
Abstract
Background and Objectives
1-Aminobenzotriazole, a known time-dependent inhibitor of cytochrome P450 (CYP) enzymes, and ketoconazole, a strong inhibitor of the human CYP3A4 isozyme, are used as standard probe inhibitors to characterize the CYP and/or non-CYP-mediated metabolism of xenobiotics. In the present investigation, 1-Aminobenzotriazole and ketoconazole are characterized as potent monoamine oxidase (MAO) inhibitors in vitro using mouse, rat and human liver microsomes and S9 fractions.
Methods
Inhibition potential of 1-aminobenzotriazole and ketoconazole was studied in mice, rat and human liver microsomes, S9 fractions, MAO-A and MAO-B expressed enzymes by monitoring the formation of 4-hydroxyquinoline (4-HQ) from kynuramine, a specific substrate of MAO by liquid chromatography–tandem mass spectrometry (LC–MS/MS). Mechanism of MAO inhibition was studied by incubating varying concentration of kynuramine with mouse, rat and human S9 fractions at varying concentration of 1–aminobenzatriazole and ketoconazole and monitoring the formation of 4-HQ.
Results
1-aminobenzotriazole and ketoconazole inhibited both MAO isozymes (MAO-A and MAO-B) with more specificity towards MAO-B. Kynuramine substrate kinetics in mouse, rat and human S9 fractions with varying 1-aminobenzotriazole and ketoconazole concentrations showed decreased maximum rate (V max) for 4-HQ formation without affecting the Michaelis-Menten constant (K m). A non-competitive inhibition model was constructed and inhibition constants (K i) for 1-aminobenzotriazole (7.87 ± 0.61, 8.61 ± 0.92, 65.2 ± 1.61 µM for mice, rat and humans, respectively) and ketoconazole (0.12 ± 0.01, 2.04 ± 0.08, 5.52 ± 0.47 µM for mice, rat and humans, respectively) were determined.
Conclusions
1-Aminobenzotriazole and ketoconazole are characterized as non-competitive inhibitors of mice, rat and human MAO in vitro and the extent of their MAO inhibition potential is species specific. 1-Aminobenzotriazole or ketoconazole can be used as a probe inhibitor in vitro for screening the involvement of MAO-dependent metabolism of new chemical entities (NCE) in early drug discovery.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1-Aminobenzotriazole or Ketoconazole are inhibitors of MAO, with specificity towards MAO-B. |
1-Aminobenzotriazole or Ketoconazole show non-competitive inhibition with decrease in V max, with a species-specific potency. |
1-Aminobenzotriazole or Ketoconazole can be used as a probe inhibitor in vitro for screening the involvement of MAO-dependent metabolism of new chemical entities (NCE) in early drug discovery. |
1 Introduction
Monoamine oxidases (MAO) are flavin-containing amino oxidoreductase enzymes that are located in cytosol of mammalian cells and sub-cellular spaces such as and mitochondria [1, 2]. MAO enzymes mediate oxidative and non-oxidative deamination of endogenous amines and xenobiotics [3]. As a pharmacotherapy, selective MAO-B inhibition has been employed to treat several CNS disorders such as anxiety, depression, Parkinson’s disease (PD) and Alzheimer’s disease (AD) [4, 5]. Selegiline and rasagiline are selective irreversible MAO-B inhibitors used in the treatment of early-stage PD, depression and dementia [6]. Clorgiline, a selective irreversible MAO-A inhibitor, is used in treatment for anxiety [5]. However, MAO inhibition by selegiline, rasagiline and clorgiline may lead to severe food–drug and drug–drug interactions (DDIs) called Serotonin Syndrome, a potentially life-threatening adverse drug reaction due to excess serotonergic activity in the central nervous system (CNS) [7].
1-Aminobenzotriazole and Ketoconazole are orally bioavailable antifungal drugs characterized by triazole and imidazole moieties, respectively, in their structure [8]. 1-Aminobenzotriazole and Ketoconazole inhibit fungal cytochrome P450 (CYP) 51, an enzyme important in the biosynthesis of ergosterol, an integral component of fungal cell walls [9]. The inhibition of CYP51 by 1-Aminobenzotriazole is characterized by metabolic activation and irreversible binding to the heme moiety leading to inactive enzyme [10]. Ketoconazole inhibits the CYP51 by binding to the N-3 nitrogen of the imidazole moiety of heme of the CYP and the simultaneous binding of a lipophilic portion of the molecule to a hydrophobic region of the enzyme protein, leading to inactivation of enzyme [11]. 1-Aminobenzotriazole but not Ketoconazole, is a time-dependent inhibitor of CYP across the species [10], whereas Ketoconazole inhibits human CYP3A4 [12]. The use of 1-Aminobenzotriazole as an antifungal drug has been limited by serious adverse reactions due its broad CYP inhibition across preclinical species and in humans [12]. Ketoconazole, an orally active azole is the first approved azole-based antifungal drug in clinical practice [13]. Ketoconazole potently inhibits human CYP3A4, the enzyme responsible for the metabolism of large variety of endogenous and exogenous compounds [14–16]. Furthermore, inhibition of human CYP3A4 by Ketoconazole has been implicated in serious adverse and fatal drug–drug interactions and caused its subsequent withdrawal from the market [17].
In early drug discovery, 1-Aminobenzotriazole is used as an in vitro probe inhibitor to characterize CYP and non-CYP and Ketoconazole as probe inhibitor for characterizing CYP3A4-mediated metabolism of new chemical entities (NCEs) using liver microsomes [1, 18]. Further, 1-Aminobenzotriazole and Ketoconazole are used to study the extent of first-pass metabolism in preclinical species and DDIs specific to human CYP3A4 in clinical trials, respectively [17, 19–21]. 1-Aminobenzotriazole has been explored as a substrate and inhibitor of cytochrome c-reductase, flavin monooxygenases, Uridine diphosphate-glucuronosyltransferase and N-acetyltransferase; it has been reported as a substrate and inhibitor of N-acetyltransferase but was neither a substrate nor an inhibitor of other phase I and phase II enzymes [22–24]. Furthermore, Ketoconazole has not been reported as an inhibitor of phase II enzymes.
In the present investigation, 1-Aminobenzotriazole and Ketoconazole in the absence of NADPH were found to diminish MAO-mediated metabolism of kynuramine, a known MAO-specific probe substrate in rat liver microsomes and S9 fractions [25]. Further investigations were performed with 1-Aminobenzotriazole and Ketoconazole as potential inhibitors of 4-hydroxyquinoline (4-HQ) formation from kynuramine mediated by MAO in liver microsomes and S9 fractions. Recombinant MAO-A and B isozymes were used to confirm and characterize the isozyme-specific inhibition by 1-Aminobenzotriazole and Ketoconazole. To the best of our knowledge, this is the first in vitro investigation demonstrating 1-Aminobenzotriazole and Ketoconazole as MAO inhibitors.
2 Materials and Methods
1-Aminobenzotriazole, Ketoconazole, 8-cyclopentyl-1,3-dipropylxanthine (CPDPX), and kynuramine were procured from Sigma-Aldrich (St. Louis, MO, USA). 4-Hydroxyquinoline was procured from Alfa Aesar (Lancashire, UK). Pooled mouse liver S9 fraction (MS9), rat liver S9 fraction (RS9), human liver S9 fraction (HS9), mouse liver microsomes (MLM), rat liver microsomes (RLM) and human liver microsomes (HLM) were purchased from Xenotech, LLC (Lenexa, KS, USA). Monoamine Oxidase A (MAO-A) and monoamine oxidase B (MAO-B) Supersomes were procured from BD Gentest (Woburn, MA, USA); LCMS grade acetonitrile and methanol were obtained from JT Baker Chemicals. All the other chemicals used were of highest quality available.
2.1 Substrate Kinetics of Kynuramine in Mice, Rat and Human Liver S9 Fraction
Kynuramine is a specific MAO probe substrate. It undergoes monoamine oxidase-mediated metabolism to form 4-HQ in a two-step reaction with an aldehyde intermediate which readily undergoes condensation to form the product 4-HQ [26] (Fig. 1). 4-HQ was measured using liquid chromatography–tandem mass spectrometry (LC–MS/MS) method. Following the initial experiments by Shaik [27], the assay parameters were fixed as follows: assay buffer was potassium phosphate 100 mM (pH 7.4) containing 3 mM MgCl2. The total reaction volume was 200 µL with liver microsomes or S9 protein at final concentration of 1 mg/mL and MAO-A or MAO-B protein at a final concentration of 0.5 mg/mL. Kynuramine was incubated with the set assay conditions in a 96-well plate (Waters) with mouse, rat, or human liver microsomes or S9 fraction, and MAO-A or MAO-B Supersomes at final assay concentrations of 0.04, 0.09, 0.19, 0.39, 0.78, 1.56, 3.12, 6.25, 12.5, 25.0, 50.0 and 100.0 µM in a humidified orbital shaker water bath at 37 °C for 30 min. Each of the concentrations was incubated in triplicate. At the end of each incubation, the reaction was terminated by adding 200 µL of acetonitrile containing 10 ng/mL of analytical internal standard (IS) CPDPX. The contents of the tube were transferred into a 96-well 1-mL plate (Millipore), the plate was centrifuged at 4000 g for 10 min in a tabletop centrifuge (Eppendorf 5810 R), and 100 µL of supernatant was transferred into a 96-well deep well analysis plate with 2-mL volume capacity (Millipore) and 200 µL of diluent was added containing a mixture of acetonitrile:MilliQ:formic acid in the ratio 80:20:0.1. The plates were loaded to an API 5500 LC–MS/MS (AB Sciex, Framingham, MA) with an auto sampler for analysis. Formation of 4-HQ from kynuramine was measured and quantified against the calibration curve using the LC–MS/MS method. Calibration curve was prepared for 4-HQ, the linear regressions with weighting factor 1/X2 of 0.99 or greater were used for calculating the 4-HQ content.

Monoamine oxidase (MAO)-mediated metabolism of kynuramine (1), kynuramine undergoes oxidative deamination to aldehyde (2), followed by condensation to form 4-hydroxyquinoline (3)
The incubations were performed on different days and each time the incubation was performed a new calibration curve was run with the samples to quantify 4-HQ and calculate enzyme kinetic parameters such as Michaelis-Menten constant (K m) and maximum rate (V max).
2.2 Screening of 1-Aminobenzotriazole and Ketoconazole for MAO Inhibition
1-Aminobenzotriazole and Ketoconazole were screened as MAO inhibitors using mouse liver S9 fractions, MAO-A or MAO-B supersomes at a final concentration of 10 µM using set assay conditions as mentioned above. In brief, 1-Aminobenzotriazole and Ketoconazole at a final concentration of 10 µM were pre-incubated with assay buffer (potassium phosphate 100 mM containing (pH 7.4) 3 mM MgCl2) and liver microsomes and S9 fractions for 10 min in triplicate in a humidified orbital shaker water bath at 37 °C. The total reaction volume was 200 µL. The reaction was initiated by the addition of kynuramine at a final concentration of 10 µM and incubated in a humidified orbital shaker water bath at 37 °C for 30 min. At the end of incubation, the reaction was terminated, processed and analyzed as illustrated in Sect. 2.1. The formation of 4-HQ was measured by LC–MS/MS and compared in the presence and absence of 1-Aminobenzotriazole or Ketoconazole.
2.3 Mechanism of Inhibition of MAO by 1-Aminobenzotriazole or Ketoconazole
The inhibition constants, K i, for 1-Aminobenzotriazole or Ketoconazole were determined for MAO using mouse, rat and human liver S9 fraction by incubating 1-Aminobenzotriazole or Ketoconazole with assay buffer (potassium phosphate 100 mM containing (pH 7.4) 3 mM MgCl2) and liver S9 at a final concentration of 1 mg/mL for 10 min in triplicate in a humidified orbital shaker water bath at 37 °C. The final concentrations of 1-Aminobenzotriazole were 0.00, 3.12, 6.25, 12.5, 25.0, 50, 100, and 200 µM for mouse, rat and human microsomes and S9 fraction. The final concentrations of Ketoconazole were 0.156, 0.312, 0.625, 1.25, 2.5, 5.0 and 10.0 µM for mouse liver microsomes and S9 fraction and 0.00, 0.78, 1.56, 3.12, 6.25, 12.5, 25.0 and 50 µM for rat and human S9 fraction or 1-Aminobenzotriazole (0.00, 3.12, 6.25, 12.5, 25.0, 50, 100 and 200 µM for all species). The assay was initiated by the addition of kynuramine at a final concentration of 0.78, 1.56, 3.12, 6.25, 12.5, 25.0, 50 and 100 µM at each concentration of 1-Aminobenzotriazole and Ketoconazole. The total reaction volume was 200 µL and incubated in a humidified orbital shaker water bath at 37 °C for 30 min. At the end of incubation, the reaction was terminated, processed and analyzed as illustrated above. The formation of 4-HQ was determined at varied concentrations of 1-Aminobenzotriazole and Ketoconazole against varied concentrations of kynuramine.
3 Bioanalysis
3.1 LC–MS/MS Method
A Shimadzu Prominence LC (Shimadzu Corporation, Kyoto, Japan) system attached to an API-5500-QTrap mass spectrometer (AB Sciex, Framingham, MA, USA) equipped with a Turboionspray™ interface (Electrospray Ionization-ESI) set at 500 °C was used for analysis of samples. Quantitation of 4-HQ was achieved by MS–MS detection in positive ion mode. The quadrupoles Q1 and Q3 were set on unit resolution, detection of all the ions was performed in the multiple reaction monitoring (MRM) mode and the analytical data were processed by Multiquant® software version 2.1 (AB Sciex, Framingham, MA, USA). Chromatographic separation of kynuramine and 4-HQ is achieved upon injecting 20 µL of processed samples and 4-HQ calibration samples on a Kinetex C18 column (2.1 × 100 mm, 5 µm, Phenomenex, Torrance, CA). A binary gradient mobile phase consisting of A) 0.1% formic acid in water (MilliQ water: Millipore, Milford, MA, USA) and B) 100% acetonitrile was delivered at 0.50 mL/min with a gradient flow with a run time of 4 min, started with 90% A for 0.5 min (0–0.5 min), changed to 40% A for the next 0.5 min (0.5–1.0 min), changed to 5% A for next 2.0 min (1.0–3.0 min), changed back to 90% A in next 0.5 min (3.0–3.5 min) and composition was held at 90% A for next 0.5 min (3.5–4.0 min); for the first 1 min and the last minute the eluent was run into waste, from 1.0 to 3.0 min the eluent plus the separated compounds were delivered into the mass spectrometer’s electrospray ionization chamber. The effluent was ionized by positive ion mode and detected by mass spectrometry.
Quantitation of 4-HQ was achieved by MS–MS detection in positive ion mode for 4-HQ and the internal standard (CPDPX). The ion spray voltage was set at 4500 V. The other parameters such as curtain gas, collision gas, ion source gas 1 (GS1) and GS2 were set at 32, medium, 60 and 70, respectively. The ion pairs of 4-HQ are 147.1 → 78.1, and CPDPX are 305.1 → 178.1. The compound-specific parameters such as declustering potential (DP), entrance potential (EP), collision energy (CE) and collision exit potential (CXP) are 95, 10, 45, 15 for 4-HQ and 216, 10, 47, 12, for CPDPX, the retention time for 4-HQ was 0.54 min and for CPDPX was 1.02 min. Detection of all ions was performed in the Multiple Reaction Monitoring (MRM) mode. The quadrupoles Q1 and Q3 were set on unit resolution and the analytical data were processed by Multiquant® software version 2.1 (AB Sciex, Framingham, MA, USA).
The 4-HQ curve was constructed for each experiment and average regression of each calibration was above 0.99 and was reproducible in the concentration range of 1–2000 nM. The US Food and Drug Administration (FDA) bioanalytical guidance was followed for the acceptance criterion, which was ±15% deviation from the theoretical values of quality control samples, low (5 nM), medium (800 nM) and high QC (1200 nM) except at lowest limit of quantitation (LLOQ), which was set at ±20% [28–30].
4 Data Analysis
The 50% inhibitoy concentration (IC50), kinetic parameters (K m, V max, and K i) for 4-HQ formation were evaluated by fitting the data in Prism software (GraphPad Software Inc., La Jolla, CA, USA). The results represent mean ± standard deviation (SD) of three independent experiments.
5 Results
5.1 Substrate Kinetics of Kynuramine with MLM, RLM, HLM, MS9, RS9, HS9, MAO-A and MAO-B-Expressed Isozymes
MAO mediates the oxidative deamination of kynuramine to 4-HQ (Fig. 1). K m and V max for kynuramine in mouse, rat and human liver S9 fractions and microsomes, and MAO-A and MAO-B-expressed isozymes were computed by plotting the rate of formation of 4-HQ with respect to the kynuramine concentration using a non-linear regression model (Fig. 2a, b). The kinetic parameters are summarized in Table 1.

Substrate kinetics of kynuramine. Formation of 4-hydroxyquinoline with increase concentration of kynuramine in a mice, rat and human S9 fractions and liver microsomes, b MAO-A-expressed enzyme and MAO-B-expressed enzyme. HLM human liver microsomes, MAO monoamine oxidase, MLM mouse liver microsomes
5.2 Determination of MAO Inhibition In Vitro in MS9, RS9, HS9, MAO-A and MAO-B-Expressed Isozymes by 1-Aminobenzotriazole and Ketoconazole
1-Aminobenzotriazole and Ketoconazole inhibited mice, rat and human MAO activity in S9 fraction (Supplemental Figure 1) and IC50 values are summarized in Supplemental Table 1 (Supplemental Figure 2). The inhibitory potency of 1-Aminobenzotriazole and Ketoconazole on MAO is in the rank order of Human < Rat < Mouse, with lowest IC50 value for mouse MAO inhibition with 1-Aminobenzotriazole and Ketoconazole as 1.98 ± 0.21 and 0.48 ± 0.06 µM, respectively. Ketoconazole and 1-Aminobenzotriazole inhibited MAO activity across the species with Ketoconazole being the more potent inhibitor compared to 1-Aminobenzotriazole. Further, 1-Aminobenzotriazole and Ketoconazole specifically inhibited MAO-B compared to MAO-A (Supplemental Figure 3) with threefold selectivity.
5.3 Mechanism of Inhibition of MAO by 1-Aminobenzotriazole and Ketoconazole
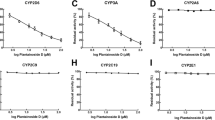
The mechanism of MAO inhibition by 1-Aminobenzotriazole and Ketoconazole was characterized by studying the effect of varying their concentrations on kynuramine K m and V max in mouse, rat and human S9 fractions with and without pre-incubation with inhibitor. The MAO inhibition potential of 1-Aminobenzotriazole and Ketoconazole in MS9, RS9 and HS9 was found to be similar to pre-incubation and co-incubation with kynuramine (data not shown), ruling out the possibility of time-dependent inhibition of MAO. Lineweaver–Burk plots were constructed for the MAO inhibition data of 1-Aminobenzotriazole and Ketoconazole in MS9, RS9, and HS9, which showed a decrease in kynuramine V max without change in K m. A non-competitive inhibition model was constructed using Graphpad prism 6 and the inhibition constant (K i) was computed for 1-Aminobenzotriazole in MS9, RS9, and HS9 (Fig. 3a–c, respectively) and Ketoconazole in MS9, RS9, and HS9 (Fig. 4a–c, respectively). The 1-Aminobenzotriazole and Ketoconazole K i values for mouse, rat and human MAO in S9 fractions are 7.87 ± 0.61 and 0.12 ± 0.01, 8.61 ± 0.92 and 2.04 ± 0.08, and 65.2 ± 1.61 and 5.52 ± 0.47 µM, respectively, as summarized in Table 2.

Inhibition of 4-hydroxyquinoline in the presence of 1-aminobenzotriazole in a mice S9 fractions, b rat S9 fractions, c human S9 fractions. Insets Lineweaver–Burk plot of inhibition of 4-hydroxyquinoline (4-HQ) in respective S9 fractions

Inhibition of 4-hydroxyquinoline in the presence of ketoconazole in a mice S9 fractions, b rat S9 fractions, c in human S9 fractions. Insets Lineweaver–Burk plot of inhibition of 4-hydroxyquinoline (4-HQ) in respective S9 fractions
6 Discussion
MAOs are expressed as MAO-A and MAO-B isozymes which play a significant role in mediating the oxidative and non-oxidative deamination of endogenous amines and xenobiotics. MAO inhibitors such as clorgiline, selegiline, rasagiline and carbidopa have been employed therapeutically in the treatment of several CNS disorders such as anxiety, depression, PD and AD [5, 6, 31]. Further, the oxazolidinone class of antibiotics exhibits non-specific inhibition of MAO isozymes leading to adverse effects [32]. In the present investigation, 1-Aminobenzotriazole, a suicide inhibitor of CYPs, and Ketoconazole, a human CYP3A4 inhibitor, were found to inhibit the MAO-mediated metabolism of kynuramine in the absence of reduced NADPH in rat liver microsomes and S9 fractions. In this paper, we systematically investigated the 1-Aminobenzotriazole and Ketoconazole inhibition potential on MAO enzymatic activity across the species (mouse, rat and human) using liver S9 fractions and microsomes as a source of MAO and kynuramine as substrate.
Kynuramine is a metabolite of indole amino acids and indolamines and it is deaminated by MAO to form an aldehyde which condenses to form 4-HQ as illustrated in Fig. 1. Kynuramine is a sensitive and specific probe substrate for characterizing the MAO activity by monitoring the formation of 4-HQ, by either by fluorometric or LC–MS/MS methods [33]. Even though fluorimetric analysis is the method of choice for determination of 4-HQ, an LC–MS/MS method was developed and used to study 4-HQ formation, to avoid spurious results due to quenching effects of inhibitors on 4-HQ signal in fluorimetric assay.
Kynuramine substrate kinetics was characterized by incubating it at various concentrations with mouse, rat and human liver S9 fractions and microsomes and with MAO-A and MAO-B-expressed enzyme. Kynuramine is metabolized to the same extent in the liver S9 and microsome fraction of all the species and expressed MAO-A and MAO-B enzyme with similar K m (~10 µM) and V max (Table 1). Further, kynuramine is metabolized in microsomes in all species, suggesting MAO activity in the endoplasmic reticulum across the species, which is contradictory to the current literature, which could be due to cross contamination during microsomes preparation and needs further investigation.
Recently, 1-Aminobenzotriazole was reported to competitively inhibit rat N-acetyl transferase (NAT) by Sun et al. [22]. In the present investigation, 1-Aminobenzotriazole and Ketoconazole inhibited kynuramine metabolism, a specific MAO substrate, in mouse, rat and human liver S9 fractions and microsomes at 10 µM. Further, inhibition of MAO by 1-Aminobenzotriazole or Ketoconazole is not time dependent, ruling out mechanism-based inhibition, which is in contrast to CYP inhibition by 1-Aminobenzotriazole [23]. MAO-A and MAO-B are expressed in many tissues, with MAO-B expression predominantly in brain [2]. 1-Aminobenzotriazole and Ketoconazole inhibit kynuramine metabolism by MAO-A and MAO-B, with specificity towards MAO-B, suggesting 1-Aminobenzotriazole as an effective inhibitor of MAO-B. In summary, 1-Aminobenzotriazole and Ketoconazole inhibit MAO activity across the species with Ketoconazole being a more potent inhibitor compared to 1-Aminobenzotriazole in vitro.
Consistent with MAO inhibition potential by 1-Aminobenzotriazole or Ketoconazole in preclinical species and humans in vitro, we further evaluated the IC50 and K i by incubating varying concentrations of 1-Aminobenzotriazole or Ketoconazole at varying concentration of kynuramine in mouse, rat and human S9 fractions. Ketoconazole showed lower K i values compared to 1-Aminobenzotriazole, confirming Ketoconazole as a potent inhibitor of MAO than 1-Aminobenzotriazole. A Lineweaver–Burk plot (Figs. 3a–c, 4a–c) for 1-Aminobenzotriazole and Ketoconazole showed an unchanged K m and a decrease in V max, which is characteristic of non-competitive inhibition. Pre-incubation or co-incubation of substrate with inhibitor did not show any change in inhibition (data not shown), hence ruling out time-dependent inhibition. 1-Aminobenzotriazole and Ketoconazole for the first time were characterized as MAO inhibitors. Further studies are required to translate the MAO in vitro inhibition potential of 1-Aminobenzotriazole and Ketoconazole in vivo.
Use of 1-Aminobenzotriazole and Ketoconazole as antifungal drugs has been discontinued due to their broad CYP inhibition across the species and especially human CYP 3A4, leading to fatal DDIs. However, with the characterization of 1-Aminobenzotriazole and Ketoconazole as MAO inhibitors in the present investigation [34, 35], 1-Aminobenzotriazole and Ketoconazole may likely be reconsidered as MAO-B inhibitors for the treatment of CNS disorders such as anxiety, depression, PD and AD by increasing their exposure in brain at a reduced dose and managing the adverse effects.
1-Aminobenzotriazole is used as an in vitro and in vivo probe inhibitor to characterize CYP and non-CYP-mediated metabolism of new chemical entities (NCE). Ketoconazole is used as an in vitro probe inhibitor for characterizing CYP3A4-mediated metabolism of NCEs using human liver microsomes [12] and further elucidating the drug–drug interaction potential specific to human CYP 3A4 [17, 19]. Based on the present investigation, 1-Aminobenzotriazole or Ketoconazole inhibits MAO in the absence of reduced NADPH and without pre-intubation, which is in contradiction to their mode of CYP inhibition [36], hence 1-Aminobenzotriazole or Ketoconazole can be used in early drug discovery to characterize the role of MAO inhibition in vitro using S9 fractions of various species based on the flowchart (Supplemental Figure 4) and their utility as in vivo MAO probe inhibitors needs to be evaluated.
7 Conclusions
1-Aminobenzotriazole and Ketoconazole are characterized as non-competitive inhibitors of mouse, rat and human MAO in vitro and the extent of their MAO inhibition potential is species specific. 1-Aminobenzotriazole or Ketoconazole can be used as a probe inhibitor in vitro for screening the involvement of MAO-dependent metabolism of new chemical entities (NCE) in early drug discovery. Further, care should be taken when either 1-Aminobenzotriazole or Ketoconazole is used as in vitro or in vivo non-specific CYP or human CYP 3A4 inhibitors, respectively, since 1-Aminobenzotriazole also inhibits MAO and NAT and Ketoconazole inhibits MAO.
References
Dalmadi B, Leibinger J, Szeberenyi S, Borbas T, Farkas S, Szombathelyi Z, et al. Identification of metabolic pathways involved in the biotransformation of tolperisone by human microsomal enzymes. Drug Metab Dispos. 2003;31(5):631–6.
Sivasubramaniam SD, Finch CC, Rodriguez MJ, Mahy N, Billett EE. A comparative study of the expression of monoamine oxidase-A and -B mRNA and protein in non-CNS human tissues. Cell Tissue Res. 2003;313(3):291–300. doi:10.1007/s00441-003-0765-6.
Furnes B, Schlenk D. Extrahepatic metabolism of carbamate and organophosphate thioether compounds by the flavin-containing monooxygenase and cytochrome P450 systems. Drug Metab Dispos. 2005;33(2):214–8. doi:10.1124/dmd.104.000984.
Strolin Benedetti M, Tipton KF. Monoamine oxidases and related amine oxidases as phase I enzymes in the metabolism of xenobiotics. J Neural Transm Suppl. 1998;52:149–71.
Yamada M, Yasuhara H. Clinical pharmacology of MAO inhibitors: safety and future. Neurotoxicology. 2004;25(1–2):215–21.
Magyar K. The pharmacology of selegiline. Int Rev Neurobiol. 2011;100:65–84.
Huang V, Gortney JS. Risk of serotonin syndrome with concomitant administration of linezolid and serotonin agonists. Pharmacotherapy. 2006;26(12):1784–93.
Como JA, Dismukes WE. Oral azole drugs as systemic antifungal therapy. N Engl J Med. 1994;330(4):263–72. doi:10.1056/NEJM199401273300407.
Lepesheva GI, Waterman MR. CYP51—the omnipotent P450. Mol Cell Endocrinol. 2004;215(1–2):165–70.
Ortiz de Montellano PR. Mathews JM. Autocatalytic alkylation of the cytochrome P-450 prosthetic haem group by 1-aminobenzotriazole. Isolation of an NN-bridged benzene–protoporphyrin IX adduct. Biochem J. 1981;195(3):761–4.
Strushkevich N, Usanov SA, Park HW. Structural basis of human CYP51 inhibition by antifungal azoles. J Mol Biol. 2010;397(4):1067–78.
Wrighton SA, Ring BJ. Inhibition of human CYP3A catalyzed 1′-hydroxy midazolam formation by ketoconazole, nifedipine, erythromycin, cimetidine, and nizatidine. Pharm Res. 1994;11(6):921–4.
Nimura K, Niwano Y, Ishiduka S, Fukumoto R. Comparison of in vitro antifungal activities of topical antimycotics launched in 1990s in Japan. Int J Antimicrob Agents. 2001;18(2):173–8.
Guengerich FP. Role of cytochrome P450 enzymes in drug-drug interactions. Adv Pharmacol. 1997;43:7–35.
Li AP, Kaminski DL, Rasmussen A. Substrates of human hepatic cytochrome P450 3A4. Toxicology. 1995;104(1–3):1–8.
Basu S, Shaik AN. Is there a paradigm shift in use of microsomes and hepatocytes in drug discovery and development? ADMET DMPK. 2016;4(2):114–6.
Honig PK, Wortham DC, Zamani K, Conner DP, Mullin JC, Cantilena LR. Terfenadine–Ketoconazole interaction. Pharmacokinetic and electrocardiographic consequences. JAMA. 1993;269(12):1513–8.
Eagling VA, Tjia JF, Back DJ. Differential selectivity of cytochrome P450 inhibitors against probe substrates in human and rat liver microsomes. Br J Clin Pharmacol. 1998;45(2):107–14.
Lin JH, Lu AY. Inhibition and induction of cytochrome P450 and the clinical implications. Clin Pharmacokinet. 1998;35(5):361–90.
Balani SK, Li P, Nguyen J, Cardoza K, Zeng H, Mu DX, et al. Effective dosing regimen of 1-aminobenzotriazole for inhibition of antipyrine clearance in guinea pigs and mice using serial sampling. Drug Metab Dispos. 2004;32(10):1092–5.
Balani SK, Zhu T, Yang TJ, Liu Z, He B, Lee FW. Effective dosing regimen of 1-aminobenzotriazole for inhibition of antipyrine clearance in rats, dogs, and monkeys. Drug Metab Dispos. 2002;30(10):1059–62.
Sun Q, Harper TW, Dierks EA, Zhang L, Chang S, Rodrigues AD, et al. 1-Aminobenzotriazole, a known cytochrome P450 inhibitor, is a substrate and inhibitor of N-acetyltransferase. Drug Metab Dispos. 2011;39(9):1674–9.
Mugford CA, Mortillo M, Mico BA, Tarloff JB. 1-Aminobenzotriazole-induced destruction of hepatic and renal cytochromes P450 in male Sprague-Dawley rats. Fundam Appl Toxicol. 1992;19(1):43–9.
Cashman JR. A convenient radiometric assay for flavin-containing monooxygenase activity. Anal Biochem. 1987;160(2):294–300.
Makino K, Arai K. Conversion of 3-hydroxykynurenine to 4,8-dihydroxyquinoline by mouse liver homogenate. Science. 1955;121(3135):143–4.
Weissbach H, Smith TE, Daly JW, Witkop B, Udenfriend S. A rapid spectrophotometric assay of mono-amine oxidase based on the rate of disappearance of kynuramine. J Biol Chem. 1960;235:1160–3.
Shaik AN, Bohnert T, Williams DA, Gan LL, LeDuc BW. Mechanism of drug–drug interactions between warfarin and statins. J Pharm Sci. 2016;105(6):1976–86.
Shaik AN, Grater R, Lulla M, Williams DA, Gan LL, Bohnert T, et al. Comparison of enzyme kinetics of warfarin analyzed by LC–MS/MS QTrap and differential mobility spectrometry. J Chromatogr B. 2016;1008:164–73.
FDA C. Guidance for industry: bioanalytical method validation. US Department of Health and Human Services. Food and Drug Administration, Center for Drug Evaluation and Research (CDER), Center for Veterinary Medicine (CV). 2001.
Basha SJ, Naveed SA, Tiwari NK, Shashikumar D, Muzeeb S, Kumar TR, et al. Concurrent determination of ezetimibe and its phase-I and II metabolites by HPLC with UV detection: quantitative application to various in vitro metabolic stability studies and for qualitative estimation in bile. J Chromatogr B Anal Technol Biomed Life Sci. 2007;853(1–2):88–96.
Strolin Benedetti M, Sanson G, Bona L, Gallina M, Persiani S, Tipton KF. The oxidation of dopamine and epinine by the two forms of monoamine oxidase from rat liver. J Neural Transm Suppl. 1998;52:233–8.
Lawrence KR, Adra M, Gillman PK. Serotonin toxicity associated with the use of linezolid: a review of postmarketing data. Clin Infect Dis. 2006;42(11):1578–83.
Weissbach H, Robertson AV, Witkop B, Udenfriend S. Rapid spectrophotometric assays for snake venom l-amino acid oxidase based on the oxidation of l-kynurenine or 3,4-dehydro-l-proline. Anal Biochem. 1960;1:286–90.
Shaik AN, Mahat MY, Kandasamy K, LeDuc BW, Khan AA, Azoles as novel inhibitors of monoamine oxidase (MAO) A and B isozymes. Drug Metab Rev, 2015;47(supl):204.
Shaik AN, LeDuc BW, Khan AA. 1-Aminobenzotriazole and ketoconazole—non competitive inhibitors of monoamine oxidase. Drug Metab Rev. 2016;48(Sup 1):128.
de Montellano PRO, Mico BA, Mathews JM, Kunze KL, Miwa GT, Lu AY. Selective inactivation of cytochrome P-450 isozymes by suicide substrates. Arch Biochem Biophys. 1981;210(2):717–28.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
No funding has been received for the conduct of this study.
Conflict of interest
The authors report no conflict of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Shaik, A.N., LeDuc, B.W. & Khan, A.A. Characterization of 1-Aminobenzotriazole and Ketoconazole as Novel Inhibitors of Monoamine Oxidase (MAO): An In Vitro Investigation. Eur J Drug Metab Pharmacokinet 42, 827–834 (2017). https://doi.org/10.1007/s13318-017-0401-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13318-017-0401-6