
Abstract
There was previously no information on nesting, seasonal body mass or the hibernation behaviour of the hedgehog in Ireland. Between 2008 and 2009, hedgehogs were caught, weighed weekly and monitored by radio tracking at a rural Irish site. Day nests were recorded in the active period and hibernacula thereafter. Arable land has been reported to be poorly utilised by hedgehogs in the UK and Denmark. In Ireland, day nests and hibernacula were constructed in the hedgerows of arable land indicating that these areas will be exploited if hedgerow is maintained and highlights the importance of maintaining hedgerows in arable areas. Individual females returned to the same day nest significantly more than males. Over the 2 years, individuals were found to occupy a mean of 1.8 (±0.9; SD) hibernacula (maximum of 3) and they rotated between nests up to four times (mean of 2.5 ± 1.6). When hedgehogs occupied multiple hibernacula, those occupied in mid winter (December and January) were occupied for significantly longer than those occupied at the start (October and November) and end (February and March) of hibernation. Studies in the UK and Denmark have reported on hedgehog hibernation. However, the winter climate in southern Ireland is milder than other areas of Europe and thus it was expected that hibernation characteristics would also differ, i.e. shorter hibernation periods, earlier emergence, lower body mass loss and the ability to survive hibernation at a lower body mass. This proved to be the case with a mean hibernation period (±SE) of 148.9 (±0.5) days, a mean body mass loss of 17.0 (±0.53) %, emergence in March and the ability of late juveniles to survive at a pre-hibernation body mass of 475 g.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A number of variables are believed to limit the distribution of the western hedgehog in an area, with nest sites (Jensen 2004), food availability (Kristiansson 1984) and the presence of predators (Micol et al. 1994) deemed to be the most important of these. For a hibernating species, like the western hedgehog, adequate nest sites are essential if a habitat is to meet the animal’s basic requirements. In Denmark, Jensen (2004) recorded that 55 % of hibernacula occurred in forested areas, with a similar result found by Riber (2006) for day nests in Denmark. In Ireland, only 9.6 % of the land area is currently covered by forest (Rudel et al. 2005). However, while Ireland has the lowest percentage of forest cover by land area of all European countries (EPA 2008), Ireland has extensive hedgerow networks, which have been recognised as important habitats for conservation on a European level (Pithon et al. 2005). Most of this hedgerow has remained intact since it was first planted in the 18th century; whereas in most other European countries, it has become a special feature, restricted to small areas (Cabot 1999). Agricultural intensification, and the associated loss of habitat, has resulted in hedgerows becoming regarded as important contributors to biodiversity conservation in the agricultural landscape (Gelling et al. 2007). Unfortunately, as farms have become more specialised and efficient, and with the growing need for larger fields, this has had a knock-on effect on hedgerows, with a reduction of 50 % of the hedgerow stock in the UK (Robinson and Sutherland 2002). However, while farmland is the most intensively managed part of the Irish landscape, it is still generally less intensive than in other European countries (Cabot 1999) and small field sizes have been maintained in many areas (Pithon et al. 2005).
Unlike other mammal species who build a secure permanent nest site, the day nests of hedgehogs are often flimsy structures designed for short term occupancy (Morris 1973). Despite this, in the UK hedgehogs have been shown to exhibit philopatry, returning to a particular nest throughout the year but usually occupying multiple nests (Morris 1969). In the UK, this was generally dependent on the size of the individual’s home range and Reeve (1981) noted a tendency for males to move nests more often than females. Individuals may use the same nest but simultaneous occupancy has not been recorded (Riber 2006).
In Denmark, Jensen (2004) reported that the average duration of hibernation was 178.8 ± 13.1 (SE) days for juveniles and 197.7 ± 2.2 days for adults. The only record of hibernation in Ireland was recorded by a Mrs. Kenny in Tipperary who noted the hibernation period of an individual hedgehog in her garden over 4 years as between 139 days in 1980 and 1982 and 150 in 1983 (Fairley 1984). In Britain, adult hedgehogs usually weigh between 600 and 700 g increasing to about 900–1,000 g in autumn (Reeve 1982). In New Zealand, Parkes (1975) reported a mean body mass for males of 706 ± 10 g and for females 688 ± 9.9 g. In previous studies, males, in contrast to females, showed a fairly constant body mass throughout the year, but with a tendency to increase body mass just prior to hibernation, having reduced their body mass during the breeding season (Reeve 1981).
Juveniles are capable of putting on body mass quickly and Reeve (1981) stated that new juveniles that appeared in early August in the UK increased their body mass to exceed 500 g by October. According to Morris (1969), some individuals can achieve 800–900 g within a year of their birth and Jackson (2006) observed that subadults were on average nearly a third heavier 1 year later.
The necessary body mass required to survive hibernation varies depending on the country’s climate. In the UK, hedgehogs have been reported to hibernate from October or November through to April (Morris 1969; Jackson 2001). According to Reeve (1982), British hedgehogs are smaller than continental hedgehogs and are able to survive winter with a lower body mass. Estimates in the UK for the minimum hibernation body mass range from 450 (Morris 1984) to 650 g (Bunnell 2002). In Denmark, they needed to reach 513 g to survive (Jensen 2004). Brockie (1990) estimated that hedgehogs in New Zealand needed to weigh at least 300 g to survive hibernation. In Sweden, Kristiansson (1984) found that hedgehogs tended to have a larger pre-hibernation body mass than British hedgehogs, something which he suggested could be due to the harsher Swedish winters. To date, the body mass required for an Irish hedgehog to survive hibernation is unknown. Grass continues growing for longer in Ireland than any other European country (Cabot 1999), and the winters are generally milder in Ireland, which may allow for a shorter hibernation period.
With this milder climate, the current study hypothesised that variations would be apparent in the hibernation activity of hedgehogs in Ireland, in comparison to elsewhere. In particular, the study aimed to test the hypotheses that hedgehogs could survive hibernation at a lower weight, hibernation is shorter, weight loss more conservative and emergence earlier than that previously observed elsewhere in Europe. With Ireland’s less intensive agricultural practices and greater density of hedgerows, the current study also aimed to investigate the location of day nests and hibernacula and the contribution of this, to habitat selection.
Materials and methods
Study area
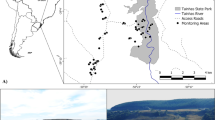
The study was carried out between June 2008 and November 2009 on a 97-ha site (51° 53′ 59.5″ N latitude, 8° 29′ 03.7″ W longitude), 36.8 km from Cork City and 5.3 km from the nearest town of Bandon. Land use was 23 % arable, 64 % pasture, 7 % residential garden, 1 % scrub, 1 % marsh and 4 % wood (Fig. 1). Residences (8 %) were clustered throughout the study site, the majority of which was associated with farm yards.

Study area: the 43 ha searched for hedgehogs in 2008 (inner square) and 97 ha searched in 2009 (outer square) near Bandon, Southern Ireland. The squares in red show the location of day nests; those in green, hibernacula
Daily minimum and maximum temperatures were measured at midday. Rainfall data were obtained from Met Eireann (www.met.ie; Fig. 2).

Minimum and maximum monthly temperature (°C) and rainfall (mm) over the study period
Capture and marking
Hedgehogs were captured by hand with the aid of spotlights. Individuals were marked using a unique colour combination of heat shrink plastic tubes (R.S. Components Ltd, Northants, UK) which were attached to the spines. Fifteen were applied to three specific regions (left of head, centre and right of head) on each animal with glue (Evo-Stik). Reflective tape (CH Marine, Cork) was also attached to one of the middle markers so that the head region could be identified while tracking. The tubes acted as a visual aid and minimised the need to recapture the animal each time for individual identification. Individuals were also marked using passive integrated transponder (P.I.T.) tags (MID Fingerprint, Dorset, UK) inserted into the upper hind leg (Doncaster et al. 2001; Jackson et al. 2004). This allowed individuals to be identified after hibernation.
All hedgehogs caught were classified as adult or juvenile, sexed, weighed using digital scales (Harvard apparatus, Kent, UK), hind foot measurements taken and growing and broken spines noted. Tagged individuals were weighed at weekly intervals. The animal was considered to be a juvenile if it satisfied all of the following criteria: body mass less than 600 g when first caught, hind foot length of less than 3.6 cm and presence of growing spines throughout the pelage. All procedures were carried out in accordance with current regulations. Licenses were obtained from the Department of Environment, Heritage and Local Government.
Direct following
Spotlighting for hedgehogs took place from 26th June 2008 until 28th September 2008. Eight hedgehogs were caught during spotlighting nights and these were fitted with a tip light (MK IV, Murrays and Co. Ltd, Patrick St, Cork, Ireland) placed on the back and were monitored for a total of 95 h over 17 nights by direct following (Table 1). Location fixes were taken every 10 min using a Garmin GPS 60 (CH Marine, Cork, Ireland). Animals were followed until they returned to the nest site, or until they could no longer be located. It was also noted whether or not they returned to the nearest nest at the end of a night’s foraging.
Radio tracking
All animals caught after 28 September 2008 were fitted with radio tags. Eight hedgehogs were fitted with 173 MHz, R1-2B transmitters (Holohil Systems Ltd, Ontario, Canada) and attached to the animal after the manner of Jackson and Green (2000). The entire tag weighed 10 g. These eight hedgehogs were radio tracked before they entered hibernation for 160 h over 33 nights (Table 1). Day nests were either identified at the end of the night or during the following day.
Six of these hedgehogs were monitored throughout hibernation and upon emergence. Individuals were considered to be hibernating if they did not leave their nest for seven consecutive nights. Hibernacula were identified in accordance with Morris’ (1973) description of winter nests. Once the animals were considered to have entered hibernation, their location was checked at weekly intervals.
Data analysis
GPS positions of nests and hibernacula were plotted onto ortho-photographs (Ordinance Survey of Ireland) of the area using the Geographic Information System (GIS) software Arc map version 9.2. When means are provided they are followed by the ± standard error, unless otherwise stated. Tests for normality were performed on Brodgar software for univariate and multivariate analysis and multivariate time series analysis version 2.6.3. PASW Statistics Version 17 was used for all further statistical analysis.
Results
Location of day nests
Day nests of radio tagged hedgehogs were located 260 times between June 2008 and November 2009, during which 117 different nests were identified (Fig. 1). Day nests were recorded 122 times in 2008 and 138 times in 2009. A total of 117 individual day nests were utilised by hedgehogs, 53 % of these were located in pasture and 30 % in arable land (Table 2). However, while the majority of day nests was in pasture, hedgehogs returned significantly more often to nests in arable land (Kruskal–Wallis, H = 8.536, df = 2, p < 0.05, followed by Dunn’s comparison test; Table 2). In general, day nests were situated amongst thick bramble. However, in warm weather (13–14.5 °C) hedgehogs were occasionally observed (n = 3) to nest amongst long grass in pasture (Table 2).
Location of hibernacula
Six hedgehogs were tracked throughout hibernation in 2008/2009 and five in 2009/2010. Seventeen hibernacula were identified at this time (Fig. 1), with the majority located in hedgerow of arable land (35 %) and in scrub (29 %; Table 2). Few hibernacula were located in pasture hedgerow (6 %) despite 37 % of day nests being located in this microhabitat (Table 2).
Use of multiple day nests
Hedgehogs had several day nests that were used only once (Fig. 3). Some nests were used more often, up to seven times by some individuals (Fig. 3). Females returned to the same nest significantly more often than males. (One-way ANOVA, following Log(x) transformation, F = 5.411, df = 1, p < 0.05; 2009 data only). This pattern was observed in two successive years. In 2009, females (n = 4) moved a mean of 2.27 ± 0.03 times vs. males (n = 7) moved a mean of 1.34 ± 0.02 times. In 2008, females (n = 3) returned to a nest a mean of 5.2 ± 0.18 times whereas the corresponding figure for males (n = 3) was 3.8 ± 0.15 occasions. In addition, nests were utilised by a number of different hedgehogs but never simultaneously.

The number of nights each individual hedgehog remained in an individual hibernaculum from the first day in which they entered hibernation (individuals are only included if they were monitored for the entire hibernation period)
Movement between hibernacula
Of the six hedgehogs monitored throughout hibernation in 2008/2009, only two remained in the same hibernaculum for the entire hibernation period (Fig. 3). In 2008/2009 and again in 2009/2010, adult male (#75EC) was the only animal to occupy a single hibernaculum (Fig. 3). One adult male (#ACOO) moved four times during hibernation between three different hibernacula.
In 2008/2009, two male hedgehogs (#A3D6 and #ACOO) swapped hibernacula on a number of occasions (n = 3) throughout hibernation. This exchange of hibernacula was observed again in 2009/2010 when an adult female (#FA56) and juvenile male (#E89D) moved between the same two hibernacula four times, entering a hibernaculum as soon as the other had exited it.
When a hedgehog occupied multiple hibernacula, the first was utilised for a significantly shorter time (19.8 ± 0.6 nights) than the second hibernaculum (81.4 ± 1.2 nights; Mann Whitney, U = 1.5, n 1 = 8, n 2 = 5, p < 0.05). Hibernacula occupied in mid winter (December–January) were also occupied for significantly longer than those hibernacula occupied at the end (January–March; 21.8 ± 0.3 nights) of hibernation (Mann Whitney, U = 1.0, p < 0.05; Fig. 3).
Emergence time
In the 2008/2009 season, four of the six hedgehogs emerged within a night of one another (30th–31st March 2009). One emerged 2 weeks later (16th April 2009). Another hibernated for 174 nights, emerging on the 12th May (Table 3). This was the longest hibernation recorded during the study. In 2009/2010, the first hedgehog emerged on the 18th March 2010.
Adult body mass during the active period
Over the active period (2008–2009), adult males (n = 7) were significantly heavier than adult females (n = 4; Z = 2.679, n 1 = 55, n 2 = 27, p < 0.01; Fig. 4). The mean body mass (±SE) of adult females was 998.9 g (±0.37), ranging from a mean of individual mean of 877.9 g (±1.98) in August to 1,116 g (±1.21) in October. The equivalent body mass of adult males was 1,065 g (±0.19; SE), ranging from 1,026 g (±0.90) in July to 1,192 g (±1.38) in October.

Mean (±SD) monthly weights of adult males and females
The active period was then grouped into four bimonthly time periods (March and April, May and June, July and August and September and October), and both sexes were significantly heavier in the period approaching hibernation than they were in July/August or the first 2 months after emergence (March and April; ♀ one way ANOVA followed by Tukey’s post hoc test, F = 4.548, df = 3, p < 0.05; ♂ one way ANOVA followed by Tukey’s post hoc test, F = 2.896, df = 3, p < 0.05).
Surprisingly, individuals tagged in successive years were found to be of a similar monthly body mass in both years (Table 3). Generally individuals were only marginally heavier or lighter (<100 g) in the same month of the following year. Notably, the greatest body mass difference (>100 g) was August 2008 and August 2009 evident in two males (75EC and 8C88; Table 3).
Juvenile body mass during the active period
Juveniles showed a mean body mass increase of 7.78 g (±0.24; SE) per day from first capture, combining years and sexes. Juveniles (n = 6) almost doubled their body mass from first capture to hibernation (mean of 180.13 % (±1.29)). The maximum body mass achieved by a juvenile prior to hibernation was 1,024 g (Table 4). This was equivalent to the weight of adults at this time.
Reduction in body mass post hibernation
Males and females were of a similar weight immediately prior to hibernation (F = 0.033, p > 0.05; Fig. 4).
In 2008/2009, the mean body mass of adult hedgehogs on entering hibernation was 1,181.3 g (±0.9; SE). After hibernation, females were found to have lost significantly more body mass (mean 301 g (±3.91; SE)) than males (mean 108 g (±2.55); t = 5.100, df = 4, p < 0.01). Females lost between 15 % and 38 % of their body mass. In comparison males lost just 3–6 % of their body mass (Table 3).
Due to tag failure over the winter, only one hedgehog was weighed immediately upon emergence in 2010. This was the juvenile male (#E89D) and upon emergence he had lost 29 % of his body mass over the hibernation period (Table 3).
Discussion
In the present study, 53 % of day nests were located in the hedgerows of the pasture and 30 % in the hedgerows of arable land. The majority of these were located in patches of brambles. In the UK, Reeve (1981) found that of 58 day nests recorded, 31 (53.5 %) were in prickly vegetation. Of those, the majority 24 (77 %) were in brambles. Morris (1969) also found that hedgehogs showed a marked preference for building nests in brambles and that these nests lasted longer (Morris 1973). Although in the present study, the use of bramble hedgerow was similar, the location was different, since in other studies arable land has been shown to be rarely used by hedgehogs (Dowie 1987; Doncaster 1994; Doncaster et al. 2001; Riber 2006; Hof 2009). For example, in the Netherlands Huijser (2000) found over 60 % of day nests in hedgerow under bramble but arable land was the least used habitat. In the UK, hedgerows survive least well in districts where arable farming prevails, as the function of stock containment has become redundant (Croxton et al. 2004). The regular and intensive post-harvest flailing undertaken in the interest of tidiness and shade reduction has resulted in some hedges becoming very reduced, and sometimes shorter than the crops they surround (Croxton et al. 2004). Hedgerows play a critical role in maintaining agricultural biodiversity by providing habitats, refuges and corridors for large numbers of species (Burel 1996), such as birds (Hinsley and Bellamy 2000) and invertebrates (Burel et al. 2004). Hof (2009) suggested that hedgerows may enhance the suitability of fields for hedgehogs, especially in arable-dominated areas. According to Pollard et al. (1977) hedgerows with good ground cover are easily the best kind of hedgerow to support biodiversity, not only providing suitable nest sites for hedgehogs, but also the leaf litter provides suitable shelter for potential prey such as molluscs and coleoptera. However, while the arable field in the present study had a well established hedgerow with a thick bramble understory, in many cases this is considered unsightly (Croxton et al. 2004) and is subsequently removed.
Hibernacula were most commonly built in the hedgerow of arable fields and scrub. While day nests were regularly built in the arable field, scrub had been used rarely in the months prior to hibernation. This corresponded to a move into areas of scrub in October and November to build hibernacula (Haigh 2011). While the scrub may have offered little in terms of available prey (Haigh 2011), the thick bramble patches in this habitat provided adequate support in which to build secure hibernacula. Morris (1973) found that hedgehogs left the exposed parts of the park that he was studying for a sheltered hibernation site, choosing locations that offered good structural support from surrounding objects. In the present study, pasture was used regularly for day nests but only two hibernacula were located in this habitat (and both of these hibernacula were situated within walls). Morris (1973) found that grass nests were particularly prone to desertion. Therefore, while hedgerow in the pasture may be sufficient for the often flimsy day nests, they may not have offer sufficient protection for use as hibernacula.
In the present study hedgehogs utilised up to 11 day nests between which they rotated. In previous studies, males have been found to utilise a greater number of nests than females (Reeve 1981; Boitani and Reggiani 1984), and this, while not significant, was also recorded in the current study. In the present study, during the active period, males occupied a much larger home range than females, occupying a mean area of 56 ha (±0.7) while females had an average home range of 16.5 ha (±0.5; Haigh 2011). The fact that male hedgehogs had a much larger home range may have made it more difficult for them to return to a particular day nest and therefore necessitated the use of a greater number of day nests. This is also supported by the fact that females returned to an individual nest significantly more often than males. However, hedgehogs did not always return to the day nest nearest to where they were foraging, but instead travelled back to specific nests, demonstrating a degree of philopatry. One adult female, in particular, was observed to travel back from the middle of the arable field to a nest located at the bottom of the garden, despite passing 2-day nests that she used regularly. The nest to which she returned was located in an abandoned silage bale and may have offered better protection on colder nights, as on one occasion the nightly temperature had decreased from 8 °C to 6 °C and subsequently to 4 °C. This individual began hibernating in this nest that was also utilised at different times by another adult female and an adult male. The sequential use of a nest by multiple individuals has been previously reported by Reeve (1981) in the UK and Riber (2006) in Denmark. Similarly in the present study, adults were never observed simultaneously using the same nest.
Only one adult male and one adult female remained in the same hibernacula for the whole period of hibernation in 2008. This adult male was also the only animal to remain in the same hibernaculum in 2009. In both years the remaining hedgehogs moved up to four times occupying a maximum of three hibernacula. Morris (1973) reported that 60 % of hibernacula was used for less than 2 months and the longest period of occupation was 6 months (Morris 1969). In the present study, the longest occupation recorded was similar, i.e. 6 months and 9 days. When a hedgehog moved during hibernation, the hibernaculum that the hedgehog utilised half way through the hibernation period was occupied for a significantly longer time than others. Jensen (2004) also found that there was a strong tendency for hedgehogs that were using multiple nests to use the one that they had entered during mid winter for the longest time. Hibernating hedgehogs are subject to regular arousals and it has been found that hedgehogs have a less profound winter sleep in the beginning of the hibernation period (Walhovd 1978). It is therefore interesting that in the present study hedgehogs occupied hibernacula entered in mid winter for longer than those entered at the start or end of hibernation. Morris (1973) noticed a tendency for hedgehogs to move hibernacula following periods of sunshine and this is more likely to occur in early spring than in mid winter. In the present study, the temperature was at its lowest in December and January in both years and only began to rise in February.
In 2008, the hedgehogs began to enter hibernation on 19th October. On 29th October, the temperature dropped from 9 °C to −2 °C, there was ground frost and no recorded invertebrate activity (Haigh 2011). Slug activity is markedly reduced at temperatures below 10 °C (Young et al. 1993). All of the hedgehogs were inactive at this time. At the end of the week as the temperature began to rise (4 °C), some of the lighter hedgehogs (n = 4) re-emerged. In New Zealand, Webb and Ellison (1998) also found that under natural winter conditions of temperature and photoperiod wild hedgehogs readily became torpid when access to food was restricted for 48 h. Therefore the lack of activity during these few days in the present study was probably driven by the lack of prey, brought on by the sudden drop in temperature.
At the beginning of this study, it was hypothesised that hibernation would be shorter and emergence would be earlier, due to Ireland’s milder climate, and the data supported this. The mean (±SE) length of hibernation recorded for the study population was 148.9 (±0.5) nights, with four of the six hedgehogs emerging on the 30th–31st March 2009 and the following year from 18th March 2010. The majority of reports in the UK and Denmark record the average duration of hibernation to be 178.8 ± 13.1 days for juveniles and 197.7 ± 2.2 days for adults (Jensen 2004; Morris 1969; Jackson 2006) with hedgehogs leaving their hibernacula during the last 2 weeks of April, with the remaining hedgehogs resuming activity in the first 2 weeks of May (Jensen 2004; Morris 1969; Jackson 2006). Similarly, Walhovd (1979) found that in Denmark the hedgehogs stayed in hibernation for 6 months and never resumed activity before May.
While hedgehogs at the site entered hibernation at different times (19th October–13th November 2008), four of the six hedgehogs resumed activity on the 30th–31st March 2009, but two adult females did not become active until 16th April and the 12th May 2009. In 2009/2010, the first hedgehog resumed activity on the 18 March 2010. This was also the first hedgehog that had entered hibernation the previous October. In 2009/2010, Ireland experienced its coldest winter since 1963; with temperatures around 2 °C lower than average (Met Eireann). An adult female (#FA56) and juvenile male (#0428) were never seen again after the winter of 2009/2010 and therefore they may not have survived. However, despite this cold winter, 2010 had the sunniest February in the south for over 40 years (Met Eireann). As stated earlier, Morris (1973) noted a tendency for hedgehogs to emerge following periods of sunshine and the earlier arousal in 2010 may therefore have been a result of high incidence of sunshine in February 2010.
At mean body masses of between 878 and 1,116 g for adult females and 1,026–1,192 g for adult males, the group in the present study was heavier than in other studies. In the UK, Reeve (1981) reported that body mass increased from 600 to 700 g in spring to about 900–1,000 g in autumn and found that it was relatively uncommon for hedgehogs to weigh more than 1,000 g. Similarly Dowding et al. (2010) recorded average body masses of 846 g ± 119 for males and 792 g ± 157 for females also in the UK. In New Zealand, Parkes (1975) recorded a mean body mass for males of 706 g ± 10.0 and 688 g ± 9.9 for females. The heavier body mass of hedgehogs in the present study may have been due to optimum feeding conditions at the site, a supposition supported by the high density of individuals occurring there (Haigh 2011), and the juveniles ability to increase body mass over short time periods. However, this larger body mass than hedgehogs in other areas of its range was also recorded amongst Irish road kill (Haigh 2011; mean 918 g ± 0.28 n = 42), a maximum body mass of 1,254 g was recorded and twelve individuals weighed above 1 kg. Variation has also been found amongst mustelid size in Ireland and England (Erlinge 1987; Dayan and Simberloff 1994). This has thought to be related to variations in prey size (Erlinge 1987) and a reduction in selective pressure in the absence of some potential competitors in Ireland (Dayan and Simberloff 1994). White and Searle (2007) found that body size of common shrews on islands was positively related to distance from mainland, suggesting a role for founder events, in determining body size of common shrews on islands. In the current study the lack of genetic variation amongst Irish hedgehogs suggested a small founder number (Haigh 2011) and this combined with the lower number of competitors in Ireland, may result in Irish hedgehogs being larger.
Both sexes showed fluctuations in body mass throughout the year, with females being at their minimum body mass in August. Morris (1969) suggested that during August the mean body mass of female’s decreases since few are pregnant at this time. Males on the other hand were at their lowest body mass in July in the present study. This was the peak of courtship activity and is probably related to a reduction in foraging at this time, due to a preoccupation with mating (Haigh 2011). Individuals that were weighed over the 2 years showed little fluctuation from 1 year to the next. However, two of the males (#75EC and #8C88) were at a lower body mass (100 and 151 g, respectively) in August 2008 than in August 2009. In August 2008, both males were still engaged in courtship behaviour, but it had ceased by this time in 2009 (Haigh 2011). In Sweden, during the mating period, Kristiansson (1984) also observed that the body mass of the adult males decreased by about 10 % from May (955 g) to June (870 g) but increased considerably from June to September (870–1,410 g). In the present study, both sexes were at their maximum body mass just prior to hibernation, with males beginning to increase in body mass in August and females in September. Morris (1969) and Reeve (1981) both reported a general increase in body mass during the active season similar to the present study, with both sexes reaching a peak in their body mass pre-hibernation.
Jackson (2006) found that the body mass of adults weighed 1 year later was on average unchanged. In the present study once individuals had reached adulthood, monthly body masses were similar 1 year later and even 2 years on. Therefore despite body mass fluctuations due to pregnancy, courtship behaviour or loss through hibernation, adult body masses remain stable on an annual basis. This indicates that during the study period, food resources must also have remained relatively constant in this area.
Once juveniles reached independence they showed a dramatic increase in body mass, demonstrating that even late litters in Southern Ireland are capable of reaching the required body mass to survive hibernation. Bunnell (2009) reported that in the UK late juveniles had a significantly higher growth rate than earlier litters, further emphasising the ability of late litters to survive. Kristiansson (1984) recorded that juvenile’s body mass increased linearly from about 280 g in August to about 600 g in October, i.e. an increase of about 210 % in 2 months. In the present study juveniles showed a mean increase of 180.1 % (±1.29) from their body mass when first weighed to hibernation, with one individual increasing by 289.3 % in less than 2 months. As was the case with the adult population, the juveniles in this Irish population were heavier than reports in other studies. In the UK, Morris (1969) reported that some animals can achieve 800–900 g within a year of their birth. However, in the present study, one individual had reached 865 g less than 2 months from independence, with another reaching a pre-hibernation body mass of 1,024 g.
The mean (±SE) weight loss during hibernation was 17.0 ± 0.53 %, which was as expected low in comparison to other studies, a fact that may be due to milder winters or the availability of more secure nest sites in Ireland. Kristiansson (1984) in Sweden recorded body mass losses of 20–40 % and Morris (1969) in the UK observed a reduction of 18 % in males and 39 % in females. However, Jensen (2004) in Denmark observed that although the majority lost 21.3–37.4 % of their body mass, one individual lost only 4.5 %. In the present study, females lost significantly more body mass than males during hibernation. This has also been recorded in other studies (Morris 1969) and may be due to them not building up as much fat reserves, due to late litters. In the present study one litter did not reach independence until October 17/18th 2008 (Haigh 2011), and late litters were also recorded amongst road kill (Haigh 2011). Walhovd (1979) found that hedgehogs, that refused to eat during the hibernation period, lost twice as much in body mass as hedgehogs that occasionally fed, but he reported that hedgehogs weighing above 1,000 g, as in the present study, rarely ate. However, while two of the males in the present study moved between hibernacula the most, the male who lost the least amount of body mass did not move for the duration of hibernation. According to Tähti and Soivio (1977), one arousal lasting only 3–4 h consumes the energy equivalent of several days in hibernation. In the present study, an adult female remained in a single nest for over 6 months and lost only 23 % of her body mass, while the female who lost 38 % of body mass moved twice. This contrasts to Walhovd’s (1979) study on captive hedgehogs, where he reported body mass losses of between 22 % and 78 %, with the greatest losses reported in two males who both remained in the same nest and did not eat for the entire 6 months of hibernation.
At the beginning of this study, it was hypothesised that the temperate climate and corresponding milder winters in Ireland might result in certain differences in the hibernation behaviour of hedgehogs in Ireland in comparison to elsewhere. The data supported this, with hedgehogs emerging earlier than reports elsewhere and showing a shorter duration of hibernation. A smaller body mass loss in this study in comparison to other countries is not surprising in light of the correspondingly shorter duration of hibernation and the availability of secure hibernacula sites. The location of both day nests and hibernacula in the hedgerow of arable land has shown that these areas will be utilised if hedgerow is maintained. It therefore highlights the importance of maintaining hedgerows in arable areas, particularly with bramble understory which in too many cases is considered unsightly and of little value.
References
Boitani L, Reggiani G (1984) Movements and activity patterns of hedgehogs (Erinaceus europaeus) in Mediterranean coastal habitats. Zeitschrift fuer Saeugetierkunde 49:193–206
Brockie R (ed) (1990) European hedgehog. The handbook of New Zealand mammals. Oxford University Press
Bunnell T (2002) The assessment of British hedgehog (Erinaceus europaeus) casualties on arrival and determination of optimum release weights using a new index. J Wildl Rehabil 25:11–22
Bunnell T (2009) Growth rate in early and late litters of the European hedgehog (Erinaceus europaeus). Lutra 52:15–22
Burel F (1996) Hedgerows and their role in agricultural landscapes. Crit Rev Plant Sci 15:169–190
Burel F, Butet A, Delettre YR, Millàn de la Peña N (2004) Differential response of selected taxa to landscape context and agricultural intensification. Landsc Urban Plan 67:195–204
Cabot D (1999) Ireland. Harper Collins, London
Croxton PJ, Franssen W, Myhill DG, Sparks TH (2004) The restoration of neglected hedges: a comparison of management treatments. Biol Conserv 117:19–23
Dayan T, Simberloff D (1994) Character displacement, sexual dimorphism, and morphological variation among British and Irish mustelids. Ecology 75:1063–1073
Doncaster C (1994) Factors regulating local variations in abundance: field tests on hedgehogs, Erinaceus europaeus. Oikos 69:182–192
Doncaster C, Rondinini C, Johnson P (2001) Field test for environmental correlates of dispersal in hedgehogs Erinaceus europaeus. J Anim Ecol 70:33–46
Dowding CV, Harris S, Poulton S, Baker PJ (2010) Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward. Anim Behav 80:13–21
Dowie M (1987) Rural hedgehogs many questions to answer. Game Conservancy Annu Rev 18:126–129
EPA (2008) State of the environment report. Environmental Protection Agency, Johnstown Castle
Erlinge S (1987) Why do European stoats Mustela erminea not follow Bergmann’s rule? Ecography 10:33–39
Fairley J (ed) (1984) An Irish beast book, 2nd edn. Blackstaff, Belfast
Gelling M, Macdonald D, Mathews F (2007) Are hedgerows the route to increased farmland small mammal density? Use of hedgerows in British pastoral habitats. Landsc Ecol 22:1019–1032
Haigh A (2011) The ecology of the european hedgehog (Erinaceus europaeus) in rural Ireland. PhD thesis, University College Cork
Hinsley SA, Bellamy PE (2000) The influence of hedge structure, management and landscape context on the value of hedgerows to birds: a review. J Environ Manag 60:33–49
Hof A (2009) A study of the current status of the hedgehog (Erinaceus europaeus), and its decline in Great Britain since 1960. PhD thesis, Royal Holloway, London
Huijser M (2000) Life on the edge. Hedgehog traffic victims and mitigation strategies in an anthropogenic landscape. PhD thesis, Wageningen University
Jackson DB (2001) Experimental removal of introduced hedgehogs improves wader nest success in the Western Isles, Scotland. J Appl Ecol 38(4):802–812
Jackson D (2006) Factors affecting the abundance of introduced hedgehogs (Erinaceus europaeus) to the Hebridean island of South Uist in the absence of natural predators and implications for nesting birds. J Zool 271(2):210–217
Jackson DB, Green RE (2000) The importance of the introduced hedgehog (Erinaceus europaeus) as a predator of the eggs of waders (Charadrii) on machair in South Uist, Scotland. Biol Conserv 93:333–348
Jackson DB, Fuller RJ, Campbell ST (2004) Long-term population changes among breeding shorebirds in the Outer Hebrides, Scotland, in relation to introduced hedgehogs (Erinaceus europaeus). Biol Conserv 117:151–166
Jensen A (2004) Overwintering of European hedgehogs Erinaceus europaeus in a Danish rural area. Acta Theriol 49:145–155
Kristiansson H (1984) Ecology of a hedgehog Erinaceus europaeus population in southern Sweden. PhD thesis, University of Lund
Met Eireann-www.met.ie
Micol T, Doncaster C, Mackinlay L (1994) Correlates of local variation in the abundance of hedgehogs Erinaceus europaeus. J Anim Ecol 63:851–860
Morris P (1969) Some Aspects on the ecology of the hedgehog (Erinaceus europaeus). PhD thesis, University of London
Morris P (1973) Winter nests of the hedgehog (Erinaceus europaeus L.). Oecologia 11:299–313
Morris P (1984) An estimate of the minimum body weight necessary for hedgehogs (Erinaceus europaeus) to survive hibernation. J Zool 203:291–294
Parkes J (1975) Some aspects of the biology of the hedgehog (Erinaceus europaeus L.) in the Manawatu, New Zealand. N Z J Zool 2:463–472
Pithon JA, Moles R, O’Halloran J (2005) The influence of coniferous afforestation on lowland farmland bird communities in Ireland: different seasons and landscape contexts. Landsc Urban Plan 71:91–103
Pollard E, Hooper MD, Moore NW (1977) Hedges. Collins, St James Place, London
Reeve NJ (1981) A field study of the hedgehog (Erinaceus europaeus) with particular reference to movements and behaviour. PhD thesis, London
Reeve NJ (1982) The home range of the hedgehog as revealed by a radio tracking study. Symp Zool Soc Lond 49:207–230
Riber AB (2006) Habitat use and behaviour of European hedgehog Erinaceus europaeus in a Danish rural area. Acta Theriol 51:363–371
Robinson RA, Sutherland WJ (2002) Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 39:157–176
Rudel TK, Coomes OT, Moran E, Achard F, Angelsen A, Xu J, Lambin E (2005) Forest transitions: towards a global understanding of land use change. Glob Environ Chang 15:23–31
Tähti H, Soivio A (1977) Respiratory and circulatory differences between induced and spontaneous arousals in hibernating hedgehogs (Erinaceus europaeus L.). Ann Zool Fenn 14:197–202
Walhovd H (1978) The overwintering pattern of Danish hedgehogs in outdoor confinement, during three successive winters. Natura Jutland 20:273–284
Walhovd H (1979) Partial arousals from hibernation in hedgehogs in outdoor hibernacula. Oecologia 40:141–153
Webb P, Ellison J (1998) Normothermy, torpor, and arousal in hedgehogs (Erinaceus europaeus) from Dunedin. N Z J Zool 25:85–90
White TA, Searle JB (2007) Factors explaining increased body size in common shrews (Sorex araneus) on Scottish Islands. J Biogeogr 34:356–363
Young AG, Port GR, Green DB (1993) Development of a forecast of slug activity: validation of models to predict slug activity from meteorological conditions. Crop Prot 12:232–236
Acknowledgements
The authors would gratefully like to acknowledge the assistance of the following people: Digger Jackson, Pat Morris, Nigel Reeve and Anouschka Hof for their invaluable advice on hedgehog research at the start of this project. Helen Bradley for all of her many hours of assistance with G.I.S., The Department of Environment, Heritage and local Government for granting licenses, Crawford Hayes fund for a PhD scholarship and all of the staff and students in U.C.C. who assisted in this study. Finally this would not have been possible without the assistance of the land owners, who co-operated throughout this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Andrzej Zalewski
Rights and permissions
About this article
Cite this article
Haigh, A., O’Riordan, R.M. & Butler, F. Nesting behaviour and seasonal body mass changes in a rural Irish population of the Western hedgehog (Erinaceus europaeus). Acta Theriol 57, 321–331 (2012). https://doi.org/10.1007/s13364-012-0080-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-012-0080-2