
Abstract
Symbiotic cellulolytic microcosms represent a promising microbial agent for enhancing degradation of cellulosic materials in biotechnological processes. In this study, structurally stable lignocellulose-degrading microbial communities were constructed from cellulolytic seed culture from sugarcane bagasse compost using swine manure and Napier grass as co-digested carbon sources under static aerobic condition at 55 °C. The lignocellulolytic microbial consortium enriched in peptone and yeast extract-based medium (PLMC) showed higher cellulose-degrading activity compared to lignocellulolytic microbial consortium isolated in the water-based medium (WLMC). The composite microbes in both consortia were originated from the seed culture and the co-digested substrates according to denaturing gradient gel electrophoresis profile. PLMC exhibited higher CMCase, xylanase, FPase, and avicelase activities in the supernatant than those of WLMC and was capable of degrading 70 % of filter paper within 1 week. PLMC was capable of degrading substrate with higher efficiency than the control by 11.7, 9.2, 15.5, and 11.9 % of total solid, suspended solid, volatile solid, and volatile suspended solid, respectively. The work demonstrated the potential of cellulolytic microcosms enriched by this approach on enhancing conversion efficiency in biogas production from cellulosic wastes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Lignocellulose, the main composition of the plant cell wall, is a complex mixture of polysaccharides by the highly crystalline and ordered cellulose fibers, connected to matrix of the amorphous hemicellulose such as xylan, galactomannan, pectic substances, and other polysaccharides that shielded by the rigid lignin, making it highly recalcitrant to physical, chemical, and biological attacks. The efficient conversion of lignocellulose by enzymes and/or microorganisms would be highly necessary. Hydrolysis of lignocellulosic fibrous component is usually the rate-limiting step in anaerobic digestion, resulting in low methane yield and conversion rate from wastes rich in plant fiber [1]. In order to improve economic viability of the anaerobic digestion process, several approaches including physical, chemical, thermochemical, and biological pretreatments have been studied for increasing susceptibility of lignocellulose to microbial digestion which result in greater overall methane yield [2].
Enhancement of bio-digestibility of lignocellulosic wastes by microbial processes is a promising strategy due to its simplicity, low capital cost, and low consumption of energy and chemicals with competitive economic feasibility [3]. Degradation of lignocelluloses in nature involves synergistic and cooperative actions of various cellulolytic, hemicellulolytic, and lignolytic microorganisms [4], particularly by the role of anaerobic bacteria under anoxic conditions. Several researchers have reported the construction of symbiotic cellulolytic microbial consortia which reduced community complexity compared to the ones existed in nature by successive sub-cultivation of target cellulosic substrates under mesophilic and thermophilic conditions. These microbial consortia were capable of degradation of their respective cellulosic wastes including rice straw [5], sugarcane bagasse [6], and switch grass [7]. The applicability of cellulolytic microbial consortia on enhancing efficiency in anaerobic digestion efficiency has been demonstrated in recent years with the uses of various fiber-rich substrates such as cassava residue [8], paper factory wastes [9], wheat straw [10], and rice straw [11], resulting in significant increase in biogas yield.
Biogas production from swine manure is widely used in small- and large-scale farms as an economically feasible approach for waste treatment and generation of heat and power for local uses. The use of active cellulolytic consortium on enhancing anaerobic digestion of the fibrous fraction in swine manure, resulting in increasing biogas conversion yield and rate along with improving cellulose and hemicellulose removal efficiencies, has been reported [11, 12]. Co-digestion of swine manure with other cellulosic materials, for example with various energy crop residue, has also received an increasing attention with advantages on reducing nitrogen toxicity from swine manure while increasing organic loading in the system [13, 14]. Napier grass (Pennisetum purpureum) is a fast-growing energy crop which has been introduced for biogas production in several countries as a co-digested or main substrate in anaerobic digestion processes [15]. Compared to other energy crop, Napier grass contains higher essential nutrients to support microbial growth with high biomass productivity at relatively inexpensive cost [16]. In order to increase efficiency on bio-methane production from swine manure co-digested with Napier grass, a thermophilic cellulolytic microcosm was isolated using swine manure and napier grass as the major carbon and nitrogen sources in water-based and synthetic medium-based media, resulting in the construction of a highly efficient cellulolytic consortium which can co-exist with native microflora in the waste substrates which solves the problems related to stability of introduced cellulolytic microcosms previously constructed in synthetic complex media [12]. To our knowledge, there has been no reported co-digestion between swine manure and napier grass. Therefore, the microcosms were constructed and characterized for their community structure by using denaturing gradient gel electrophoresis (DGGE) and enzymatic activity profiles and showed high efficiency on degradation of the co-digested cellulosic substrates.
2 Materials and methods
2.1 Inoculum seed and substrate preparation
Swine manure was collected from a farm in Ratchaburi province, Thailand. The total solid (TS) and volatile solid (VS) of swine manure were 250 ± 2 and 186 ± 1 g kg−1 fresh, respectively, according to standard method for the Examination of Water and Wastewater [17]. Napier grass was supplied by UAC Public Company Limited (Chiang Mai, Thailand). The Napier grass sample contained 320 ± 3 and 293 ± 2 g kg−1 of TS and VS, respectively. The Napier grass was cut and sieved through 0.5 mm sieved by an ultracentrifuge mill machine (ZM 200, Retsch, Dusseldorf, Germany) before use. The swine manure and Napier grass samples were used with no sterilization. The microbial seed samples were randomly collected from decayed bagasse compost at Eastern Sugar and Cane PLC in Sa Kaeo province, Thailand. All samples were kept at 4 °C.
2.2 Isolation of lignocellulolytic microbial consortium
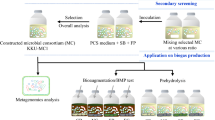
The symbiotic cellulytic microcosms were isolated using sugarcane bagasse compost as the seed cultures according to the method modified from Haruta et al. [5]. One gram of the bagasse compost was inoculated into 40 ml of distilled water or PCS medium (0.1 % yeast extract, 0.2 % CaCO3, 0.5 % peptone, 0.5 % NaCl) containing 2 g fresh swine manure and 2 g of napier grass without sterilization of the substrates in a 50-ml screw-cap tubes with a Whatman no. 1 filter paper strip (1 × 10 cm) (Whatman, Maidstone, Kent, UK) as an indicator for cellulase activity. The mixture was incubated at 55 °C under static condition. When the filter paper strip disintegrated (approximately 4–10 days), 4 ml (10 % of total volume) of the culture was then transferred into fresh medium. The procedure was repeated until a structurally stable cellulolytic microbial community was obtained as indicated by DGGE analysis. The lignocellulosic microbial consortium was maintained by continual sub-cultivation as described above or in sterile water with 20 % glycerol with no cellulosic substrate at −80 °C for long-term storage. The structurally stable consortium was used for all subsequent experimental studies. The structurally stable lignocellulosic microbial consortia grown in water or PCS-based media were designed as WLMC and PLMC, respectively.
2.3 Denaturing gradient gel electrophoresis of 16S rRNA fragments analysis
Total genomic DNA was extracted from the selected lignocellulolytic microbial community by the method described by Zhou et al. [18]. The purified DNA was used as a template for amplification of the partial 16S rDNA fragment using Taq DNA polymerase (Fermentas, Vilnius, Lithunia) according to the manufacturer’s protocol with 338GC-F forward primer, which was attached to a GC clamp at the 5′-terminus (5′-cgcccgccgcgcgcggcgggcggggcgggggcacggggggactcctacgggaggca-3′, GC clamp sequence is underlined), and 518R reverse primer (5′-attaccgcggctgctgg-3′). Reactions were performed in a MyCycler thermal cycler (Bio-Rad Laboratories, Hercules, CA). The temperature profile consisted of 94 °C for 3 min, followed by 30 cycles of denaturation at 94 °C for 1 min, annealing at 55 °C for 1 min, and extension at 72 °C for 2 min, followed by a final extension step at 72 °C for 3 min. The profiles of amplified 16S rDNA fragments were analyzed by DGGE on the DCode™ system (Bio-Rad) according to Wongwilaiwalin et al. [6].
2.4 Cellulosic substrate degradation efficiency
The efficiency of cellulolytic microcosms on cellulosic substrate degradation was assessed by inoculating 4 ml of the lignocellulosic microbial consortia cultivated for 7 days into 40 ml of the respective fresh medium containing the cellulosic substrates and a filter paper strip in a 50-ml screw-cap tube as described above. All cultures were incubated under static condition at 55 °C for 7 days. The supernatant fractions after cell pelleting were analyzed for TS, VS, suspended solid (SS), and volatile suspended solid (VSS) removal efficiency using a standard method [17].
2.5 Enzyme activity analysis
The culture supernatants of lignocellulosic microbial consortia and their controls taken at 7 days of cultivation were clarified by centrifugation at 7500×g for 15 min and used for enzyme profile analysis. Lignocellulose-degrading activities in the supernatant were analyzed based on the amount of liberated reducing sugars using the 3,5-dinitrosalisylic acid (DNS) method as described in Rattanachomsri et al. [19]. One hundred microliter reactions contained the appropriate dilution of enzyme in 50 mM sodium phosphate buffer, pH 7.0, and 0.5 % (w/v) of the corresponding substrate: carboxymethyl cellulose for CMCase activity, microcrystalline cellulose Avicel® for avicelase activity, and beechwood xylan for xylanase activity. The reaction was incubated at 60 °C for 10 min and the amount of reducing sugars was determined from the absorbance measurements at 540 nm and interpolated from a standard curve of the corresponding sugars. The filter paper (FPase) activity was assayed using a 1 × 1 cm (50 mg) Whatman filter paper no. 1 as the substrate as described above and incubated at 60 °C for 4 h. Assay reactions were performed in triplicates and the mean was reported with standard deviation ≤5 %. One international unit (IU) was defined as the amount of enzyme which produced 1 μmol of reducing sugar in 1 min.
3 Results and discussion
3.1 Lignocellulolytic microbial consortium isolation
Structurally and functionally stable cellulolytic microcosms originating from microflora in bagasse compost were obtained by successive sub-cultivation on swine manure and milled Napier grass under facultative anoxic conditions. After several rounds of sub-cultivations, the most active consortia of PLMC and WLMC were selected as a representative from each line based on their cellulose-degrading capability. According to the DGGE patterns, the community structures of both consortia tended to stabilize after 7–12 rounds of successive sub-cultivations with a typical incubation time of 4–7 days for PLMC and 7–10 days for WLMC (Fig. 1) for each batch cycle as observed from >70 % filter paper disintegration. The sub-cultivation procedure can be continuously repeated for more than 30 generations in 9 months with no significant loss in functional activity.

DGGE profiles of lignocellulosic microbial consortium from WLMC profiles between 7th and 11th generations with indicated band nos. 1–8, WC was native background microbe in distilled water (a); PLMC profiles between 8th and 12th generations with indicated band nos. 1–9, PC was native background microbe in PCS medium (b)
PLMC and WLMC showed differences in their 16S rRNA gene patterns according to DGGE analysis. Their community profiles were also markedly different to those of water-base medium without seed consortium (WC) and PCS-based media without seed consortium (PC) as controls which contained only the native microflora in swine manure and napier grass, suggesting modification of the native microbial community structures in the substrates by the seed cultures from bagasse compost. The DGGE pattern of WLMC contained eight major bands while band no. 5 was the most dominated one based on the band’s intensity (Fig. 1a). Band nos. 7 and 8 were found conserved between WLMC and the control WC suggesting that they represented native bacteria in the co-digested substrates that can co-exist with the bacteria in cellulolytic seed cultures. The DGGE pattern of PLMC contained nine major bands (Fig. 1b). Compared with the PC control, band nos. 3, 4, 6, 8, and 9 were suggested to represent the bacteria originated from the substrates that can co-exist with the ones from bagasse compost (bands 1 and 2). The more complex band patterns of PLMC compared to WLMC strongly suggested that the enrichment of more diverse bacteria due to nutrient in the PCS medium. As no significant changes were observed during the sub-cultivation of WLMC (7th–11th generations) and PLMC (8th–12th generations), the results thus indicate structural stabilities of the microcosms comprising of composite microbes from the seed cellulolytic culture in the background of co-existed microflora in the native co-digested substrates due to the selective effects from carbon/nitrogen sources, temperature, and oxygen availability during the sub-cultivation process. Interestingly, several reports to date have demonstrated the successful anaerobic co-digestion of livestock wastes and lignocelluloses such as swine manure and rice straw [20], chicken manure and corn stover [21], chicken manure with coconut/coffee grounds/cassava [22], dairy manure, and wheat straw [14]. However, none has reported the co-digestion between swine manure and Napier grass.
Many previous studies showed the successful construction of structurally and functionally stable cellulolytic microbial consortia from rice straw [5] and switch grass [7] and obtained stable communities within approximately ten rounds of sub-cultivation. These microbial consortia isolated under mesophilic or thermophilic temperatures under facultative anoxic conditions contained clostridia as the major cellulolytic microbes together with various co-existed non-cellulolytic facultative bacteria, for example beta-proteobacterium and Rhodocyclaceae which have roles on consuming oxygen in the system to generate anaerobic conditions to promote the growth of cellulolytic clostridia [8]. Their metabolic roles on controlling pH and removal of inhibitors were also demonstrated [23].
3.2 Cellulosic substrate degradation efficiency analysis
Degradation efficiency of the co-digested swine manure and Napier grass substrates by the PLMC and WLMC was evaluated in comparison to the controls containing swine manure and Napier grass in PC (PCS-based medium) and WC (water-based medium). After 7 days of incubation, the samples were collected and analyzed for the remaining weight of the substrates (Fig. 2). The WLMC consortium showed no significant degradation efficiency compared with its WC (as control). While the PLMC was capable of degrading substrate with higher efficiency than the control PC by 11.7, 9.2, 15.5, and 11.9 % of TS, SS, VS, and VSS, respectively, this result proved that PLMC had the ability to destruct the mixed substrate within the first week much better than the control culture containing only native microflora in the co-digested substrates. The lower activity of WLMC could be due to the lower nutrient content in the media which could lead to less active cellulolytic microcosms. Variation in lignocellulosic substrate degradation efficiency was in the range of 30–60 % after 7 days of incubation using different cellulolytic microbial consortia on several agricultural wastes, e.g., bagasse cane, corn stover, rice straw, and paper pulp waste [6]. The ratio of inoculum volume to substrate also showed effects and correlation with substrate degradation efficiency [12]. The optimum inoculum volume of the microcosms to the substrates could be further studied to increase their lignocellulose degradation efficiency for further application in biotechnological processes.

The cellulosic substrate degradation efficiency analysis of WC, WLMC, PC, and PLMC that determined total solid, suspended solid, volatile solid, and volatile suspended solid at 7 days of incubation culture
3.3 Lignocellulolytic enzyme activity profiles
Lignocellulolytic enzyme activities comprising of CMCase, xylanase, FPase, and avicelase were examined in the supernatant fraction of the microcosms at 7 days of incubation (Fig. 3). The FPase and avicelase from PLMC and WLMC were not significantly different to those of the controls. Xylanase activity from WLMC (0.07 IU/ml) was nearly equal to WC control, whereas that of PLMC (0.12 IU/ml) was significantly higher than that of PC and also slightly higher than that of WLMC. Both microcosms showed higher CMCase activity compared with its control. The highest CMCase activity of 0.12 IU/ml was found for PLMC. The trend of enzyme profile from previous lignocellulosic microbial consortia demonstrated that the CMCase activity slightly increased then maintained after 1–2 days after seed, inoculation while the xylanase activity showed higher activity compared to the cellulase activities, which is higher than CMCase activity, and then decreased during the late phase of batch cycle [6, 8].

Lignocellulosic enzyme activities of WC, WLMC, PC, and PLMC at 7 days of incubation culture
4 Conclusion
The structurally stable symbiotic cellulolytic microbial microcosms were constructed in this study. The consortia comprised composite microbes from both the seed cellulolytic culture co-existed with the microflora from the co-digested substrates. This study is the first report of co-digestion between swine manure and Napier grass. The results suggested the potential of using the cellulolytic microcosms for enhancing degradation of cellulosic materials in anaerobic digestion. Moreover, the microcosms developed in this study could be potent candidates for further application on enhancing degradation of cellulosic substrates in anaerobic digestion and other biotechnological processes.
References
Vavilin VA, Fernandez B, Palatsi J, Flotats X (2008) Hydrolysis kinetics in anaerobic degradation of particulate organic material: an overview. Waste Manag 28:939–951. doi:10.1016/j.wasman.2007.03.028
Sun Y, Cheng J (2002) Hydrolysis of lignocellulosic materials for ethanol production: a review. Bioresource Technol 83:1–11
Recktenwald M, Wawrzynczyk J, Dey ES, Norrlöw O (2008) Enhanced efficiency of industrial-scale anaerobic digestion by the addition of glycosidic enzymes. J Environ Sci Heal A Tox Hazard Subst Environ Eng 43:1536–1540. doi:10.1080/10934520802293693
Howard RL, Abotsi E, Jansen van Rensburg EL, Howard S (2003) Lignocellulose biotechnology: issues of bioconversion and enzyme production: review. Afr J Biotechnol 2:602–619
Haruta S, Cui Z, Huang Z, Li M, Ishii M, Igarashi Y (2002) Construction of a stable microbial community with high cellulose-degradation ability. Appl Microbiol Biotechnol 59:529–534. doi:10.1007/s00253-002-1026-4
Wongwilaiwalin S, Rattanachomsri U, Laothanachareon T, Eurwilaichitr L, Igarashi Y, Champreda V (2010) Analysis of a thermophilic lignocellulose degrading microbial consortium and multi-species lignocellulolytic enzyme system. Enzyme Microb Tech 47:283–290. doi:10.1016/j.enzmictec.2010.07.013
Yang HY, Wu H, Wang XF, Cui ZJ, Li YH (2011) Selection and characteristics of a switchgrass-colonizing microbial community to produce extracellular cellulases and xylanases. Bioresource Technol 102:3546–3550. doi:10.1016/j.biortech.2010.09.009
Zhang Q, He J, Tian M, Mao Z, Tang L, Zhang J, Zhang H (2011) Enhancement of methane production from cassava residues by biological pretreatment using a constructed microbial consortium. Bioresource Technol 102:8899–8906. doi:10.1016/j.biortech.2011.06.061
Yuan X, Cao Y, Li J, Wen B, Zhu W, Wang X, Cui Z (2012) Effect or pretreatment by a microbial consortium on methane production of waste paper and cardboard. Bioresource Technol 118:281–288. doi:10.1016/j.biortech.2012.05.058
Wen B, Yuan X, Cao Y, Liu Y, Wang X, Cui Z (2012) Optimization of liquid fermentation of microbial consortium WSD-5 followed by saccharification and acidification of wheat straw. Bioresource Technol 118:141–149. doi:10.1016/j.biortech.2012.05.025
Yan L, Gao Y, Wan Y, Liu Q, Sun Z, Fu B, Wen X, Cui Z, Wang W (2012) Diversity of a mesophilic lignocellulolytic microbial consortium which is useful for enhancement of biogas production. Bioresource Technol 111:49–54. doi:10.1016/j.biortech.2012.01.173
Tuesorn S, Wongwilaiwalin S, Champreda V, Leethochawalit M, Nopharatana A, Techkarnjanaruk S, Chaiprasert P (2013) Enhancement of biogas production from swine manure by a lignocellulolytic microbial consortium. Bioresource Technol 144:579–586. doi:10.1016/j.biortech.2013.07.013
Cuetos MJ, Fernandez C, Gomez X, Moran A (2011) Anaerobic co-digestion of swine manure with energy crops. Biotechnol Bioprocess Eng 16:1044–1052
Wang X, Yang Y, Feng Y, Ren G, Han X (2012) Optimizing feeding composition and carbon–nitrogen ratios for improved methane yield during anaerobic co-digestion of dairy, chicken manure and wheat straw. Bioresource Technol 120:78–83. doi:10.1016/j.biortech.2012.06.058
Sawasdee V, Pisutpaisalb N (2014) Feasibility of biogas production from napier grass: The 6th International Conference on Applied Energy—ICAE2014. Energy Procedia 61:1229–1233. doi:10.1016/j.egypro.2014.11.1064
Okaraonye CC, Ikewuchi JC (2009) Nutritional and antinutritional components of Pennisetum purpureum (Schumach). Pak J Nutr 8:32–34. doi:10.3923/pjn.2009.32.34
APHA (2005) Standard methods for the examination of water and wastewater. American Public Health Association, Washington, DC
Zhou J, Bruns MA, Tiedje JM (1996) DNA recovery from soils of diverse composition. Appl Environ Microb 62:316–322
Rattanachomsri U, Tanapongpipat S, Eurwilaichitr L, Champreda V (2009) Simultaneous non-thermal saccharification of cassava pulp by multi-enzyme activity and ethanol fermentation by Candida tropicalis. J Biosci Bioeng 107:488–493. doi:10.1016/j.jbiosc.2008.12.024
Ye J, Li D, Sun Y, Wang G, Yuan Z, Zhen F, Wang Y (2013) Improved biogas production from rice straw by co-digestion with kitchen waste and pig manure. Waste Manag 33:2653–2658. doi:10.1016/j.wasman.2013.05.014
Li, Y, Zhang, R, Chen, C, Liu, G, He, Y, Liu, X (2013) Biogas production from co-digestion of corn stover and chicken manure under anaerobic wet, hemi-solid, and solid state conditions. Bioresource Technol 149: 406–412. doi.org/10.1016/j.biortech.2013.09.091
Abouelenien, F, Namba, Y, Kosseva, MR, Nishio, N, Nakashimada, Y (2014) Enhancement of methane production from co-digestion of chicken manure with agricultural wastes. Bioresource Technol 159: 80–87. doi.org/10.1016/j.biortech.2014.02.050
Kato S, Haruta S, Cui ZJ, Ishii M, Igarashi Y (2005) Stable coexistence of five bacterial strains as a cellulose degrading community. Appl Environ Microb 71:7099–7106
Acknowledgments
This study was financially supported by The Joint Graduate School of Energy and Environment (JGSEE) at King Mongkut’s University of Technology Thonburi (KMUTT).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wongwilaiwalin, S., Mhuantong, W., Tangphatsornruang, S. et al. Isolation of cellulolytic microcosms from bagasse compost in co-digested fibrous substrates. Biomass Conv. Bioref. 6, 421–426 (2016). https://doi.org/10.1007/s13399-016-0199-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13399-016-0199-5