
Abstract
Little information exists on the nesting of hedgehogs (Erinaceus europaeus) in the harsh northern environment, although this aspect has specific importance for the species’ survival in this region. We studied the nest types and nesting site selection of radio tagged hedgehogs in urban areas in Eastern Finland. Altogether, four different nest types were discovered and one of them (the pre-hibernation nest) is described here for the first time. Hedgehogs preferred residential areas for nesting during the mating and post-mating seasons while in the pre-hibernation season and hibernation period nesting was more common in forest areas. Nest constructions were closely related to materials found in the nest vicinity. During the active period, males used on average 24 nests and changed them 30 times, whereas females used 17 nests and changed them 26 times. The number of female nests and frequency of nest change increased from post-mating to pre-hibernation seasons, while in males both decreased. In general, female and male nests were situated within their established long-term home range, but in the pre-hibernation season over 30 % of male nests were situated outside of it. In post-mating and pre-hibernation seasons, females avoided other individuals of the same sex; this was demonstrated by the lack of nests in other female core areas. Hedgehogs hibernate for around 223 days, which is the longest reported hibernation period for this species. The results of the present study emphasise the importance of forest and diverse residential areas including forest patches for the hedgehog’s nesting habitat.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nests are ecologically very important for many small mammals, offering concealment and protection from predators, thermal insulation and a place for nursing, especially in harsh climates (e.g., Vogt and Lynch 1982; Redman et al. 1999; Hanski et al. 2000). Nests are of great importance for solitary and nocturnal hedgehogs (Erinaceus europaeus), but unlike many other mammals, which maintain a secure permanent nest, the majority of hedgehog nests are designed only for short-term occupancy (Morris 1973). During the summer, nests are used as a daytime retreat and, for females with litters, as a refuge (Reeve 1994). In winter hedgehog spend periods of up to several months hibernating in a nest. These nests are constructed more thoroughly than day nests, which are often flimsy structures (Morris 1973). Both the available materials and location have an effect on the quality of the nest (Morris 1973; Boitani and Reggiani 1984; Huijser 2000). Usually, the nests are situated above ground under bushes or dense shrubs, in grasses and sometimes in burrows (Reeve and Morris 1985; Huijser 2000; Haigh et al. 2012).
The most common nesting habitat is a hedgerow or forest (Huijser 2000; Riber 2006; Haigh et al. 2012) and forest is especially preferred for hibernation (Huijser 2000; Jensen 2004). Across its home range, the hedgehog has several day nests, which are changed with varying intensity (Reeve and Morris 1985). Day nests may also be abandoned and later reused, for example, as hibernation nests (Boitani and Reggiani 1984; Reeve and Morris 1985; Haigh et al. 2012). Adults have rarely been observed using the same nest simultaneously (Morris 1973), but the sequential use of a nest by multiple individuals has been reported (Reeve and Morris 1985; Riber 2006; Haigh et al. 2012). In Northern Europe, hedgehogs can hibernate for over 200 days from August–October until April–May, but in milder areas, the hibernation period is more variable, and some individuals may remain active almost all year round (Parkes 1975; Jensen 2004; Haigh et al. 2012; Rautio et al. 2013).
The European hedgehog has a fairly wide geographical distribution and occurs in a great variety of climatic conditions. In Fennoscandia, the species lives at the northernmost limit of its distribution range. All important activities such as breeding and rearing young, foraging for maintenance and fat storing for hibernation have to be done in a relatively short period, which is challenging for juveniles in particular. The behaviour of a species at the northern edge of its range is likely to differ from that observed in southern areas, where the available habitats and climate are different. Knowledge of hedgehog behavioural ecology in Northern Europe is fairly scarce (Kristiansson 1984; Rautio et al. 2013), and there is no information on the nesting of hedgehogs, although this aspect is of specific importance for species survival in a harsh environment. In addition, a significant decline in hedgehog population has been evidenced in recent years in Europe (Hof 2009), and an understanding of the space use patterns of the species is an important prerequisite for conservation.
Since hedgehogs have large home ranges in harsh climate (Rautio et al. 2013), the current study hypothesised that variations would be apparent also in the nesting behaviour and in number of nests. In this work, as a contribution to the knowledge of the nesting ecology of hedgehogs, we examine the nest function and structure and nest habitat selection in urban areas in the northern edge of hedgehog range. More specifically, we study the differences between males and females in sequential nest use, frequency of resting without nests and how nest habitat selection differs between nest types. We hypothesise that since male hedgehogs have generally larger home ranges than female hedgehogs (Rautio et al. 2013), males also have a greater number of nests and they change nests more frequently. We also address the spatial distribution of nests in a home range and how it changes during the active period and between sexes. Since the home ranges of individuals of both sexes overlap, but females share core areas with other females only during the mating season (Rautio et al. 2013), we expect that the proportion of nests in overlapping areas changes during the active period, but the change is different among the sexes.
Materials and methods
Study area
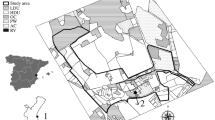
The research was conducted in Joensuu (63°N, 29°E) a medium-sized town in Eastern Finland with a population of 73,000. The study area (about 7 km2) consisted of the three main habitat classes: (1) constructed areas (48 %) including the city centre, residential and facility areas, (2) other constructed areas (29 %) including leisure activity areas and road areas and (3) forest and semi-natural areas (23 %) including deciduous, mixed, coniferous and transitional woodland and scrub (see Fig. 1). The research was undertaken between spring 2004 and early summer 2006. The ground was covered with snow for about 5 months (from the end of November to the end of April), with the average snow depth in the study area in January being 36 cm (ranging from 21 to 70 cm).

The proportion of a habitats in the study area and b hedgehog nest types with c number of nests by location in specific habitats. The residential area and leisure activity area included small patches of forest (17 % and 3 % of the total area, respectively)
Radio tracking
Adult hedgehogs (13 males, 12 females) were tracked by VHF radio telemetry (see details in Rautio et al. 2013; Appendix). The number and spatial distribution of nests, nest usage patterns and nest habitat selection of hedgehogs was monitored during active period, from spring (April) until hibernation commenced in autumn (October) annually. Home range analyses of the tracked individuals (Rautio et al. 2013) were used to estimate the spatial distribution of nests. In autumn, the radio tracked hedgehogs (five females, six males) were considered to have entered hibernation if they did not leave their nest for a month. Their wintertime location was checked at monthly intervals. Research activities were undertaken according to the regulations of University of Joensuu Ethical Commission on Animal Experiments. A license for live capturing and tagging hedgehogs in the field was granted by the North Karelia Environment Centre.
The hedgehog is a nocturnal species, so it can be expected to stay stationary throughout the day also during its active period. This allowed the authors to locate the nest sites between dawn and dusk. Using a handheld radio receiver, it was possible to find the exact location of a nest occupied by a tagged individual. The observed nests were marked in the terrain by tying an inconspicuous string with a unique identification code nearby. The code revealed the date when the nest was first observed and the identity of the individual occupying the nest during the first observation.
Nest types were divided into four categories based on their function: (1) day nests, (2) breeding nests, (3) pre-hibernation nests and (4) hibernation nests. Day nests were used as a daytime retreat during the hedgehogs’ active period. Breeding nests were used during the nursing period (from the end of June until the end of July) until juveniles were weaned. Pre-hibernation nests were used during late summer and autumn (August–September). Hibernation nests were occupied after active period, between August and May. Some nests had several functions, but the type was categorized according to the main function of the nest.
Construction materials were documented and nest locations were divided into four categories: (1) under a bush or against a tree, (2) cavities on the ground (in a moss tussock or under a tree root), (3) vicinity of a building (under a woodpile, under or on the side of a building) or other, (4) in loose leaves, twigs or grass. The exact nest site location was missing from 16 % of nests (e.g., those in private gardens). The habitat type of each nest site was obtained from CORINE land cover data (2006, © EEA; Fig. 1).
The total number of nests used and changed was calculated only for the hedgehogs tracked throughout the active period. For sex-related comparisons, the active period was divided into three even seasons: the mating season (May 1–June 15), post-mating season (June 16–July 31), and pre-hibernation season (August 1–September 15). This enabled the study of possible changes in nest use and number during these seasons. If the daytime location of a hedgehog in a nest was missing (as in 195 cases out of 2378 nest locations), the hedgehog was expected to use the same nest it had used on the previous day. Sequential use of a nest by multiple individuals was also recorded. Geographic nest locations were included in the home-range analysis (Kernel 50 % and Kernel 95 %; see Rautio et al. 2013) in order to estimate spatial patterns of nest usage and nesting habitat selection. The number of nests in the core area (K50, the area of intensive use), outside K95 and in the part of the home range overlapping with their neighbour’s (K95 and K50) were analysed by season (mating, post-mating, per-hibernation season) and sex (ArcGIS 10; ESRI Inc., Redlands, CA, USA).
Statistical analyses
Fisher’s exact test was used to test the differences between females and males in (1) frequencies of sequential nest use and (2) frequencies of resting without a nest. The differences in proportion of each nest type in three main habitat classes (constructed areas, other constructed areas and forest and semi-natural areas; see Fig. 1) were modelled with generalized estimating equations (multinomial distribution with cumulative logit link function). Differences in (1) the number of nests and (2) the nest change frequency were modelled with linear mixed models, where sex, season and their interaction were included as fixed effects and the individual as a random factor. The proportion of the total number of nests overlapping with K95 areas of other individuals were modelled with generalized estimating equations (events/trials data; binomial distribution with logit link function) where sex, season and their interaction were included as fixed effects and individual as a random factor. Due to the low number of individuals with overlapping core areas (see Rautio et al. 2013) and the low number of nests located in overlapping core areas, the proportion of nests overlapping with K50s of other individuals could not be modelled. Statistical analyses were performed with SPSS 19.0 (SPSS Inc., Illinois, USA) and program R 2.15.0 (Development Core Team 2012). Before analyses, the normality of the variables was checked.
Results
Nest types
Of the 344 observed nests during the study period, 283 were day, 14 breeding, 36 pre-hibernation and 11 hibernation nests. Nest type was typically permanent and only about 2 % of the nests were later reused for a different purpose. Five nests were also used the following active period. For those nests, which building materials were studied (n = 198), materials were gathered from the vicinity of the nests. The majority of nests situated in coniferous forest were built by burrowing into a moss tussock or under a tree root, so data for building materials used in those nests (n = 63) were not included to the following division. At least two different construction materials were used in 59 % of the nests (n = 135). Leaves were used in 73 %, grass in 35 %, twigs in 26 %, forest litter in 20 %, moss in 14 % and rubbish (e.g., plastic bags, wrapping paper) in 6 % of the nests.
A single day nest could be utilized by up to three different hedgehogs, but never simultaneously. Out of the 23 studied individuals 23 % of males and 25 % of females used a nest previously occupied by another individual (frequencies did not differ statistically by sex; Fisher’s exact test, p = 1.000). However, among those hedgehogs that utilized nest previously occupied by another individual, more cases per hedgehog were observed in males than in females (Table 1). The males reused both the nests of other males (61 %) and females (39 %), whereas females almost exclusively reused male nests (80 %). Almost half of the hedgehogs were observed resting without a nest, (the frequencies between sexes did not differ statistically, Fisher’s exact test, p = 0.684). However, among those hedgehogs that were observed resting without a nest, more cases per hedgehog were observed in females than in males (Table 1).
Nest site and nesting habitat
The three most favoured nest sites were as follows: under a bush or against a tree (36 %), vicinity of a building or other (20 %) and cavities on the ground (19 %) (Fig. 1). With very few exceptions, the tracked hedgehogs did not use the nest boxes as resting sites. The exact number of nest boxes offered by local people in the study area was not known, but there were boxes in at least five different private gardens.
We found a significant difference in the proportion of different nest types situated in the three main habitat classes (multinomial regression Wald chi-square=17.0, p < 0.001). Constructed areas (B = −1.8, p < 0.001) and other constructed areas (B = −1.5, p = 0.011) had differing proportions of nest types to those in forest and semi-natural areas. The proportion of day nests was higher in constructed areas than in other constructed and forest and semi-natural areas (Fig. 1). In contrast, the proportion of pre-hibernation and hibernation nests was higher in forest and semi-natural areas than in other areas. Approximately 32 % of all nests in residential areas and 16 % of those in leisure activity areas were situated in forest patches (Fig. 1). Over 40 % of breeding nests were also situated in forest patches.
Number of nests, nest usage and spatial distribution of nests within home range
During the active period, males (n = 3) used on average 24 (±0.8 SE, range 23–26) nests and changed them 30 (±1.5, range 26–33) times, whereas females (n = 4) used 17 (±3.6, range 10–22) nests and changed them 26 (±1.5, range 24–29) times. Although males had more nests on average than females during the active period, there were no statistic differences between sexes in the number of nests used (parameter estimate=−0.43, t = −0.26, p = 0.800) or in the nest change frequency (parameter estimate=1.00, t = 0.30, p = 0.771) when the season was taken into account (Fig. 2). The difference in nest number (parameter estimate=4.29, t = 2.47, p = 0.024) and in nest change frequency (parameter estimate = 10.64, t = 3.02, p = 0.008) between post-mating and pre-hibernation seasons was statistically significant. However, there were no differences in either nest number (parameter estimate=3.14, t = 1.89, p = 0.077) or in nest change frequency (parameter estimate=5.00, t = 1.48, p = 0.158) between mating and pre-hibernation season. The interactions were non-significant between mating and pre-hibernation seasons in nest number (parameter estimate=−2.65, t = −1.16, p = 0.263), and in nest change frequency (parameter estimate=−5.97, t = −1.28, p = 0.217). During the mating season males had 11 nests and they changed those 18 times and females had eight nests and they changed those 13 times. However, the change in nest number (parameter estimate for interaction = −7.71, t = −3.21, p = 0.005) or in change frequency (parameter estimate=−19.79, t = 4.05, p = 0.001) between post-mating and pre-hibernation seasons was unequal between sexes. Number of female nests increased from an average of four post-mating nests to seven pre-hibernation nests, and at the same time the frequency of nest change increased from five to 14. On contrary, in males, the number of nests decreased from an average of 12 to 8, and at the same time the frequency of nest change decreased from 24 to 13 (Fig. 2).

a Number of nests and b nest change frequency in radio tracked hedgehogs (intersection line median, box first and third quartiles, whiskers largest and smallest observations falling within distance of 1.5 times the box size from the nearest quartile, circles outliers, observations with values between 1.5 and 3 box lengths from the upper edge of the box)
In the mating season, 46 % and in the post-mating season over 60 % of females’ nests were located in the core areas of the home ranges (K50) decreasing to 29 % in the pre-hibernation season (Fig. 3). Males had 49 % of nests in core area during the mating season, but during the rest of the active period proportion decreased. In the mating season 12 % and in the post-mating season only 4 % of females nests were located outside of home ranges (K95) increasing to 11 % in the pre-hibernation season. In males, less than 10 % of nests were situated outside K95 during the mating season, but proportion increased during the post-mating season and in the pre-hibernation season over 30 % of nests were outside K95 (Figs. 3 and 4).

Proportion of radio tracked hedgehog nests in a K50 and b outside K95 compared to the total number of nests

Typical home range of male hedgehog and day nests (August 1–31, 2004), pre-hibernation nests (September 1–10, 2004) and hibernation nest (September 11, 2004–April 14, 2005) during one study season
Over 20 % of the female and 12 % of male nests were located within the core area of the home range (K50) while overlapping with the neighbours’ during the active period (Table 2). Both sexes had nests in another individual’s K50 during the mating season, but after that there were no female nests in another female’s core area, also proportion of males nests in another males’ core area decreased. In addition, during the active period, 45 % of the female and 26 % of male nests were located in a part of the home range (K95) that overlapped with their neighbours (there was a statistically significant difference between sexes, Tables 2 and 3). The change in proportion of nests in a part of the home range (K95) that overlapped with their neighbours between mating and pre-hibernation and post-mating and pre-hibernation seasons was unequal between males and females (the both interactions were statistically significant, Tables 2 and 3). In the mating season, approximately half of the females and 27 % of male nests were located in overlapping area, but proportion decreased to 40 % in females and 16 % in males during the post-mating season. During the pre-hibernation season, proportion of nests in overlapping part was only 30 % in females, but 45 % in males.
In August–September, the hedgehogs were still active and used day nests in their home range until they moved to pre-hibernation nests in which they stayed almost around the clock and individual movement activity was very low. Males had on average four (±1.1, range 2–9) pre-hibernation nests that they started to use at the earliest at the beginning of August (Fig. 4). Females had three (±0.5, 2–5) nests that they started to use at the earliest a few weeks later than males (see Appendix). All hedgehogs tracked throughout the hibernation period, except one male, stayed in the same hibernation nest for over 210 days. The males hibernated on average 224 (±4.8) days and females 223 (±2.5) days.
Discussion
The long research period enabled the extensive and versatile observation of hedgehog nesting behaviour at the northern limit of the species range. The pre-hibernation nest, observed for the first time in this study, may be one adaptation for the long northern winter. This nest type is used in preparation for overwintering and is likely to be built to serve as a backup, if the actual hibernation nest were to be destroyed. Our results indicated that forest areas were one of the most important nesting habitats for urban hedgehogs, especially in autumn and winter, when coniferous forests provide a secure hibernation site. To build their hibernation nest, hedgehogs choose locations with surrounding objects that offer good structural support (Morris 1973; Haigh 2011; Haigh et al. 2012; this study). In total, 36 % of all the nests in our study area were situated in a forest, although forest comprised only around 23 % of the study area. Both pre-hibernation and hibernation nests were often built by burrowing into a moss tussock, in contrast to more southern areas where hibernation nests are made of leaves (Morris 1973; Huijser 2000; Haigh 2011). Despite the differences in nest construction material and location, both pre-hibernation nests (present study) and hibernation nests (Morris 1973; present study) have a compact structure with thick and well-supported walls.
Like earlier studies (Morris 1973; Haigh 2011), the present study demonstrated that hedgehogs moved from constructed areas to the forest before overwintering. In addition, we found that hedgehogs had a clear transition period for hibernation during which they built pre-hibernation nests. The transition period for wintering has not been described before and is probably more pronounced in northern areas. The heaviest males started the transition already at the beginning of August. The notable proportion of male nests outside home ranges indicated a decrease in movement and a concentration of activity in wintering areas. After the transition period had begun, the hedgehogs’ behaviour changed. Foraging activity was reduced, new nests were constructed and the hedgehogs remained mostly in their nests. The transition to pre-hibernation nests took place over 2 weeks later in females. The transition period may be related to the spontaneous bouts of transient shallow torpor at the onset of hibernation observed in hedgehogs (Fowler and Racey 1990a). The function of these bouts is unclear, but they may indicate physiological preparation for hibernation or further energy conservation.
Males not only begin to hibernate earlier than females, but they also emerge earlier in order to be ready to find mates as early as possible in the spring (Göransson et al. 1976; Reeve 1994; Rautio et al. 2013; present study). When males left hibernation nests, 2 weeks before females, there was still snow in the forest and the temperature was often below freezing. This indicates that their internal biological clock or day length (Fowler and Racey 1990b) determine when hedgehogs start their spring activity.
In spring and summer, hedgehogs favoured residential areas, including gardens and forest patches that offered a wide variety of nest sites. Urban day nesting sites, in particular, were situated in both constructed and forest and semi-natural areas. More than half of the day nests were situated in constructed areas while one third was found in forests and semi-natural areas. In rural areas, over 80 % of day nests have been observed in hedgerows or forested areas (Huijser 2000; Riber 2006). We found that residential areas were the most favoured in constructed areas and patches of forest were often important resting and breeding habitats. Our data indicated that nest boxes in gardens are of no great significance for adult hedgehogs, although they may serve as a nesting place for juveniles and as occasional resting places for adults (Riber 2006). Day nests were often flimsy, using leaves as a primary building material. The observed nests occasionally fulfilled several functions; some day nests were used later on as pre-hibernation nests. Hedgehogs can return to a previously used nest after a few days or even over a year later (Boitani and Reggiani 1984; Haigh 2011; present study). Few of the day nests studied here fulfilled the same function again in the following active period after more than a year.
The hedgehogs’ relatively large home range in northern latitudes (Rautio et al. 2013) was reflected also in higher numbers of nests in comparison to southern latitudes (Reeve and Morris 1985; Haigh et al. 2012). According to our study, hedgehogs used over 20 nests during their active period. Males with greater home ranges (Rautio et al. 2013) also have a higher number of nests which they change more frequently than females do (Reeve and Morris 1985; Haigh et al. 2012), and this difference between sexes, although not statistically significant, was also recorded in this study. During the mating season, males have the largest home range (Rautio et al. 2013), and they express the widest variety of nesting behaviour, including a high number of nests per individual, usage of the same nests sequentially by different males and resting without a nest. This kind of flexibility may enable males to search for females efficiently (Rautio et al. 2013). In April, males may use nests sequentially because nest construction material is probably scarce to construct a new nest. In our study area, males begin to prepare for the upcoming winter by foraging actively immediately after a short but physically hard mating season (Rautio et al. 2013). Although male home ranges decreased after the mating season, they continued to construct numerous nests that they changed frequently. High nest change frequency may suggest that males change their foraging area often but within a relatively small area during the post-mating season.
During nursing, females’ movements are concentrated in the core area of their home ranges (Rautio et al. 2013) and over 60 % of their nests were thus situated within this area. Females spent several weeks in the same breeding nest with their young. Resting without a nest was also common during the nursing period. This may be due to the increased need for nutrition caused by lactation and the possible resulting daily activity, the desire of females to retire in peace out of reach of their young and the high ambient temperatures in midsummer. Resting without a nest has previously been observed mainly in high ambient temperatures (Huijser 2000) with few exceptions (Reeve and Morris 1985). After weaning, females begin to change day nests more frequently (Boitani and Reggiani 1984; present study), their home ranges increase (Rautio et al. 2013) and nests are situated across a wider area of the home range (present study).
A promiscuous hedgehog mating system (e.g., Reeve 1994; Jackson 2006; Moran et al. 2009) is consistent with the frequent overlap of male and female home ranges (Rautio et al. 2013) and also with the frequency of nests in overlapping parts of the home range. Hedgehogs were observed to utilize nests in overlapping areas most frequently in the mating season. Although home ranges and also nests overlapped widely with those of the same and opposite sex during the mating season, females had no nests in other females’ core areas after that season. This further confirms that females avoid other females during the nursing period (Rautio et al. 2013). In addition, it seems to be rare that a female uses the same nest as another female (Haigh et al. 2012, present study). According to Reeve and Morris (1985), females have not been observed using a nest previously utilized by a male. We did find, however, a few instances of females using a nest formerly used by a male. Nests may play a considerable role in the spread and life cycles of various hedgehog parasites (Brinck and Löfqvist 1973; Philpot and Bowen 1992; Reeve 1994) and sequential use of nests by hedgehogs may ease the spread of parasites from one individual to another (Reeve and Morris 1985).
The studied hedgehogs spent over 85 % of their total time in nests annually, which emphasizes that nesting sites and habitats are essential factors in hedgehog survival. The hedgehog’s hibernation period is longest in northern conditions (Kristiansson 1984; Jensen 2004; this study), and an individual may remain over 8 months in the same hibernation nest. Hedgehogs in this research spent over 223 days in their hibernation nests and did not change nests in winter. In contrast, in temperate areas nests can be changed up to five times during one hibernation period (Morris 1973; Jensen 2004; Haigh et al. 2012).
This study offers new knowledge on hedgehogs nesting behaviour in an urban environment in northern edge of its range. Basic ecological information is needed to ensure effective conservation of this species, as the hedgehog population has been observed to decline significantly in Europe (Hof 2009). We conclude that forested areas and forest patches within the constructed areas are important for hedgehog nesting and particularly as a hibernation habitat. The availability and connectivity of natural and semi-natural habitat is thus important in urban areas (Dowding 2007; Baker and Harris 2007). Increased housing density and more intensive land management are therefore likely to reduce the suitability of the urban environment for hedgehogs and other urban animals. Hedgehog nesting behaviour indicates also latitude adaptation to northern conditions. Large home ranges (Rautio et al. 2013) as well as numerous nests in a home range are needed in harsher climate. A transition period in pre-hibernation nests may allow hedgehogs to conserve energy for the longest hibernation period in their geographical range. In the future, devices such as microGPS-dataloggers (Recio et al. 2013) can enable an even more precise estimation of spatial ecology of the hedgehog.
References
Baker PJ, Harris S (2007) Urban mammals: what does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mamm Rev 37:297–315
Boitani L, Reggiani G (1984) Movements and activity patterns of hedgehog (Erinaceus europaeus) in Mediterranean costal habits. Z Saugetierkd 49:193–206
Brinck P, Löfqvist J (1973) The hedgehog Erinaceus europaeus and its flea Archaeopsylla erinacei. Zoon Suppl 1:97–103
Dowding CV (2007) An investigation of factors relating to the perceived decline of European hedgehogs (Erinaceus europaeus) in Britain. Dissertation, University of Bristol
Fowler PA, Racey PA (1990a) Daily and seasonal cycles of body temperature and aspects of heterothermy in the hedgehog Erinaceus europaeus. J Comp Physiol B 160:299–307
Fowler PA, Racey PA (1990b) Effect of melatonin administration and long day-length on endocrine cycles in the hedgehog Erinaceus europaeus. J Pineal Res 8:193–204
Göransson G, Karlsson J, Lindgren A (1976) Hedgehog and traffic. Fauna och Flora 71:1–6 [In Swedish with English summary]
Haigh A (2011) The Ecology of the European hedgehog (Erinaceus europaeus) in rural Ireland. Dissertation, University College Cork
Haigh A, O’Riordan RM, Butler F (2012) Nesting behaviour and seasonal body mass changes in a rural Irish population of the Western hedgehog (Erinaceus europaeus). Acta Theriol 57:321–331
Hanski IK, Stevens PC, Ihalempiä P, Selonen V (2000) Home-range size, movements, and nest-site use in the Siberian flying squirrel, Pteromys volans. J Mammal 81:798–809
Hof A (2009) A study of the current status of the hedgehog (Erinaceus europaeus) and its decline in Great Britain since 1960. Dissertation, University of London
Huijser M (2000) Life on the edge. Hedgehog traffic victims and mitigation strategies in an anthropogenic landscape. Dissertation, University of Wageningen
Jackson DB (2006) The breeding biology of introduced hedgehogs (Erinaceus europaeus) on a Scottish Island: lessons for population control and bird conservation. J Zool 268:303–314
Jensen AB (2004) Overwintering of European hedgehogs Erinaceus europaeus in a Danish rural area. Acta Theoriol 49:145–155
Kristiansson H (1984) Ecology of a hedgehog Erinaceus europaeus population in southern Sweden. Dissertation, University of Lund
Moran S, Turner PD, Reilly CO (2009) Multiple paternity in the European hedgehog. J Zool 278:349–353
Morris P (1973) Winter nests of the hedgehog (Erinaceus europaeus L.). Oecologia 11:299–313
Parkes J (1975) Some aspects of the biology of the hedgehog (Erinaceus europaeus L.) in the Manawatu, New Zealand. New Zeal J Zool 2:463–472
Philpot C, Bowen R (1992) Hazards from hedgehogs: two case reports with a survey of the epidemiology of hedgehog ringworm. Clin Exp Dermatol 17:156–158
Rautio A, Valtonen A, Kunnasranta M (2013) The effects of sex and season on home range in European hedgehogs at the northern edge of species range. Ann Zool Fenn 50:107–123
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Recio MR, Mathieu R, Latham MC, Latham ADM, Seddon PJ (2013) Quantifying fine-scale resource selection by introduced European hedgehogs (Erinaceus europaeus) in ecologically sensitive areas. Biol Invasions. doi:10.1007/s10530-013-0410-6
Redman P, Selman C, Speakman JR (1999) Male short-tailed field voles (Microtus agrestis) build better insulated nests than females. J Comp Physiol B 169:581–587
Reeve N (1994) Hedgehogs. Academic Press, London
Reeve N, Morris P (1985) Construction and use of summer nests by the hedgehog (Erinaceus europaeus). Mammalia 49:187–194
Riber AB (2006) Habitat use and behaviour of European hedgehog Erinaceus europaeus in a Danish rural area. Acta Theriol 51:363–371
Vogt FD, Lynch GR (1982) Influence of ambient temperature, nest availability, huddling, and daily torpor on energy expenditure in the white-footed mouse Peromyscus leucopus. Physiol Zool 55:56–63
Acknowledgments
Thanks are due to M. Haatanen, A. Kirjavainen and P. Rännänen for their invaluable help. The authors are also very grateful to K. Sotejeff-Wilson and L. Murto for linguistic revision, M. Valtonen for commenting on the manuscript, and J.-P. Rautio for help with the figures. Constructive criticism by two anonymous reviewers helped to improve the manuscript. This study was supported by the Maj and Tor Nessling Foundation, the Jenny and Antti Wihuri Foundation, the E. J. Sariola Foundation, the Ella and Georg Ehrnrooth Foundation, the Finnish Cultural Foundation; the North Karelia Regional Fund, the Oskar Öflunds Foundation and the Finnish Doctoral Programme in Environmental Science and Technology (EnSTe).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Andrzej Zalewski
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 13 kb)
Rights and permissions
About this article
Cite this article
Rautio, A., Valtonen, A., Auttila, M. et al. Nesting patterns of European hedgehogs (Erinaceus europaeus) under northern conditions. Acta Theriol 59, 173–181 (2014). https://doi.org/10.1007/s13364-013-0150-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-013-0150-0