
Abstract
Commercial perennial agriculture is prone to declining productivity due to negative plant-soil feedback. An alternative to costly and environmentally harmful conventional treatment such as soil fumigation could be to manipulate soil microbial diversity through careful selection and management of cover crop mixtures. Although cover crops are already used in these systems for other reasons, their capacity to influence soil biota is unexploited. Here, we examine the role of plant diversity and identity on plant-soil feedbacks in the context of perennial agriculture. We identify key microorganisms involved in these feedbacks and explore plant-based strategies for mitigating decline of perennial crop plants. We conclude that (1) increasing plant diversity increases soil microbial diversity, minimizing the proliferation of soil-borne pathogens; (2) populations of beneficial microbes can be increased by increasing plant functional group richness, e.g., legumes, C4 grasses, C3 grasses, and non-leguminous forbs; (3) brassicas suppress fungal pathogens and promote disease-suppressive bacteria; (4) native plants may further promote beneficial soil microbiota; and (5) frequent tillage, herbicide use, and copper fungicides can harm populations of beneficial microbes and, in some cases, contribute to greater crop decline. Non-crop vegetation management is a viable and cost-effective means of minimizing crop decline in perennial monocultures but is in need of more direct experimental investigation in perennial agroecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Contents
-
1. Introduction
-
2. Manipulating cover crops to influence soil microbial communities
-
2.1 Root exudates attract rhizosphere microbes
-
2.2 Plant litter affects decomposer community
-
2.3 Plant cover affects soil moisture
-
-
3. The role of beneficial microbes in crop decline
-
3.1 AM Fungi
-
3.2 Disease-protective fungi
-
3.3 Disease-suppressive bacteria
-
3.4 Entomopathogenic fungi
-
-
4. Considerations for managing non-crop vegetation
-
4.1 Plant diversity
-
4.2 Considerations within plant functional groups
-
4.2.1 Legumes
-
4.2.2 C3 and C4 grasses
-
4.2.3 Brassicas
-
-
4.3 Native plant species in cover crops
-
4.4 Management practices that disrupt cover crop-mediated soil feedbacks
-
4.4.1 Tillage
-
4.4.2 Herbicide use
-
4.4.3 Fungicides
-
-
-
5. Conclusions
-
Acknowledgments
-
References
1 Introduction
Perennial crops often experience reduced productivity over time due to the accumulation of soil-borne pests and pathogens (Hamel et al. 2005; Mazzola and Manici 2012; Úrbez-Torres et al. 2014). This is particularly problematic in woody perennial systems where crop rotation is not possible, and ultimately, replanting is necessary to restore production levels. However, addressing this problem from an ecological perspective may lead to more sustainable solutions or avoidance of decline altogether.
An ecological concept that is useful for understanding crop decline is plant-soil feedbacks. This concept describes the reciprocal effects of plants and their associated soil microbial communities (Bever 1994). Negative soil feedback occurs when plants promote soil microorganisms that are deleterious to their own growth, contributing to the maintenance of plant coexistence in natural systems through density-dependent regulation of dominant species (Bever et al. 2015). In perennial monocultures, however, negative feedback leads to crop decline and replant problems (Hamel et al. 2005; Mazzola and Manici 2012). That is, the deleterious soil microbial community also suppresses the growth of neighboring crop plants.
The negative effect of soil microbial communities in monocultures is not altogether surprising, given the negative relationship between biodiversity and the frequency of parasitism (Civitello et al. 2015). In many systems, low levels of diversity will allow a parasite of the dominant host species to more easily find a suitable host. Increased diversity makes hosts more difficult to find and disease outbreaks less frequent and leads to the “dilution effect” associated with high species richness (Keesing et al. 2010). For plant-soil ecosystems in particular, it is well established that increased soil diversity decreases incidence of plant disease (Garbeva et al. 2004a; van Elsas et al. 2002) and improves plant productivity (van der Heijden et al. 1998, 2008).
Can growers capitalize on this dilution effect by increasing soil microbial diversity in perennial systems? While many mechanisms contribute to forming soil microbial communities, e.g., abiotic filters (Fierer and Jackson 2006; Lauber et al. 2008), there is an extensive body of literature documenting the ability of plants to “train” their associated microbial communities (Badri and Vivanco 2009; Fanin et al. 2014; Hartmann et al. 2009; Rovira 1969). While growers are limited in their ability to manipulate the diversity of crop plants in their cropping system, cover crop identity and diversity can be an efficient way to increase soil microbial diversity and suppress soil-borne pests that cause crop decline (Garbeva et al. 2004a). Cover crops are already a common feature in many perennial systems (Fig. 1), but their potential impact on the biotic component of soils is often overlooked. For the purpose of this review, we define the term “cover crop” as managed vegetation grown between crop plant rows, including annual and perennial swards.

Examples of common cover cropping strategies and experimentation at the Summerland Research and Development Center, British Columbia. Top: a permanent cover of mixed grass species in a cherry orchard and bottom: a long-term cover crop experiment in wine grapes evaluating mixtures and monocultures of native and introduced grasses
Here, we incorporate ecological knowledge of plant-soil feedbacks into the context of perennial agriculture to explore the use of cover crops to increase microbial diversity and manage crop decline. The specific aims of this review are to (1) synthesize our current understanding of how plant communities influence soil microbial communities into the context of cover crops; (2) highlight key beneficial soil microbes and their role in affecting crop decline; and (3) present plant-based strategies to mitigate decline of perennial crops through soil microbial diversity.
2 Manipulating cover crops to influence soil microbial communities
2.1 Root exudates attract rhizosphere microbes
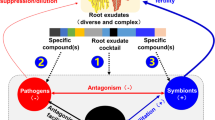
Perhaps, the most well-studied mechanism through which plants affect the soil microbial community is root exudates (Badri and Vivanco 2009; Broeckling et al. 2008; Rovira 1969). Root exudates contain C-rich compounds, including amino acids, organic acids, sugars, phenolics, secondary metabolites, and proteins that are excreted mostly from root hairs and the cells immediately behind the penetrating root tip (Badri and Vivanco 2009). Root exudates attract and sustain a variety of rhizosphere microorganisms, including arbuscular mycorrhizal (AM) fungi (Akiyama et al. 2005), entomopathogens in response to root herbivory (Rasmann et al. 2005), and N-fixing bacteria (Long 2001). However, root exudates also attract host-specific pathogens (Nicol et al. 2003; Hamel et al. 2005; Hofmann et al. 2009) and thus can lead to both positive and negative soil feedback. Since exudate composition, quantity, and seasonality depend on host plant identity (Broeckling et al. 2008; Schweitzer et al. 2008), a cover crop that includes a variety of plants should be able to maintain greater diversity of root-associated microbes with higher overall benefits to crops (Bardgett and van der Putten 2014; Garbeva et al. 2004a) (Fig. 2).

Depiction of how plant-soil feedbacks might influence crop decline in the context of a highly diverse permanent vineyard cover crop (adapted from Bever et al. 2010). a A diverse cover crop influences the soil microbial community through deposition of root exudates and litter creating a diverse microbial community (SMCcover) which provides feedback b to the cover crop maintaining plant diversity. c The vine “trains” its own soil microbial community (SMCcrop), which may include pest and pathogen buildup due to a low diversity of plant-derived resources. d If plant diversity in the vineyard is limited to vines only, SMCcrop would feedback negatively on the vine leading to crop decline due to monoculture. e However, a highly diverse SMCcover provides inoculum of microbial diversity that may interact with SMCcrop and the vine directly, increasing the microbial diversity available to roots of the vine and preventing decline due to negative feedback
2.2 Plant litter affects decomposer community
While growers are familiar with the effect of soil organic material on disease suppression, e.g., compost addition (Hoitink and Boehm 1999), a similar effect can occur with cover crop plant litter. The identity of decomposing plant litter has been shown to affect both the activity (Bardgett and Shine 1999) and community structure of soil microbes (Fanin et al. 2014). Litter quality is largely a result of both the ratio of carbon to nutrients and the proportions of different C chemistry, i.e., low-molecular-weight compounds (easily labile) versus more complex forms such as lignin (more recalcitrant) (Cotrufo et al. 2013).
The quality and quantity of litter differ greatly among plant species (Cornelissen and Thompson 1997) and can determine saprotrophic community composition (Fanin et al. 2014). High-quality plant litter (lower C/nutrient ratio, greater proportion of labile to recalcitrant C chemistry) is broken down more rapidly and thus favors faster-growing copiotrophic microbes, including disease-suppressive pseudomonads (Bastian et al. 2009), while oligotrophic microbes such as fungal decomposers (Holland and Coleman 1987) and Acidobacteria (Bastian et al. 2009) are largely responsible for the decomposition of low-quality litter. Recently, Fanin et al. (2016) compared microbial communities from soils to which litter was added that differed in C/N ratio, lignin, and total nutrients and found that litter type had a strong effect on shaping soil microbial communities, surpassed only by the influence of soil type. Other aspects of plant litter chemistry, such as glucosinolate content, can also have a dramatic and long-term effect on the soil microbial community (Mazzola et al. 2015). Stimulating soil saprotrophic communities with a diverse composition of plant litter may reduce soil-borne pathogens, either through increased competition for resources (Fontaine et al. 2003) or by increasing the abundance of beneficial, pathogen antagonistic microbes (Garbeva et al. 2004b).
Whether litter is left on the surface (as in a perennial cover crop) or incorporated into the soil via tillage will also affect the decomposer communities and, thus, many pathogens (Govaerts et al. 2007; Holland and Coleman 1987). When litter is left on the soil surface, as in a no-till system, saprotrophic microbial communities become more structured due to the stratification of different nutrients through the soil (Holland and Coleman 1987). This heterogeneity of resources leads to increased microbial diversity found in no-till systems (Govaerts et al. 2007) and could increase the ability of soils to suppress disease (Stirling et al. 2012), but this has not been investigated specifically in a cover cropped perennial system.
2.3 Plant cover affects soil moisture
Another way in which cover crops may influence the soil microbial community is by altering soil moisture dynamics (Bezemer et al. 2006; Lange et al. 2014). In addition to differences in transpiration, plants differ in the proportion of the soil surface covered by their canopy, affecting temperature and evaporation from the soil and, therefore, soil moisture (Lange et al. 2014). In dryland ecosystems, more water from the top 15 cm of soil is typically lost to evaporation than is used by plants (Loik et al. 2004). The maintenance of soil cover, as in a no-till system, can decrease soil temperature and increase soil moisture retention at shallow depths compared to tilled soil (Pannkuk et al. 1997). Depending on the traits of the ground cover vegetation, e.g., root system architecture, photosynthetic pathway, and dormant states, differences in soil moisture from plant cover can be ecologically significant for soil microbes (Chowdhury et al. 2011; de Vries et al. 2012). Of course, this can be exacerbated by soil texture and salt content (Chowdhury et al. 2011).
Tolerance of microbes to fluctuating soil moisture levels is largely phylogenetically determined (Placella et al. 2012). Microorganisms that provide ecosystem services at shallower depths, such as entomopathogenic fungi, may benefit most from the preservation of moisture by plant cover (Pell et al. 2010). This mechanism has not been tested specifically, but there is some evidence for the persistence of natural populations of Beauveria bassiana when a cover is maintained (Shapiro-Ilan et al. 2012). Although plant cover and, thus, soil moisture can be an important driver of a soil microbial community (Lange et al. 2014), more work needs to be done to elucidate this effect on crop decline outcomes as some pathogens also benefit from increased soil moisture (Pieczarka and Abawi 1978; Kuan and Erwin 1982).
3 The role of beneficial microbes in crop decline
While discouraging negative feedback is a critical part of managing crop decline, promoting beneficial microbes may be as important. Because generalist pathogens may not be as affected by the diversity of a plant community per se and are common in some perennial systems, e.g., “Cylindrocarpon” in apple (Mazzola and Manici 2012), avocado (Vitale et al. 2012), and grape (Úrbez-Torres et al. 2014), the ability of a cover crop to encourage beneficial microbes might be paramount to avoiding crop decline if these pathogens are prevalent. Microbial antagonists such as AM fungi, disease-protective fungi, e.g., Trichoderma spp., and disease-suppressive bacteria, e.g., pseudomonads, are instrumental in preventing soil-borne disease whether through competition for resources or niche space (Larsen and Bodker 2001), direct antibiosis (Haas and Defago 2005), or by induction of plant systemic resistance (Pozo et al. 2002). Entomopathogenic fungi provide protection from soil-dwelling insect pests, outbreaks of which are another form of negative feedback in these systems. Here, we examine the role that beneficial microbes play in preventing crop decline and provide evidence that their populations may be regulated, in part, by plant communities.
3.1 Mycorrhizal fungi
Most perennial crops are mycorrhizal, forming a root symbiosis with fungi, e.g., apple (Gnekow and Marschner 1989), citrus (Menge et al. 1978), olive (Roldán-Fajardo and Barea 1985), raspberry (Taylor and Harrier 2000), and grape (Trouvelot et al. 2015). This mutualism confers many benefits to hosts, particularly nutritional and stress tolerance (Smith and Read 2008), but is threatened in most agricultural systems (Verbruggen et al. 2010) due to management activities that inhibit the fungi such as tillage (Brito et al. 2012), fungicide use (Graham et al. 1986), and some forms of weed control (Schreiner et al. 2001).
AM fungi (phylum Glomeromycota), which are the most widespread fungal mutualists, are considered pathogen antagonists in addition to nutritional symbionts (Azcón-Aguilar and Barea 1996; Cameron et al. 2013). Competition for root space is one way that AM fungi can limit root disease (Cameron et al. 2013). AM fungi may also inhibit pathogenic fungi through induced systemic resistance (ISR), by evoking an enhanced immune response in the host plant (Pozo et al. 2002). Finally, AM fungal hyphae in the soil cultivate their own diverse microbial communities (Scheublin et al. 2010) and microbes that inhabit this “hyphosphere” can include fungi and bacteria that play a role in protection from soil-borne pathogens (Filion et al. 1999).
Because some AM fungal taxa may offer more disease protection than others (Maherali and Klironomos 2007; Sikes et al. 2009), greater diversity of AM fungi could lead to broader disease suppression due to the increased probability of the presence of taxa effective in guarding against pathogens. Due to the tight link between plant diversity and AM fungal diversity (Hart et al. 2003; van der Heijden et al. 1998), using diverse cover crop mixes could increase the suite of AM fungal partners available to crop plants, with a greater chance of symbioses forming between crop plants and multiple AM fungi that are effective in alleviating both abiotic and biotic stresses. Although nutrient transfer from a cover crop (Bromus hordeaceus or Medicago polymorpha) to neighboring grapevines has been shown to be mediated by AM fungi (Cheng and Baumgartner 2004, 2006), the effect of vegetation type and diversity on the AM fungal community and its relationship to disease outcomes has not been studied.
3.2 Disease-protective fungi
Many saprotrophic fungi opportunistically colonize roots as endophytes and aid in controlling pathogenic fungi. A notable example is Trichoderma, which can parasitize other fungi using cell wall-degrading enzymes and antibiotic compounds (Harman et al. 2004). Non-pathogenic strains of Fusarium oxysporum have been observed to function in a similar way (Benhamou et al. 2002), and there are myriad other fungi that are also known to protect roots from pathogens, e.g., Penicillium (Hossain et al. 2007) and Clonostachys (Luongo et al. 2005). Because many of these fungi can also promote increased root and plant growth (Harman et al. 2004), they may also contribute to positive feedbacks similar to those experienced by plants associating with AM fungal mutualists (Avis et al. 2008).
Some disease-protective fungi are sold commercially as biocontrol agents, e.g., Trichoderma spp., but they are naturally common in soil and may be affected by plant diversity as well as soil disturbance due to their additional roles as decomposers and endophytes. There is some empirical evidence for a plant host effect on populations of pathogen-antagonistic fungi. For example, Berg et al. (2005) found Trichoderma and other Verticillium antagonists such as Penicillium and Monographella to vary in diversity and abundance across sites and among host plant (strawberry and oilseed rape) rhizosphere soils. Recently, Taheri et al. (2016) reported that fungal antagonist communities differed depending on the identity of the legume in crop rotation. Unfortunately, understanding of the mechanisms by which plant communities selectively amplify disease-protective fungi does not go beyond single plant species or genotypes (Hartmann et al. 2009). Future work focusing on the role of entire communities would aid in predicting disease outcomes for agroecosystems, which are naturally complex assemblages.
3.3 Disease-suppressive bacteria
Root associated, disease-suppressive bacteria contribute another mechanism by which plants resist disease (Haas and Defago 2005; Sturz and Christie 2003). These rhizosphere bacteria, which include fluorescent pseudomonads, inhibit bacterial and fungal pathogens and even root-feeding nematodes though competition for C, N, and Fe resources as well as through production of antimicrobial compounds such as pyrrolnitrin (PRN) and 2,4-diacytlphloroglucinol (DAPG) (Haas and Defago 2005). Although the mode of action of some of these compounds is not completely understood, both PRN and DAPG appear to interfere with fungal respiration (de Souza et al. 2003; Tripathi and Gottlieb 1969). Like AM fungi, pseudomonads can also offer indirect protection through ISR (van Loon et al. 1998; Haas and Defago 2005).
Plant genotype seems to have an effect on prevalence of DAPG-producing bacteria in rhizosphere soils (Hartmann et al. 2009; Mazzola et al. 2004), and there is evidence linking higher plant diversity with greater abundance of DAPG and PRN producers and disease suppression (Latz et al. 2012), although the specific mechanisms responsible for this effect have not yet been elucidated. Increased abundance of disease-suppressive bacteria is often associated with the onset of disease-suppressive soils after continuous monoculture of wheat (Raaijmakers and Weller 1998). Because pathogen outbreak is required for onset of soil suppressiveness and differences in the level of disease suppression seem to depend on host plant genotype (Mazzola et al. 2004), this phenomenon may involve an interaction between specific plants, fungal pathogens, and resident populations of pseudomonads (Weller et al. 2002). Although many details of bacteria-plant interactions have been studied (see review by Hartmann et al. 2009), the mechanisms by which certain plants tend to increase disease-suppressive bacteria remain in the realm of speculation. Despite this gap in knowledge, promoting disease-suppressive bacteria through the use of certain cover crops appears promising. Further assessment of commonly used cover crop species as well as the effect of plant diversity in driving populations of disease-suppressive bacteria in perennial systems is needed.
3.4 Entomopathogenic fungi
Entomopathogenic fungi (orders Entomophthorales and Hypocreales) are pathogens of many insects, including crop herbivores (Vega et al. 2009). These fungi attack insects by penetrating the exoskeleton with chitin-degrading enzymes, eventually killing them by consuming nutrients in the hemolymph or through release of toxins (Clarkson and Charnley 1996). Because they spend most of their life cycle in the soil, these fungi may be affected by cover crops and help control insect-related problems associated with crop decline. The most well-studied taxa of these entomopathogenic fungi include Beauveria and Metarhizium spp. (Meyling and Eilenberg 2007), both of which have been cultured and sold commercially as biocontrol agents. However, naturally occurring entomopathogens may be effective in reducing insect pests where the habitat encourages their persistence (Meyling and Eilenberg 2007; Pell et al. 2010).
Although their distribution was traditionally thought to be driven by the presence and movement of insect hosts, they are also found as root endophytes (Behie et al. 2015) and show rhizosphere competence (Wyrebek et al. 2011), suggesting that their distribution may depend on plant community identity. This endophytic growth strategy may be key in their additional potential role as fungal pathogen antagonists (Ownley et al. 2008). Although there has been an effort to establish a connection between habitat type and occurrence of these fungi on a landscape scale (Bidochka et al. 2002; Medo and Cagan 2011; Meyling et al. 2011), using cover crops for conservation biocontrol via entomopathogens has seen little experimentation (Meyling and Eilenberg 2007). Vegetation management strategies that encourage these beneficial fungi might also contribute to a reduction in crop decline caused by insect herbivore outbreaks, possibly even supplementing parasitism by beneficial insects.
4 Considerations for managing non-crop vegetation
Manipulating cover crops may be the simplest tool to help mitigate or reverse crop decline issues in perennial systems such as orchards and vineyards as these systems are amenable to manipulation of plant species growing between crop rows. Different parameters, including species diversity, plant identity, and whether the plants are native or exotic, need to be considered when choosing a cover crop mix. Obviously, factors other than vegetation such as soil type, climate, management of water and nutrients, etc. need also to be considered if a certain mix is to be successful. While it is unlikely that there is a one-size-fits-all solution, certain fundamental properties of the new plant community may help in guiding management. The influence of different cover crop types on soil microbes involved in plant-soil feedback is summarized in Table 1.
4.1 Plant diversity
It is well accepted that greater plant diversity leads to a suite of benefits of interest to growers including ecosystem functioning (Bardgett and van der Putten 2014). Reliability of ecosystem services also improves through functional redundancies associated with enhanced diversity (Naeem 1998; van Bruggen et al. 2006). In terms of plant-soil feedbacks specifically, research shows that negative feedback decreases with increasing plant diversity (Maron et al. 2011; Schnitzer et al. 2011) because highly diverse plant communities are less affected by soil-borne pathogens than are monocultures or low-diversity communities (Maron et al. 2011). In perennial systems, the permanent siting of crop plants and the spatial structuring of cover crops between crop rows mean that growers can most easily manipulate plant diversity between rows. Increasing the diversity of cover crop species could enhance heterogeneity of soil microbial communities at multiple spatial scales leading to less negative feedback and decline of crop plants (Fig. 2).
While the overall species richness of cover crop vegetation may be the ultimate driver of soil microbial diversity and, thus, soil feedback (Eisenhauer et al. 2011), plant functional group diversity may also be an important consideration (Milcu et al. 2013). Common cover crop mixtures may consist of a mix of any of the four main (non-woody) functional groups (namely C3 and C4 grasses, legumes, and non-leguminous forbs), and plants with different functional traits often grow well together due to temporal and spatial niche differentiation (Roscher et al. 2013). Similarly, greater diversity of plant functional groups also creates heterogeneity of niches inhabitable by soil microbes. In that case, a cover crop with high plant functional group diversity should promote enhanced microbial diversity and subsequent ecosystem services such as disease suppression. Unfortunately, the effect of cover crop diversity on disease suppression has not yet been studied in perennial systems.
4.2 Considerations within plant functional groups
4.2.1 Legumes
Legumes can have significant effects on many soil microbes in addition to their nitrogen-fixing rhizobial partners. Abundance of AM fungi can be enhanced with the inclusion of legumes due to the high P cost of N2 fixation and, thus, the need for AM symbionts at the nodulation sites of most legumes (Scheublin et al. 2004). The resulting high-quality litter may also attract copiotrophic microbes such as fluorescent pseudomonads (Bastian et al. 2009). However, disease-suppressive bacteria have been shown to decrease when legumes are present in a mix (Latz et al. 2012, 2015) possibly because of plant defense mechanisms, e.g., saponins, which are more commonly found in legumes than most grasses (Osbourn 2003). Further study should focus on other plant-protective secondary metabolites produced by different legumes (Wink 2013) to better understand how they affect relationships with soil microorganisms. This knowledge could aid growers in choosing specific legumes for building beneficial microbial populations in perennial systems.
4.2.2 C3 and C4 grasses
Within grasses, the photosynthetic pathway can also influence plant soil feedbacks. C4 grasses are highly dependent on AM fungi for productivity (Hetrick et al. 1988; Wilson and Hartnett 1998) as they occur primarily in arid, high-light-intensity environments (Sage et al. 2011) where efficient water and P scavenging and uptake are crucial. C3 grasses, on the other hand, are typically adapted to cooler climates or seasons with more available water. Although C3 grasses do form associations with AM fungi, they are less reliant on them for survival due to highly fibrous root systems that are more effective at absorbing nutrients than are the coarser roots of C4 grasses (Hetrick et al. 1988).
C3 grasses may offer other benefits, such as increasing the abundance of disease-suppressive bacteria (Latz et al. 2015) because they may have evolved a greater need for association with protective microorganisms due to their finely branched root systems, which are more susceptible to pathogen attack (Sikes et al. 2009). Variation even at the level of genotype may influence plant soil feedback. Specific genotypes of wheat are associated with different populations of DAPG-producing bacteria in apple replant soils, leading to variation in negative feedback on newly planted trees by pathogens that had built up in the orchard soil (Gu and Mazzola 2003).
Therefore, the inclusion of C4 or C3 grasses in a cover crop mix is context dependent and depends heavily on climate and edaphic conditions: C4 grasses might increase the abundance of AM fungi, while C3 grasses could provide resources for more copiotrophic beneficial microbes such as disease-suppressive bacteria in the cooler months.
4.2.3 Brassicas
Some common cover crops are not mycorrhizal, which may suppress the mycorrhizal community associating with the crop plant (Karasawa et al. 2001; Stinson et al. 2006). For example, most of the Brassicaceae are non-mycorrhizal and inhibit AM fungal spore germination due to the antifungal volatiles (sulfur-containing isothiocyanates) produced by their roots (Schreiner and Koide 1993). Subsequently, these plants may also decrease endemic populations of entomopathogenic fungi, as seen with Alliaria petiolata (garlic mustard) invasion (Keesing et al. 2011). At the same time, this antifungal activity may be beneficial to crops, as brassicas may inhibit fungal pathogens (Walker et al. 1937). Indeed, brassicas are often used as “biofumigants” to kill fungal pathogens and decrease viability of their dormant spores (Smolinska and Horbowicz 1999).
The effects of brassicas are not solely antimicrobial, however, and may confer other benefits. For example, disease-suppressive bacteria have been shown to increase substantially with incorporation of brassicaceous plant material (Hollister et al. 2013) or through rhizosphere selection by living plants (Berg et al. 2005) and may help to alleviate negative feedback as seen with apple replant disease (Mazzola et al. 2015). Whether brassicas in the cover crop are as effective as Brassica plant litter in suppressing fungal pathogens is not clear. While decomposing Brassica litter can dramatically decrease fungal diversity (Hollister et al. 2013; Mazzola et al. 2015), living brassicas may not be broadly anti-fungal as they are susceptible to specific fungal pathogens, e.g., Olpidium brassicae (Bennett et al. 2014). Further, some disease-protective fungi, such as Trichoderma, have been shown to respond positively to both seed meal (Galletti et al. 2008) and living Brassica plants (Kirkegaard et al. 2004). Because much of the research surrounding brassicas has focused on their use as seed meals or green manures and thus is most relevant only to replant situations, future work should investigate their potential to shape microbial communities as living plants within a cover crop system as they are often planted as part of a mix or occur naturally as weedy species.
4.3 Native plant species in cover crops
Plant provenance may be as important as identity in cover crop selection. Native plants, as opposed to exotics or cultivars, may be better adapted to the local climate, requiring fewer inputs when employed as a cover crop (Costello 2010). For example, perennial grasses native to climates that typically experience long dry summers may be effectively dormant during those months and thus provide many ecosystem services associated with cover crops without risk of competition with the crop plant (Costello 2010). Local AM fungal populations may also associate preferentially with native plants compared to exotics (Rúa et al. 2016), leading to a more functional symbiosis and greater inoculum potential.
Whether these benefits transcend across microbial taxa is not clear, because the use of native plants in agricultural settings is not well studied. The most appropriate comparisons contrast microbial communities from neighboring natural grasslands with agronomic fields (Garbeva et al. 2006; Holland et al. 2016; Meyling et al. 2009), which offers some insight into the relationship between plant provenance and soil microbes but is conflated by too many confounding variables. Garbeva et al. (2006) found greater suppression of the generalist pathogen Rhizoctonia solani AG3 in soil collected from a native grassland as compared to soil from the same site that had been converted to agricultural production. Similarly, Meyling et al. (2009) saw greater diversity of the entomopathogenic fungus, B. bassiana, in a natural area bordering a cropping system. The increased fungal diversity and unique AM fungal community seen by Holland et al. (2016) may also have been driven by native plants in the natural sites adjacent to vineyard blocks. Agricultural habitats that more closely resemble the surrounding natural landscape may therefore benefit from the efficiency of co-evolved organisms, but it is unknown if these benefits would remain in the presence of other disturbances associated with agricultural management.
Establishment of native species can be difficult in disturbed settings, and careful management, e.g., mowing frequency and weed management prior to planting, is paramount to successful stands (Penfold and Collins 2012). Knowledge of the native species’ resource requirements (water use, fertility, and soil type preference) and growth habits (germination and flowering time, rhizome formation, etc.) is also helpful for establishment. Although native plants have not typically been used as cover crops in perennial systems, their potential to promote beneficial native soil biota while consuming few resources makes them an attractive option and warrants further investigation.
4.4 Management practices that disrupt cover crop-mediated soil feedbacks
Despite a healthy and diverse cover crop, benefits may be masked or even negated by other management practices. While any practice may alter microbial communities and, thus, ecosystem services provided by a cover crop (including irrigation, fertilization, mowing, compaction, etc.), frequent tillage, herbicide use, and fungicides may have the greatest known potential to negatively affect the soil microbial community in relation to disease outcomes.
4.4.1 Tillage
Perhaps, more than any other practice, tillage negatively affects most soil microbes and their functioning (López-Pineiro et al. 2013; Lupwayi et al. 1998). This largely results from a decrease in soil fungi, especially AM fungi (Brito et al. 2012), through the disruption of mycelial networks and reduced colonization of roots and soil (Jasper et al. 1989). Furthermore, with frequent tillage, the AM fungal community often shifts with selection for species that sporulate efficiently but may invest fewer resources in extra-radical hyphae, perhaps conferring less benefit to host plants (Verbruggen and Kiers 2010).
Bacterial functional diversity may also be greatly reduced through tillage practices (Lupwayi et al. 1998). Although much of the literature on how tillage influences the soil microbial community is based on annual vegetable or grain cropping systems, a causal link exists between maintenance of permanent cover crops and increased soil microbial biomass and diversity in perennial systems such as grape (López-Pineiro et al. 2013) and citrus (Balota and Martins Auler 2011), though implications for disease outcomes have not been explored specifically in this context.
Reduced or no-till cover crop management in perennial systems may benefit from more than just the avoidance of soil disturbance. Because the effect of a plant community on the diversity and functioning of a soil microbial community is more pronounced with time (Eisenhauer et al. 2011), a permanent cover crop may be more advantageous for promoting microbe-mediated ecosystem services to crop plants than the incorporation of annual cover crops or clean cultivation in the long term.
Of course, tillage can be necessary and beneficial in certain instances, e.g., replanting, especially if it helps distribute disease-suppressive pseudomonads built up on the roots of cover crops (Gu and Mazzola 2003). However, the loss of diversity and heterogeneity of the soil microbial community that comes with frequent tillage may be counterproductive in preventing negative feedbacks from dominating in perennial monocultures.
4.4.2 Herbicide use
The herbicide glyphosate, which is ubiquitous in perennial agriculture, can negatively affect beneficial soil microbes. First, it can be directly toxic to microbes, including fungi, bacteria, and algae that use the shikimate pathway for production of aromatic amino acids (Steinrucken and Amrhein 1980). There is evidence that field concentrations of glyphosate have a fungicidal effect against the entomopathogenic fungi B. bassiana and Metarhizium anisopliae (Morjan et al. 2002). Perhaps, most alarming, glyphosate has been shown to affect woody perennials even when applied sparingly as a 1 % solution to bark (Levésque and Rahe 1992), weakening the plant sufficiently to enable damage by weak pathogens such as Cylindrocarpon spp. that were present on roots but kept in check by costly plant defense mechanisms (Levésque and Rahe 1992). This suggests that glyphosate can disrupt the delicate balance of soil feedback and may contribute to decline of crop plants.
Alternative weed management strategies in crop rows include the use of cultivation equipment, flaming, mulching, and seeding living mulches (Krohn and Ferree 2005; Hartwig and Ammon 2002). While some growers take the approach of tolerating weeds in crop rows, this strategy can either be dangerous if weeds harbor generalist pathogens that also affect crop plants (Agustí-Brisach et al. 2011) or advantageous if the weeds are a source of beneficial microbes such as disease-suppressive bacteria (Sturz et al. 2001). The local weed community likely influences these outcomes. Since weed resistance to glyphosate is becoming more common (Ghanizadeh et al. 2015; Koger et al. 2004), repeated use of materials with this mode of action often results in few dominant weed species, which may contribute to decreased soil microbial diversity.
4.4.3 Fungicides
Fungicides are used extensively in many perennial systems to control foliar pathogens such as mildews, and their non-target effects on soil microbes can be significant (Bünemann et al. 2006). Even with woody perennials such as apples or grapes where foliar fungicides are directed up into the canopy, residual material accumulates on the soil surface and has the potential to affect the soil microbial community (Mackie et al. 2012). This is especially problematic for metals such as copper and copper sulfate, which are widely used in both conventional and organically managed systems (Mackie et al. 2012). Heavy metals like copper do not biodegrade and thus accumulate in agricultural soils with a history of copper fungicide use (Wang et al. 2009). Although some microbes can tolerate or adapt to high concentrations of heavy metals (Hassen et al. 1998; Ezzouhri et al. 2009), the reduction in microbial diversity caused by elimination of copper-intolerant species can result in decreased ecosystem functioning in the presence of additional stressors (Tobor-Kaplon et al. 2005).
Overall, copper contamination seems to have a negative effect on total soil microbial biomass and functioning (Fernandez-Calvino et al. 2010; Kandeler et al. 1996) and the potential for inhibiting specific groups of beneficial fungi, such as AM fungi, has also been demonstrated (Graham et al. 1986). Although foliar fungicide treatments are often a necessity in many perennial systems, the material used should be considered carefully as long-term effects on soil microbial functioning could be affected by accumulation of active ingredients in the soil.
5 Conclusions
The lessons learned from ecological studies have much to offer in terms of understanding and alleviating crop decline. These studies show that greater plant diversity suppresses plant disease and promotes increased overall resistance and resilience of the ecosystem. However, many of these results are from highly manipulated, artificial systems or experiments within natural systems. Consequently, these theories need rigorous testing in agricultural settings. Although there has been some success in alleviating replant disease using plant-based strategies, due to the lack of experimentation with long-term cover crop mixes in perennial systems, we can only hypothesize that crop decline could be altogether avoided with a carefully selected, diverse cover crop. Before these ideas can be adopted by growers, it will be necessary to address major gaps in knowledge, including the following: (a) How do the cover crops that we already use affect disease outcomes? (b) Does plant provenance (native vs. exotic) affect plant-soil feedback? and (c) How robust is plant-soil feedback to differences in management regimes? Improving knowledge in this area can help growers take advantage of the effect that plants have in shaping microbial communities by managing non-crop vegetation. Perennial cover crop mixtures with highly diverse, locally adapted plant communities may be best suited if the aim is preventing crop decline caused by negative soil feedback.
References
Agustí-Brisach C, Gramaje D, Leon M, Garcia-Jimenez J, Armengol J (2011) Evaluation of vineyard weeds as potential hosts of black-foot and petri disease pathogens. Plant Dis 95:803–810. doi:10.1094/pdis-12-10-0888
Akiyama K, Matsuzaki K, Hayashi H (2005) Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435:824–827. doi:10.1038/nature03608
Avis TJ, Gravel V, Antoun H, Tweddell RJ (2008) Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity. Soil Biol Biochem 40:1733–1740. doi:10.1016/j.soilbio.2008.02.013
Azcón-Aguilar C, Barea JM (1996) Arbuscular mycorrhizas and biological control of soil-borne plant pathogens—an overview of the mechanisms involved. Mycorrhiza 6:457–464
Badri DV, Vivanco JM (2009) Regulation and function of root exudates. Plant Cell Environ 32:666–681. doi:10.1111/j.1365-3040.2009.01926.x
Balota EL, Martins Auler PA (2011) Soil microbial biomass under different management and tillage systems of permanent intercropped cover species in an orange orchard. Rev Bras Cienc Solo 35:1873–1883
Bardgett RD, Shine A (1999) Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands. Soil Biol Biochem 31:317–321. doi:10.1016/s0038-0717(98)00121-7
Bardgett RD, van der Putten WH (2014) Belowground biodiversity and ecosystem functioning. Nature 515:505–511. doi:10.1038/nature13855
Bastian F, Bouziri L, Nicolardot B, Ranjard L (2009) Impact of wheat straw decomposition on successional patterns of soil microbial community structure. Soil Biol Biochem 41:262–275. doi:10.1016/j.soilbio.2008.10.024
Behie SW, Jones SJ, Bidochka MJ (2015) Plant tissue localization of the endophytic insect pathogenic fungi Metarhizium and Beauveria. Fungal Ecol 13:112–119. doi:10.1016/j.funeco.2014.08.001
Benhamou N, Garand C, Goulet A (2002) Ability of nonpathogenic Fusarium oxysporum strain Fo47 to induce resistance against Pythium ultimum infection in cucumber. Appl Environ Microbiol 68:4044–4060. doi:10.1128/aem.68.8.4044-4060.2002
Bennett AJ, Hilton S, Bending GD, Chandler D, Mills P (2014) Impact of fresh root material and mature crop residues of oilseed rape (Brassica napus) on microbial communities associated with subsequent oilseed rape. Biol Fert Soils 50:1267–1279. doi:10.1007/s00374-014-0934-7
Berg G, Roskot N, Steidle A, Eberl L, Zock A, Smalla K (2002) Plant-dependent genotypic and phenotypic diversity of antagonistic rhizobacteria isolated from different Verticillium host plants. Appl Environ Microbiol 68:3328–3338. doi:10.1128/aem.68.7.3328-3338.2002
Berg G, Zachow C, Lottmann J, Gotz M, Costa R, Smalla K (2005) Impact of plant species and site on rhizosphere-associated fungi antagonistic to Verticillium dahliae Kleb. Appl Environ Microbiol 71:4203–4213. doi:10.1128/aem.71.8.4203-4213.2005
Bever JD (1994) Feedback between plants and their soil communities in an old field community. Ecology 75:1965–1977. doi:10.2307/1941601
Bever JD, Dickie IA, Facelli E, Facelli JM, Klironomos J, Moora M, Rillig MC, Stock WD, Tibbett M, Zobel M (2010) Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol 25:468–478. doi:10.1016/j.tree.2010.05.004
Bever JD, Mangan SA, Alexander HM (2015) Maintenance of plant species diversity by pathogens. Annu Rev Ecol Evol Syst 46:305–325. doi:10.1146/annurev-ecolsys-112414-054306
Bezemer TM, Lawson CS, Hedlund K, Edwards AR, Brook AJ, Igual JM, Mortimer SR, Van der Putten WH (2006) Plant species and functional group effects on abiotic and microbial soil properties and plant-soil feedback responses in two grasslands. J Ecol 94:893–904. doi:10.1111/j.1365-2745.2006.01158.x
Bidochka MJ, Menzies FV, Kamp AM (2002) Genetic groups of the insect-pathogenic fungus Beauveria bassiana are associated with habitat and thermal growth preferences. Arch Microbiol 178:531–537. doi:10.1007/s00203-002-0490-7
Brito I, Goss MJ, de Carvalho M, Chatagnier O, van Tuinen D (2012) Impact of tillage system on arbuscular mycorrhiza fungal communities in the soil under Mediterranean conditions. Soil Tillage Res 121:63–67. doi:10.1016/j.still.2012.01.012
Broeckling CD, Broz AK, Bergelson J, Manter DK, Vivanco JM (2008) Root exudates regulate soil fungal community composition and diversity. Appl Environ Microbiol 74:738–744. doi:10.1128/aem.02188-07
Bunemann EK, Schwenke GD, Van Zwieten L (2006) Impact of agricultural inputs on soil organisms - a review. Austr J Soil Res 44:379–406. doi:10.1071/sr05125
Cameron DD, Neal AL, van Wees SCM, Ton J (2013) Mycorrhiza-induced resistance: more than the sum of its parts? Trends Plant Sci 18:539–545. doi:10.1016/j.tplants.2013.06.004
Cheng XM, Baumgartner K (2004) Arbuscular mycorrhizal fungi-mediated nitrogen transfer from vineyard cover crops to grapevines. Biol Fertil Soils 40:406–412. doi:10.1007/s00374-004-0797-4
Cheng X, Baumgartner K (2006) Effects of mycorrhizal roots and extraradical hyphae on N-15 uptake from vineyard cover crop litter and the soil microbial community. Soil Biol Biochem 38:2665–2675. doi:10.1016/j.soilbio.2006.03.023
Chowdhury N, Marschner P, Burns R (2011) Response of microbial activity and community structure to decreasing soil osmotic and matric potential. Plant Soil 344:241–254. doi:10.1007/s11104-011-0743-9
Civitello DJ, Cohen J, Fatima H, Halstead NT, Liriano J, McMahon TA, Ortega CN, Sauer EL, Sehgal T, Young S, Rohr JR (2015) Biodiversity inhibits parasites: broad evidence for the dilution effect. Proc Natl Acad Sci U S A 112:8667–8671. doi:10.1073/pnas.1506279112
Clarkson JM, Charnley AK (1996) New insights into the mechanisms of fungal pathogenesis in insects. Trends Microbiol 4:197–203. doi:10.1016/0966-842x(96)10022-6
Cornelissen JHC, Thompson K (1997) Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol 135:109–114. doi:10.1046/j.1469-8137.1997.00628.x
Costello MJ (2010) Grapevine and soil water relations with nodding needlegrass (Nassella cernua), a California native grass, as a cover crop. Hortscience 45:621–627
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E (2013) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob Chang Biol 19:988–995. doi:10.1111/gcb.12113
de Souza JT, Arnould C, Deulvot C, Lemanceau P, Gianinazzi-Pearson V, Raaijmakers JM (2003) Effect of 2,4-diacetylphloroglucinol on Pythium: cellular responses and variation in sensitivity among propagules and species. Phytopathology 93:966–975. doi:10.1094/phyto.2003.93.8.966
de Vries FT, Manning P, Tallowin JRB, Mortimer SR, Pilgrim ES, Harrison KA, Hobbs PJ, Quirk H, Shipley B, Cornelissen JHC, Kattge J, Bardgett RD (2012) Abiotic drivers and plant traits explain landscape-scale patterns in soil microbial communities. Ecol Lett 15:1230–1239. doi:10.1111/j.1461-0248.2012.01844.x
Eisenhauer N, Milcu A, Sabais ACW, Bessler H, Brenner J, Engels C, Klarner B, Maraun M, Partsch S, Roscher C, Schonert F, Temperton VM, Thomisch K, Weigelt A, Weisser WW, Scheu S (2011) Plant diversity surpasses plant functional groups and plant productivity as driver of soil biota in the long term. Plos One 6. doi: 10.1371/journal.pone.0016055
Ezzouhri L, Castro E, Moya M, Espinola F, Lairini K (2009) Heavy metal tolerance of filamentous fungi isolated from polluted sites in Tangier, Morocco. Afr J Microbiol Res 3:35–48
Fanin N, Haettenschwiler S, Fromin N (2014) Litter fingerprint on microbial biomass, activity, and community structure in the underlying soil. Plant Soil 379:79–91. doi:10.1007/s11104-014-2051-7
Fanin N, Fromin N, Bertrand I (2016) Functional breadth and home-field advantage generate functional differences among soil microbial decomposers. Ecology 97:1023–1037. doi:10.1890/15-1263.1
Fernandez-Calvino D, Soler-Rovira P, Polo A, Diaz-Ravina M, Arias-Estevez M, Plaza C (2010) Enzyme activities in vineyard soils long-term treated with copper-based fungicides. Soil Biol Biochem 42:2119–2127. doi:10.1016/j.soilbio.2010.08.007
Fierer N, Jackson RB (2006) The diversity and biogeography of soil bacterial communities. Proc Natl Acad Sci U S A 103:626–631. doi:10.1073/pnas.0507535103
Filion M, St-Arnaud M, Fortin JA (1999) Direct interaction between the arbuscular mycorrhizal fungus Glomus intraradices and different rhizosphere microorganisms. New Phytol 141:525–533. doi:10.1046/j.1469-8137.1999.00366.x
Fontaine S, Mariotti A, Abbadie L (2003) The priming effect of organic matter: a question of microbial competition? Soil Biol Biochem 35:837–843. doi:10.1016/s0038-0717(03)00123-8
Galletti S, Sala E, Leoni O, Burzi PL, Cerato C (2008) Trichoderma spp. tolerance to Brassica carinata seed meal for a combined use in biofumigation. Biol Control 45:319–327. doi:10.1016/j.biocontrol.2008.01.014
Garbeva P, van Veen JA, van Elsas JD (2004a) Microbial diversity in soil: selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu Rev Phytopathol 42:243–270. doi:10.1146/annurev.phyto.42.012604.135455
Garbeva P, Voesenek K, van Elsas JD (2004b) Quantitative detection and diversity of the pyrrolnitrin biosynthetic locus in soil under different treatments. Soil Biol Biochem 36:1453–1463. doi:10.1016/j.soilbio.2004.03.009
Garbeva P, Postma J, van Veen JA, van Elsas JD (2006) Effect of above-ground plant species on soil microbial community structure and its impact on suppression of Rhizoctonia solani AG3. Environ Microbiol 8:233–246. doi:10.1111/j.1462-2920.2005.00888.x
Ghanizadeh H, Harrington KC, James TK (2015) Glyphosate-resistant Lolium multiflorum and Lolium perenne populations from New Zealand are also resistant to glufosinate and amitrole. Crop Prot 78:1–4. doi:10.1016/j.cropro.2015.08.008
Gnekow MA, Marschner H (1989) Role of VA mycorrhiza in growth and mineral-nutrition of apple (Malus pumila var domestica) rootstock cuttings. Plant Soil 119:285–293. doi:10.1007/bf02370421
Govaerts B, Mezzalama M, Unno Y, Sayre KD, Luna-Guido M, Vanherck K, Dendooven L, Deckers J (2007) Influence of tillage, residue management, and crop rotation on soil microbial biomass and catabolic diversity. Appl Soil Ecol 37:18–30. doi:10.1016/j.apsoil.2007.03.006
Graham JH, Timmer LW, Fardelmann D (1986) Toxicity of fungicidal copper in soil to citrus seedlings and vesicular-arbuscular mycorrhizal fungi. Phytopathology 76:66–70. doi:10.1094/Phyto-76-66
Gu YH, Mazzola M (2003) Modification of fluorescent pseudomonad community and control of apple replant disease induced in a wheat cultivar-specific manner. Appl Soil Ecol 24:57–72. doi:10.1016/s0929-1393(03)00066-0
Haas D, Defago G (2005) Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat Rev Microbiol 3:307–319. doi:10.1038/nrmicro1129
Hamel C, Vujanovic V, Jeannotte R, Nakano-Hylander A, St-Arnaud M (2005) Negative feedback on a perennial crop: Fusarium crown and root rot of asparagus is related to changes in soil microbial community structure. Plant Soil 268:75–87. doi:10.1007/s11104-004-0228-1
Harman GE, Howell CR, Viterbo A, Chet I, Lorito M (2004) Trichoderma species—opportunistic, avirulent plant symbionts. Nat Rev Microbiol 2:43–56. doi:10.1038/nrmicro797
Hart MM, Reader RJ, Klironomos JN (2003) Plant coexistence mediated by arbuscular mycorrhizal fungi. Trends Ecol Evol 18:418–423. doi:10.1016/s0169-5347(03)00127-7
Hartmann A, Schmid M, van Tuinen D, Berg G (2009) Plant-driven selection of microbes. Plant Soil 321:235–257. doi:10.1007/s11104-008-9814-y
Hartwig NL, Ammon HU (2002) 50th anniversary—invited article—cover crops and living mulches. Weed Sci 50:688–699. doi:10.1614/0043-1745(2002)050[0688:aiacca]2.0.co;2
Hassen A, Saidi N, Cherif M, Boudabous A (1998) Resistance of environmental bacteria to heavy metals. Bioresour Technol 64:7–15. doi:10.1016/s0960-8524(97)00161-2
Hetrick BAD, Kitt DG, Wilson GT (1988) Mycorrhizal dependence and growth habit of warm-season and cool-season tallgrass prairie plants. Can J Bot 66:1376–1380
Hofmann A, Wittenmayer L, Arnold G, Schicber A, Merbach W (2009) Root exudation of phloridzin by apple seedlings (Malus domestica Borkh.) with symptoms of apple replant disease. J Appl Bot Food Qual 82:193–198
Hoitink HAJ, Boehm MJ (1999) Biocontrol within the context of soil microbial communities: a substrate-dependent phenomenon. Annu Rev Phytopathol 37:427–446. doi:10.1146/annurev.phyto.37.1.427
Holland EA, Coleman DC (1987) Litter placement effects on microbial and organic-matter dynamics in an agroecosystem. Ecology 68:425–433. doi:10.2307/1939274
Holland TC, Bowen PA, Bogdanoff CP, Lowery TD, Shaposhnikova O, Smith S, Hart MM (2016) Evaluating the diversity of soil microbial communities in vineyards relative to adjacent native ecosystems. Appl Soil Ecol 100:91–103. doi:10.1016/j.apsoil.2015.12.001
Hollister EB, Hu P, Wang AS, Hons FM, Gentry TJ (2013) Differential impacts of brassicaceous and nonbrassicaceous oilseed meals on soil bacterial and fungal communities. FEMS Microbiol Ecol 83:632–641. doi:10.1111/1574-6941.12020
Hossain MM, Sultana F, Kubota M, Koyarna H, Hyakumachi M (2007) The plant growth-promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant Cell Physiol 48:1724–1736. doi:10.1093/pcp/pcm144
Jasper DA, Abbott LK, Robson AD (1989) Soil disturbance reduces the infectivity of external hyphae of vesicular arbuscular mycorrhizal fungi. New Phytol 112:93–99. doi:10.1111/j.1469-8137.1989.tb00313.x
Kandeler E, Kampichler C, Horak O (1996) Influence of heavy metals on the functional diversity of soil microbial communities. Biol Fertil Soils 23:299–306. doi:10.1007/s003740050174
Karasawa T, Kasahara Y, Takebe M (2001) Variable response of growth and arbuscular mycorrhizal colonization of maize plants to preceding crops in various types of soils. Biol Fertil Soils 33:286–293. doi:10.1007/s003740000321
Keesing F, Belden LK, Daszak P, Dobson A, Harvell CD, Holt RD, Hudson P, Jolles A, Jones KE, Mitchell CE, Myers SS, Bogich T, Ostfeld RS (2010) Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 468:647–652. doi:10.1038/nature09575
Keesing F, Oberoi P, Vaicekonyte R, Gowen K, Henry L, Mount S, Johns P, Ostfeld RS (2011) Effects of garlic mustard (Alliaria petiolata) on entomopathogenic fungi. Ecoscience 18:164–168. doi:10.2980/18-2-3385
Kirkegaard JA, Simpfendorfer S, Holland J, Bambach R, Moore KJ, Rebetzke GJ (2004) Effect of previous crops on crown rot and yield of durum and bread wheat in northern NSW. Aust J Agric Res 55:321–334. doi:10.1071/ar03178
Klabi R, Bell TH, Hamel C, Iwaasa A, Schellenberg M, Raies A, St-Arnaud M (2015) Plant assemblage composition and soil P concentration differentially affect communities of AM and total fungi in a semi-arid grassland. FEMS Microbiol Ecol 91:1–13. doi:10.1093/femsec/fiu015
Koger CH, Poston DH, Hayes RM, Montgomery RE (2004) Glyphosate-resistant horseweed (Conyza canadensis) in Mississippi. Weed Technol 18:820–825. doi:10.1614/wt-03-218r
Krohn NG, Ferree DC (2005) Effects of low-growing perennial ornamental groundcovers on the growth and fruiting of ‘Seyval blanc’ grapevines. Hortscience 40:561–568
Kuan TL, Erwin DC (1982) Effect of soil matric potential on Phytophthora root-rot of alfalfa. Phytopathology 72:543–548. doi:10.1094/Phyto-72-543
Lange M, Habekost M, Eisenhauer N, Roscher C, Bessler H, Engels C, Oelmann Y, Scheu S, Wilcke W, Schulze E-D, Gleixner G (2014) Biotic and abiotic properties mediating plant diversity effects on soil microbial communities in an experimental grassland. Plos One 9. doi: 10.1371/journal.pone.0096182
Larkin RP, Griffin TS, Honeycutt CW (2010) Rotation and cover crop effects on soilborne potato diseases, tuber yield, and soil microbial communities. Plant Dis 94:1491–1502. doi:10.1094/pdis-03-10-0172
Larsen J, Bodker L (2001) Interactions between pea root-inhabiting fungi examined using signature fatty acids. New Phytol 149:487–493. doi:10.1046/j.1469-8137.2001.00049.x
Latz E, Eisenhauer N, Rall BC, Allan E, Roscher C, Scheu S, Jousset A (2012) Plant diversity improves protection against soil-borne pathogens by fostering antagonistic bacterial communities. J Ecol 100:597–604. doi:10.1111/j.1365-2745.2011.01940.x
Latz E, Eisenhauer N, Scheu S, Jousset A (2015) Plant identity drives the expression of biocontrol factors in a rhizosphere bacterium across a plant diversity gradient. Funct Ecol 29:1225–1234. doi:10.1111/1365-2435.12417
Lauber CL, Strickland MS, Bradford MA, Fierer N (2008) The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol Biochem 40:2407–2415. doi:10.1016/j.soilbio.2008.05.021
Levésque CA, Rahe JE (1992) Herbicide interactions with fungal root pathogens, with special reference to glyphosate. Annu Rev Phytopathol 30:579–602. doi:10.1146/annurev.py.30.090192.003051
Loik ME, Breshears DD, Lauenroth WK, Belnap J (2004) A multi-scale perspective of water pulses in dryland ecosystems: climatology and ecohydrology of the western USA. Oecologia 141:269–281. doi:10.1007/s00442-004-1570-y
Long SR (2001) Genes and signals in the Rhizobium-legume symbiosis. Plant Physiol 125:69–72. doi:10.1104/pp.125.1.69
Lopez-Pineiro A, Munoz A, Zamora E, Ramirez M (2013) Influence of the management regime and phenological state of the vines on the physicochemical properties and the seasonal fluctuations of the microorganisms in a vineyard soil under semi-arid conditions. Soil Till Res 126:119–126. doi:10.1016/j.still.2012.09.007
Luongo L, Galli M, Corazza L, Meekes E, De Haas L, Van der Plas CL, Kohl J (2005) Potential of fungal antagonists for biocontrol of Fusarium spp. in wheat and maize through competition in crop debris. Biocontrol Sci Tech 15:229–242. doi:10.1080/09583150400016852
Lupwayi NZ, Rice WA, Clayton GW (1998) Soil microbial diversity and community structure under wheat as influenced by tillage and crop rotation. Soil Biol Biochem 30:1733–1741. doi:10.1016/s0038-0717(98)00025-x
Mackie KA, Mueller T, Kandeler E (2012) Remediation of copper in vineyards—a mini review. Environ Pollut 167:16–26. doi:10.1016/j.envpol.2012.03.023
Maherali H, Klironomos JN (2007) Influence of phylogeny on fungal community assembly and ecosystem functioning. Science 316:1746–1748. doi:10.1126/science.1143082
Maron JL, Marler M, Klironomos JN, Cleveland CC (2011) Soil fungal pathogens and the relationship between plant diversity and productivity. Ecol Lett 14:36–41. doi:10.1111/j.1461-0248.2010.01547.x
Mazzola M, Manici LM (2012) Apple replant disease: role of microbial ecology in cause and control. Annu Rev Phytopathol 50(50):45–65. doi:10.1146/annurev-phyto-081211-173005
Mazzola M, Funnell DL, Raaijmakers JM (2004) Wheat cultivar-specific selection of 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas species from resident soil populations. Microb Ecol 48:338–348. doi:10.1007/s00248-003-1067-y
Mazzola M, Hewavitharana SS, Strauss SL (2015) Brassica seed meal soil amendments transform the rhizosphere microbiome and improve apple production through resistance to pathogen reinfestation. Phytopathology 105:460–469. doi:10.1094/phyto-09-14-0247-r
Medo J, Cagan L (2011) Factors affecting the occurrence of entomopathogenic fungi in soils of Slovakia as revealed using two methods. Biol Control 59:200–208. doi:10.1016/j.biocontrol.2011.07.020
Menge JA, Johnson ELV, Platt RG (1978) Mycorrhizal dependency of several citrus cultivars under three nutrient regimes. New Phytol 81:553–559. doi:10.1111/j.1469-8137.1978.tb01628.x
Meyling NV, Eilenberg J (2007) Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: potential for conservation biological control. Biol Control 43:145–155. doi:10.1016/j.biocontrol.2007.07.007
Meyling NV, Lubeck M, Buckley EP, Eilenberg J, Rehner SA (2009) Community composition, host range and genetic structure of the fungal entomopathogen Beauveria in adjoining agricultural and seminatural habitats. Mol Ecol 18:1282–1293. doi:10.1111/j.1365-294X.2009.04095.x
Meyling NV, Thorup-Kristensen K, Eilenberg J (2011) Below- and aboveground abundance and distribution of fungal entomopathogens in experimental conventional and organic cropping. Biol Control 59:180–186. doi:10.1016/j.biocontrol.2011.07.017
Milcu A, Allan E, Roscher C, Jenkins T, Meyer ST, Flynn D, Bessler H, Buscot F, Engels C, Gubsch M, Koenig S, Lipowsky A, Loranger J, Renker C, Scherber C, Schmid B, Thebault E, Wubet T, Weisser WW, Scheu S, Eisenhauer N (2013) Functionally and phylogenetically diverse plant communities key to soil biota. Ecology 94:1878–1885. doi:10.1890/12-1936.1
Morjan WE, Pedigo LP, Lewis LC (2002) Fungicidal effects of glyphosate and glyphosate formulations on four species of entomopathogenic fungi. Environ Entomol 31:1206–1212. doi:10.1603/0046-225x-31.6.1206
Naeem S (1998) Species redundancy and ecosystem reliability. Conserv Biol 12:39–45. doi:10.1046/j.1523-1739.1998.96379.x
Nicol RW, Yousef L, Traquair JA, Bernards MA (2003) Ginsenosides stimulate the growth of soilborne pathogens of American ginseng. Phytochemistry 64:257–264. doi:10.1016/s0031-9422(03)00271-1
Osbourn AE (2003) Saponins in cereals. Phytochemistry 62:1–4. doi:10.1016/s0031-9422(02)00393-x
Ownley BH, Griffin MR, Klingeman WE, Gwinn KD, Moulton JK, Pereira RM (2008) Beauveria bassiana: endophytic colonization and plant disease control. J Invertebr Pathol 98:267–270. doi:10.1016/j.jip.2008.01.010
Pannkuk CD, Papendick RI, Saxton KE (1997) Fallow management effects on soil water storage and wheat yields in the Pacific Northwest. Agron J 89:386–391
Pell JK, Hannam JJ, Steinkraus DC (2010) Conservation biological control using fungal entomopathogens. Biocontrol 55:187–198. doi:10.1007/s10526-009-9245-6
Penfold C, Collins C (2012) Native cover crops in viticulture. Australian Government/Grape and Wine Research and Development Corporation. http://research.wineaustralia.com/wp-content/uploads/2012/09/2012-05-FS-Native-Cover-Crops.pdf. Accessed 12 June 2016
Pieczarka DJ, Abawi GS (1978) Influence of soil-water potential and temperature on severity of Pythium root-rot of snap beans. Phytopathology 68:766–772
Placella SA, Brodie EL, Firestone MK (2012) Rainfall-induced carbon dioxide pulses result from sequential resuscitation of phylogenetically clustered microbial groups. Proc Natl Acad Sci U S A 109:10931–10936. doi:10.1073/pnas.1204306109
Pozo MJ, Cordier C, Dumas-Gaudot E, Gianinazzi S, Barea JM, Azcón-Aguilar C (2002) Localized versus systemic effect of arbuscular mycorrhizal fungi on defence responses to Phytophthora infection in tomato plants. J Exp Bot 53:525–534. doi:10.1093/jexbot/53.368.525
Raaijmakers JM, Weller DM (1998) Natural plant protection by 2,4-diacetylphloroglucinol—producing Pseudomonas spp. in take-all decline soils. Mol Plant-Microbe Interact 11:144–152. doi:10.1094/mpmi.1998.11.2.144
Rasmann S, Kollner TG, Degenhardt J, Hiltpold I, Toepfer S, Kuhlmann U, Gershenzon J, Turlings TCJ (2005) Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 434:732–737. doi:10.1038/nature03451
Roldán-Fajardo BE, Barea JM (1985) Mycorrhizal dependency in the olive tree (Olea europaea L.). In: Gianinazzi-Pearson V, Gianinazzi S (eds) Physiology and genetical aspects of mycorrhizal. INRA, Paris, pp 323–326
Roscher C, Schumacher J, Lipowsky A, Gubsch M, Weigelt A, Pompe S, Kolle O, Buchmann N, Schmid B, Schulze E-D (2013) A functional trait-based approach to understand community assembly and diversity-productivity relationships over 7 years in experimental grasslands. Perspect Plant Ecol Evol Syst 15:139–149. doi:10.1016/j.ppees.2013.02.004
Rovira AD (1969) Plant root exudates. Bot Rev 35:35. doi:10.1007/bf02859887
Rúa M et al (2016) Home-field advantage? evidence of local adaptation among plants, soil, and arbuscular mycorrhizal fungi through meta-analysis. BMC Evol Biol 16:122. doi:10.1186/s12862-016-0698-9
Sage RF, Christin P-A, Edwards EJ (2011) The C-4 plant lineages of planet Earth. J Exp Bot 62:3155–3169. doi:10.1093/jxb/err048
Scheublin TR, Ridgway KP, Young JPW, van der Heijden MGA (2004) Nonlegumes, legumes, and root nodules harbor different arbuscular mycorrhizal fungal communities. Appl Environ Microbiol 70:6240–6246. doi:10.1128/aem.70.10.6240-6246.2004
Scheublin TR, Sanders IR, Keel C, van der Meer JR (2010) Characterisation of microbial communities colonising the hyphal surfaces of arbuscular mycorrhizal fungi. Isme J 4:752–763. doi:10.1038/ismej.2010.5
Schnitzer SA, Klironomos JN, HilleRisLambers J, Kinkel LL, Reich PB, Xiao K, Rillig MC, Sikes BA, Callaway RM, Mangan SA, van Nes EH, Scheffer M (2011) Soil microbes drive the classic plant diversity-productivity pattern. Ecology 92:296–303. doi:10.1890/10-0773.1
Schreiner RP, Koide RT (1993) Mustards, mustard oils, and mycorrhizas. New Phytol 123:107–113
Schreiner RP, Ivors KL, Pinkerton JN (2001) Soil solarization reduces arbuscular mycorrhizal fungi as a consequence of weed suppression. Mycorrhiza 11:273–277
Schweitzer JA, Bailey JK, Fischer DG, LeRoy CJ, Lonsdorf EV, Whitham TG, Hart SC (2008) Plant-soil-microorganism interactions: heritable relationship between plant genotype and associated soil microorganisms. Ecology 89:773–781. doi:10.1890/07-0337.1
Shapiro-Ilan DI, Gardner WA, Wells L, Wood BW (2012) Cumulative impact of a clover cover crop on the persistence and efficacy of Beauveria bassiana in suppressing the pecan weevil (Coleoptera: Curculionidae). Environ Entomol 41:298–307. doi:10.1603/en11229
Sikes BA, Cottenie K, Klironomos JN (2009) Plant and fungal identity determines pathogen protection of plant roots by arbuscular mycorrhizas. J Ecol 97:1274–1280. doi:10.1111/j.1365-2745.2009.01557.x
Smith SE, Read DJ (2008) Mycorrhizal symbiosis. Academic, New York
Smolinska U, Horbowicz M (1999) Fungicidal activity of volatiles from selected cruciferous plants against resting propagules of soil-borne fungal pathogens. J Phytopathol 147:119–124. doi:10.1111/j.1439-0434.1999.tb03817.x
Steinrucken HC, Amrhein N (1980) The herbicide glyphosate is a potent inhibitor of 5-enolpyruvyl-shikimic-acid 3-phosphate synthase. Biochem Biophys Res Commun 94:1207–1212. doi:10.1016/0006-291x(80)90547-1
Stinson KA, Campbell SA, Powell JR, Wolfe BE, Callaway RM, Thelen GC, Hallett SG, Prati D, Klironomos JN (2006) Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. Plos Biol 4:727–731. doi:10.1371/journal.pbio.0040140
Stirling GR, Smith MK, Smith JP, Stirling AM, Hamill SD (2012) Organic inputs, tillage and rotation practices influence soil health and suppressiveness to soilborne pests and pathogens of ginger. Australas Plant Pathol 41:99–112. doi:10.1007/s13313-011-0096-0
Sturz AV, Christie BR (2003) Beneficial microbial allelopathies in the root zone: the management of soil quality and plant disease with rhizobacteria. Soil Tillage Res 72:107–123. doi:10.1016/s0167-1987(03)00082-5
Sturz AV, Matheson BG, Arsenault W, Kimpinski J, Christie BR (2001) Weeds as a source of plant growth promoting rhizobacteria in agricultural soils. Can J Microbiol 47:1013–1024. doi:10.1139/cjm-47-11-1013
Taheri AE, Hamel C, Gan Y (2016) Cropping practices impact fungal endophytes and pathogens in durum wheat roots. Appl Soil Ecol 100:104–111. doi:10.1016/j.apsoil.2015.12.007
Taylor J, Harrier L (2000) A comparison of nine species of arbuscular mycorrhizal fungi on the development and nutrition of micropropagated Rubus idaeus L. cv. Glen Prosen (red raspberry). Plant Soil 225:53–61. doi:10.1023/a:1026519431096
Tobor-Kaplon MA, Bloem J, Romkens P, de Ruiter PC (2005) Functional stability of microbial communities in contaminated soils. Oikos 111:119–129. doi:10.1111/j.0030-1299.2005.13512.x
Tripathi RK, Gottlieb D (1969) Mechanism of action of antifungal antibiotic pyrrolnitrin. J Bacteriol 100:310
Trouvelot S, Bonneau L, Redecker D, van Tuinen D, Adrian M, Wipf D (2015) Arbuscular mycorrhiza symbiosis in viticulture: a review. Agron Sustain Dev 35:1449–1467. doi:10.1007/s13593-015-0329-7
Úrbez-Torres JR, Haag P, Bowen P, O’Gorman DT (2014) Grapevine trunk diseases in British Columbia: incidence and characterization of the fungal pathogens associated with black foot disease of grapevine. Plant Dis 98:456–468. doi:10.1094/pdis-05-13-0524-re
van Bruggen AHC, Semenov AM, van Diepeningen AD, de Vos OJ, Blok WJ (2006) Relation between soil health, wave-like fluctuations in microbial populations, and soil-borne plant disease management. Eur J Plant Pathol 115:105–122. doi:10.1007/s10658-005-1250-8
van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396:69–72
van der Heijden MGA, Bardgett RD, van Straalen NM (2008) The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296–310. doi:10.1111/j.1461-0248.2007.01139.x
van Elsas JD, Garbeva P, Salles J (2002) Effects of agronomical measures on the microbial diversity of soils as related to the suppression of soil-borne plant pathogens. Biodegradable 13:29–40. doi:10.1023/a:1016393915414
van Loon LC, Bakker P, Pieterse CMJ (1998) Systemic resistance induced by rhizosphere bacteria. Annu Rev Phytopathol 36:453–483. doi:10.1146/annurev.phyto.36.1.453
Vega FE, Goettel MS, Blackwell M, Chandler D, Jackson MA, Keller S, Koike M, Maniania NK, Monzon A, Ownley BH, Pell JK, Rangel DEN, Roy HE (2009) Fungal entomopathogens: new insights on their ecology. Fungal Ecol 2:149–159. doi:10.1016/j.funeco.2009.05.001
Verbruggen E, Kiers ET (2010) Evolutionary ecology of mycorrhizal functional diversity in agricultural systems. Evol Appl 3:547–560. doi:10.1111/j.1752-4571.2010.00145.x
Verbruggen E, Roling WFM, Gamper HA, Kowalchuk GA, Verhoef HA, van der Heijden MGA (2010) Positive effects of organic farming on below-ground mutualists: large-scale comparison of mycorrhizal fungal communities in agricultural soils. New Phytol 186:968–979. doi:10.1111/j.1469-8137.2010.03230.x
Vitale A, Aiello D, Guarnaccia V, Perrone G, Stea G, Polizzi G (2012) First report of root rot caused by Ilyonectria (=Neonectria) macrodidyma on avocado (Persea americana) in Italy. J Phytopathol 160:156–159. doi:10.1111/j.1439-0434.2011.01869.x
Walker JC, Morell S, Foster HH (1937) Toxicity of mustard oils and related sulfur compounds to certain fungi. Am J Bot 24:536–541. doi:10.2307/2437076
Wang Q-Y, Zhou D-M, Cang L (2009) Microbial and enzyme properties of apple orchard soil as affected by long-term application of copper fungicide. Soil Biol Biochem 41:1504–1509. doi:10.1016/j.soilbio.2009.04.010
Weller DM, Raaijmakers JM, Gardener BBM, Thomashow LS (2002) Microbial populations responsible for specific soil suppressiveness to plant pathogens. Ann Rev Phytopathol 40:309–348. doi:10.1146/annurev.phyto.40.030402.110010
Wilson GWT, Hartnett DC (1998) Interspecific variation in plant responses to mycorrhizal colonization in tallgrass prairie. Am J Bot 85:1732–1738. doi:10.2307/2446507
Wink M (2013) Evolution of secondary metabolites in legumes (Fabaceae). S Afr J Bot 89:164–175. doi:10.1016/j.sajb.2013.06.006
Wyrebek M, Huber C, Sasan RK, Bidochka MJ (2011) Three sympatrically occurring species of Metarhizium show plant rhizosphere specificity. Microbiol-Sgm 157:2904–2911. doi:10.1099/mic.0.051102-0
Acknowledgments
The authors wish to acknowledge funding sources that made this work possible. EV was supported by the British Columbia Wine Grape Council and the Growing Forward program of Agriculture and Agri-food Canada. MH was supported by the Organic Science Cluster/Growing Forward program of Agriculture and Agri-food Canada.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Vukicevich, E., Lowery, T., Bowen, P. et al. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 36, 48 (2016). https://doi.org/10.1007/s13593-016-0385-7
Accepted:
Published:
DOI: https://doi.org/10.1007/s13593-016-0385-7