amygdala response to food cues in the absence of hunger is associated with weight gain susceptibility. J Neurosci. 2015;35(20):7964–76. Activity in the amygdala in response to tasting a milkshake in the sated, but not in the hungry state, predicted weight gain in susceptible individuals.
McCaffery JM, Haley AP, Sweet LH, Phelan S, Raynor HA, Del Parigi A, et al. Differential functional magnetic resonance imaging response to food pictures in successful weight-loss maintainers relative to normal-weight and obese controls. Am J Clin Nutr. 2009;90:928–34.
• Maciejewski ML, Arterburn DE, Van Scoyoc L, Smith VA, Yancy WS Jr, Weidenbacher HJ, et al. Bariatric surgery and long-term durability of weight loss. JAMA Surg. 2016;151(11):1046–55. Study showing the 10-year sustained effect of bariatric surgery on weight loss.
Garvey WT, Mechanick JI, Brett EM, Garber AJ, Hurley DL, Jastreboff AM, et al. American Association of Clinical Endocrinologists and American College of Endocrinology comprehensive clinical practice guidelines for medical care of patients with obesity. Endocr Pract. 2016;22(Suppl 3):1–203.
Funding
This work was funded in part by grants from the NIH (K23 DK098286-02 to R Belfort-DeAguiar from the NIDDK), (K08-AA023545 to D Seo from the NIAAA), and a Young Investigator Grant (Seo) from the Brain and Behavior Research Foundation. This publication was made possible by CTSA Grant Number UL1 TR00142 from the National Center for Advancing Translational Science (NCATS), a component of the National Institutes of Health (NIH). Its contents are solely the responsibility of the authors and do not necessarily represent the official view of NIH.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
Renata Belfort-DeAguiar and Dongju Seo declare they have no conflict of interest.
Human and Animal Rights and Informed Consent
All reported studies/experiments with human subjects performed by the authors have been previously published and complied with all applicable ethical standards (including the Helsinki declaration and its amendments, institutional/national research committee standards, and international/national/institutional guidelines).
Additional information
This article is part of the Topical Collection on Obesity Treatment
Rights and permissions
About this article

Cite this article
Belfort-DeAguiar, R., Seo, D. Food Cues and Obesity: Overpowering Hormones and Energy Balance Regulation. Curr Obes Rep 7, 122–129 (2018). https://doi.org/10.1007/s13679-018-0303-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13679-018-0303-1