
Abstract
The rust pathogen, Puccinia psidii, was first detected in South Africa in 2013 on a single non-native ornamental Myrtus communis tree. This prompted surveys of the country to determine its geographic distribution and host range. Previously developed microsatellite markers where used to characterize the genetic diversity of P. psidii isolates collected from these surveys. In addition, artificial inoculation studies and field observations were used to evaluate the susceptibility of native Myrtaceae to infection by P. psidii. The pathogen was found on native Myrtaceae in isolated natural situations and it was also common on exotic Myrtaceae in nurseries and gardens. Marker analysis showed that a single genotype of the rust is present in South Africa and that this is different to the so-called “pandemic” strain recorded in countries outside Brazil. It was found to have a broad distribution in South Africa with collections as far as 1500 km apart. The data provide firm evidence for a single introduction of the pathogen from an as yet unkown source. Its wide distribution, particularly in relatively isolated natural areas, suggests that P. psidii has been present in South Africa for much longer than implied by its first detection in the country.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The rust pathogen, Puccinia psidii Winter (Urediniales, Sphaerophragmiaceae), has been described as “the greatest threat to the ecosystem” by Australian scientists and newspapers (Dayton and Higgins 2011; Glen et al. 2007; Pegg et al. 2014b). Subsequent to its first detection in Australia (Carnegie et al. 2010), P. psidii has led to the near extinction of 12 species of native Australian Myrtaceae (Carnegie et al. 2016; Pegg et al. 2014b). The pathogen has also had a significant impact on agriculture and commercial plantation forestry in other regions where it occurs, devastating the allspice (Pimenta dioica) industry in Jamaica (MacLachlan 1938) and resulting in considerable management costs to the eucalypt industry in Brazil (Alfenas et al. 2004; Ferreira 1983; Graça et al. 2011).
Puccinia psidii was first reported in 1884 from southern Brazil, infecting Psidium guajava (Winter 1884). The fungus has subsequently been reported from North America (Marlatt and Kimbrough 1979) and Hawaii (Uchida et al. 2006), China (Zhuang and Wei 2011), Japan (Kawanishi et al. 2009), Australia (Carnegie et al. 2010), Africa (Roux et al. 2013), New Caledonia (Giblin 2013; Machado et al. 2015) and Indonesia (McTaggart et al. 2016). All of these reports, outside Brazil, were attributed to a single genoptype of P. psidii (Machado et al. 2015), referred to as the ‘pandemic’ genotype (Ross-Davis et al. 2013). It is widely believed that P. psidii is native to Latin America due to its long history in that region and the fact that native Myrtaceae in the region are generally tolerant to infection, as opposed to the susceptible introduced species of Myrtaceae (Coutinho et al. 1998; Glen et al. 2007). Recent population genetic studies on collections of P. psidii from the Americas also suggest that host specific genotypes are native to Central and South America (Graça et al. 2013; Zhong et al. 2008, 2011).
For many years it was believed that P. guajava (guava) was the host of origin of P. psidii and that it had undergone a host jump to non-native Eucalyptus and other Myrtaceae (Alfenas et al. 2005; Castro de et al. 1983; Tommerup et al. 2003). However, a study by Graça et al. (2013) showed that isolates from guava and eucalypts in Brazil represented distinct genotypes/biotypes of the pathogen. The source population of the rust that occurs on eucalypts remains unknown although it seems likely that this would be in South or Central America.
Puccinia psidii has been reported from more than 73 genera and 450 species of plants in the Myrtaceae (Pegg et al. 2014b; Giblin and Carnegie 2014; Zauza et al. 2015). This plant family includes approximately 140 genera accommodating about 5500 species and it occurs worldwide (Biffin et al. 2010). Three genera of the Myrtaceae occur in South Africa, namely Eugenia, Metrosideros and Syzygium (Coates Palgrave 2002). A closely related genus, Heteropyxis, is endemic to southern Africa and has been classified either in its own family, the Heteropyxidaceae, or a sub-family within the Myrtaceae (Stevens 2012; Wilson et al. 2005). Of the South African species of Myrtaceae, only H. natalensis has been tested for its susceptibility to P. psidii. Artificial inoculation trials in Brazil showed that H. natalensis was highly susceptible to the genotype of rust fungus used in that study (Alfenas et al. 2005). Despite the relatively low number of native Myrtaceae in South Africa and other African countries, compared to Australia and South America, these plants are important components of the natural ecosystem, providing fruits for humans and animals, timber and medicinal compounds (Coates Palgrave 2002).
Puccinia psidii was first detected in South Africa in May 2013, on a single Myrtus communis tree, in a private garden on the south coast of KwaZulu-Natal (Roux et al. 2013). Subsequent to its detection, many articles have been published in garden and botanical magazines as well as in the media to raise awareness about the disease in South Africa (Roux 2015; Roux and Pegg 2014). Disease alerts and regular updates of Myrtle rust have also been provided to the South African forestry industry through newsletters and the web (http://fabinet.up.ac.za/archive). The aims of this study were to determine the distribution of P. psidii subsequent to its first report in South Africa; determine whether the pandemic genotype of P. psidii is present in South Africa by comparison with specimens from Australia using microsatellite markers, and evaluate the relative susceptibility of native South African Myrtaceae to P. psidii using inoculation studies under greenhouse conditions.
Materials and methods
Disease surveys and sample collections
Surveys of Eucalyptus plantations, nurseries and native stands of Myrtaceae were conducted in 2013, 2014 and 2015 in order to assess the current distribution of P. psidii in South Africa. Sites were selected based on occurrence maps of native Myrtaceae (www.sanbi.org) published by the South African National Biodiversity Institute (SANBI) and as identified by Roux et al. (2015) as high risk areas for the occurrence of P. psidii in the country. These included areas along the coast of the KwaZulu-Natal Province, the Mbombela region in the Mpumalanga Province and the Tzaneen and Soutpansberg regions in the Limpopo Province. Reports of possible infected plants in private gardens, made by the general public, were investigated after awareness campaigns in the media and garden forums. Eucalypt nurseries and plantations in high risk areas were visited on an ad hoc basis, and monitored by forestry staff.
Fragment analyses and genotyping
Seven P. psidii samples collected from countrywide surveys, as well as four isolates from Australia, were used in the genetic analyses (Table 1). Urediniospores were harvested from infected leaf samples using a vacuum pump and stored in 2 mL screw- cap vials at −80 °C prior to DNA extraction. The genotypes of samples were determined using seven microsatellite markers (PpSS012, PpSS014, PpSS018, PpSS022, PpSS102, PpSS161 and PpSS195) developed by Zhong et al. (2008) and modified by Graça et al. (2013). Genomic DNA was extracted from a single uredinium per host using the Ultraclean® Microbial DNA Isolation Kit (MoBio Laboratories, Carlsbad, California, USA). PCR mixtures included 1× PCR FastStart Taq Buffer with MgCl2 (Sigma-Aldrich, St. Louis, Missouri, USA), 200 μM dNTPs, 0.1 μM forward (labeled with NED™, FAM™, PET® or VIC™ fluorescent dye) and reverse primers, 1 unit FastStart Taq DNA polymerase (Sigma-Aldrich), and DNA template in 12,5 μl reaction volumes. PCR amplifications were made using a Veriti® Thermal Cycler (Life Technologies, Carlsbad, California, USA) with one cycle at 95 °C for 5 min, followed by three cycles at 95 °C for 30s, 52–56 °C (depending on the optimal annealing temperature per primer pair) for 30s, 72 °C for 80s, then an additional 35 cycles at 94 °C for 15 s, 52–56 °C for 15 s and 45 s at 72 °C, and final extension for 5 min at 72 °C. Fragment analyses were performed using an ABI Applied Biosystems 3500xl sequencer and Liz (-250) size standard (Thermo Fisher Scientific, Carlsbad, USA) at the Sequencing Facility of the Faculty of Natural and Agricultural Science, University of Pretoria. Alleles were scored and sizes determined using GeneMapper® Software Version 4.1 (Applied Biosystems, Thermo Fisher Scientific, Carlsbad, USA).
Inoculation studies
Native South African Myrtaceae were sourced from nurseries in the Gauteng, KwaZulu-Natal and Limpopo Provinces and acclimatized to greenhouse conditions at the University of Pretoria prior to inoculation with P. psidii. Five plants each of six native species (Eugenia erythrophyllum, E. verdoornii, Heteropyxis canescens, H. natalanesis, Syzygium cordatum, S. legatii) were obtained for inoculation studies. Four weeks prior to inoculation, branch tips were pruned back to produce even-aged, new-growth shoots that would be most susceptible to infection.
Inoculum was sourced from a natural P. psidii infection on Eugenia natalitia seedlings near Tzaneen in the Limpopo Province. Spores were harvested with a fine-haired paint brush and suspended in 1 mL of sterile distilled water (SDW) with 0.05 % Tween 20. This mixture was applied to new leaves of Syzygium jambos for propagation of the spores. The plants were watered and covered with clear plastic bags in a glasshouse for 48 h to maintain humidity and facilitate infection, then grown under glasshouse conditions at ~25 °C for two weeks. Inoculations were done such that the first 12 h after inoculation were at night, thus providing darkness necessary for spore germination. The spores were harvested using a vacuum pump, desiccated for 5–7 days, and stored at -80 °C.
Desiccated P. psidii urediniospores were removed from -80 °C storage and added to sterile distilled water (SDW) with 0.05 % Tween 20. The spore suspension was incubated at room temperature to facilitate rehydration of the spores and mixed to ensure an even distribution of the inoculant. Spore counts were made using a haemocytometer and the suspension adjusted to a concentration of 1 × 105 spores/ml. Native plants were inoculated with a fine mist of the spore suspension using an artist airbrush (Iwata LPH-80) powered by a 1/6 HP tubular compressor (Iwata IS 875HT Smart Jet Plus). The inoculum was applied to both the abaxial and adaxial leaf surfaces until just before the point of run off following the method described by Pegg et al. (2014a). Syzygium jambos plants were included as indicators of positive infection. The plants were assessed for symptoms after 14 days and rated for disease resistance based on the scale developed by Junghans et al. (2003a).
Results
Disease surveys and sample collections
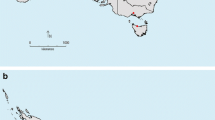
Puccinia psidii was present in the Gauteng, KwaZulu-Natal and Limpopo Provinces of South Africa (Fig. 1). In all areas, both uredinia and telia were common (Fig. 2). Puccinia psidii was common on non-native Myrtus communis in commercial ornamental nurseries that stocked this plant in the Gauteng Province (four nurseries). The only other report from a non-native host was from a Backhousia citriodora plant found in a private garden in Gauteng.

Map of South Africa showing areas from which Puccinia psidii has been confirmed (Base map produced by Dr. Ilaria Germizhuizen, ICFR, South Africa)

Typical symptoms of Puccinia psidii infection on Myrtaceae in South Africa. a Leaf spots with yellow masses of uredinia and rust brown telia on Eugenia erythrophyllum, b telia on E. erythrophyllum, c teliospore and urediniospores on a Myrtus communis leaf, d dying leaves and shoot of E. erythrophyllum covered with masses of yellow uredinia, e infected flower buds of Backhousia citriodora, f young, infected shoots of E. umtamvunensis with uredinia
New reports from KwaZulu-Natal (KZN) include one from a private garden in the town of Melmoth in eastern KZN, from M. communis, and a second from native Myrtaceae in a private nursery on the south coast of the province. Infected hosts in the nursery included Eugenia erythrophylla, E. natalitia, E. umtamvunensis and Heteropyxis natalensis (Table 2). During two previous visits to the nursery in 2013 and 2014, no rust had been observed on these plants, but during March of 2015 the disease was found on several plants of each of the species.
In the Limpopo, P. psidii was found only on native E. natalitia and only in a natural forest. The first discovery of the pathogen in the Province was in the Wolkberg Wilderness area near Tzaneen, on two plants. Subsequent visits to the region revealed infections of the fungus at a number of other sites, all in native forest patches and only on E. natalitia. The pathogen was also found on E. natalitia at two sites in the Soutpansberg Mountains, i.e. at the western most point of the mountain range near the town of Vivo and at the eastern most part of the range near the city of Thohoyandou (Fig. 1).
No reports of possible Puccinia psidii symptoms on plantation Eucalyptus species in South Africa were received during the period 2013–2015. Visits to commercial plantation forestry nurseries in the same period also failed to provide evidence of P. psidii on eucalypt seedlings, cuttings or hedges.
Fragment analyses and genotyping
Isolates used for fragment analyses were collected across three provinces (Gauteng, Limpopo and KwaZulu-Natal) and included those from Backhousia citriodora and Myrtus communis (Gauteng), Eugenia natalitia (Limpopo), Eugenia erythrophylla, H. natalensis and M. communis (KZN). Four samples of P. psidii from Queensland, Australia were included for comparison with the South African material (Table 1). The seven P. psidii samples from South Africa had identical alleles at all seven microsatellite loci. Six of the loci contained two alleles each while locus PpSS195 was homozygous for allele 212. The single multilocus genotype (MLG) found in South Africa was different to that of the single MLG obtained from the four Australian isolates of P. psidii (Table 3).
Inoculation studies
Of the six native South African species of Myrtaceae tested, Eugenia erythrophylla was the most susceptible, and all seedlings of this species showed very high levels of infection. Eugenia verdoorniae was also highly susceptible and all seedlings of this species developed uredinia after inoculation. Some uredinia developed on Syzygium legattii and H. canescens, but these species appeared to have a relatively high level of tolerance to infection. Syzygium cordatum appeared resistant to the South African genotype of P. psidii under glasshouse conditions (Table 4).
Discussion
Puccinia psidii was first discovered in South Africa on a single, non-native ornamental plant in a garden (Roux et al. 2013). Early efforts to find additional sources of infection in that region failed. However, as surveys have expanded and with the help of public awareness campaigns, the rust has been found on numerous host plants and it is clearly widely distributed in South Africa. The results of this study have thus shown, for the first time, that P. psidii occurs under natural conditions on a number of native species of Myrtaceae. While it has also been found on various non-native Myrtaceae in gardens and nurseries, it is interesting that it has yet to be detected on species of Eucalyptus that are widely established in South African plantations.
An important result of this study was that all the isolates of P. psidii from South Africa represented a single genotype of the pathogen based on seven microsatellite markers. The isolates tested included those from both native and non-native plants and collections were as much as 1500 km apart. These results show that there was possibly only one introduction of P. psidii into South Africa, and that the pathogen could have been present in natural environments for much longer than the time when it was first discovered in the country.
The genotype of P. psidii in South Africa was found to be different to the one currently known in Australia. Previous studies have shown that the Australian genotype is identical to that found in Indonesia (McTaggart et al. 2016), New Caledonia, China (Machado et al. 2015) and Hawaii (Machado et al. 2015; Sandhu et al. 2016). Several genotyping studies using a variety of microsatellite markers have traced the global movement of collections of P. psidii (Graça et al. 2011; Machado et al. 2015; Ross-Davis et al. 2013; Zhong et al. 2011). These previous studies have identified a “pandemic” genotype that is clonal, and that has a wide host and geographic range in the Caribbean, Mexico and the USA (Ross-Davis et al. 2013). The “pandemic” genotype is distinct from the MLGs found in Brazil (Graça et al. 2013; Ross-Davis et al. 2013). What is clear at this stage is that the South African genotype is different to the one occurring in Hawaii and Australia (pandemic genotype) and thus, neither of these genotypes has a known source. It is most likely that in both cases the source material for infection would be a plant or plants in South or Central America. Resolution of this important question will require field surveys and additional collections in those areas.
In South America where P. psidii is most likely native, multiple host-associated MLGs have been found (Aparecido et al. 2003; Graça et al. 2013; Machado et al. 2015). In Hawaii and Australia, by contrast, a single genotype that infects multiple hosts occurs (Carnegie and Lidbetter 2012; Graça et al. 2013; Loope 2010; Sandhu et al. 2016; Zhong et al. 2011), indicative of an introduced pathogen. Additionally, the genetic diversity of P. psidii in Brazil is reported to be much higher than that in Australia and Hawaii (Graça et al. 2013; Machado et al. 2015). The same situation is found in South Africa, where multiple native and non-native genera and species of Myrtaceae are affected by a single P. psidii genotype.
Telia were abundant on infected plants in South Africa, both on native and non-native hosts. This is in contrast to reports from other countries where they are reported to be rare (Glen et al. 2007; Pegg et al. 2014b; Pérez et al. 2011). The presence of abundant telia in South Africa could result in recombination and generation of novel genotypes in the country. Telia may also facilitate the survival (overwintering) of the pathogen, allowing for rapid population build-up, after situations not suitable for development of uredinia. Recent studies have shown there are mutations, but no recombination in Australian populations of P. psidii (Machado et al. 2015). Future studies with additional isolates of P. psidii and microsatellite markers, will be required to consider the diversity and whether recombination has occurred in populations of the South African genotype.
Puccinia psidii has a very wide distribution in South Africa including native and non-native Myrtaceae hosts. This extensive distribution suggests that it has been present in the country for substantially longer than the three years since it was first reported in the country. It seems most likely that the pathogen has been distributed widely across South Africa with the nursery trade. Myrtus communis, the host on which it was first detected (Roux et al. 2013), is highly susceptible to infection. Puccinia psidii infection was found in every visited nursery that sold this plant in the greater Tshwane and Ekhuruleni Municipal areas of the Gauteng Province. Some nursery owners reported that M. communis plants had died prior to 2013 and that this had resulted in the plant being abandoned from production programmes. This is consistent with the fact that the trade in ornamental plants and “plants for planting” has been implicated as a major pathway in the spread of plant pathogens globally (Liebhold et al. 2012; Palm and Rossman 2003). This has also resulted in calls for bans in the trade of detrimental ornamental plants and alternative global strategies in a bid to reduce the spread of insect pests and pathogens (Anonymous 2011; Wingfield et al. 2015).
Field surveys and artificial inoculation studies conducted as part of this study have substantially increased the known host range of P. psidii in South Africa. Under natural conditions, in nurseries and in forests, the host range of P. psidii in the country now includes five species of native Myrtaceae and two non-native species. Under greenhouse conditions, and using artificial inoculations, an additional three native Myrtaceae and one non-native (S. jambos) were successfully infected with the South African genotype of P. psidii. Amongst the susceptible native Myrtaceae, two (E. erythrophylla, E. verdoorniae) are near threatened species and one (E. umtamvunensis) is endangered. All three of these Eugenia species are endemic to South Africa and occur in a very small area known as Pondoland in the southern part of the KwaZulu-Natal Province, stretching into the Transkei area of the Eastern-Cape Province. Of those tested under greenhouse conditions, E. erythrophylla was highly susceptible. Equally susceptible was the commonly used indicator plant S. jambos. Although infections occurred on S. cordatum, levels were very low for this wide spread African Myrtaceae species.
The Myrtaceae comprise 17 tribes and an estimated 5500 species (Biffin et al. 2010). The P. psidii genotype discovered in South Africa infects hosts in the Myrteae, Syzygieae, Backhousieae and Psiloxyloideae. This suggests that it may have the potential to infect many other hosts in the sub-family Myrtoideae, including commercially important eucalypts and P. guajava. Morin et al. (2012), conducted extensive host range studies of Australian Myrtaceae and concluded that there is no apparent association between the presence or absence of P. psidii disease symptoms and the phylogenetic relatedness of host taxa. Furthermore, they showed that all Australian Myrtaceae species have the potential to become infected with P. psidii to some degree. The occurrence of resistance (R) genes in eucalypts (Junghans et al. 2003b; Mamani et al. 2010), however, suggests the retention of ancient R-genes (Tobias et al. 2016) and bodes well for breeding programmes seeking to produce disease tolerant plants for commercial purposes. It would serve the commercial plantation industry and guava growers well to screen their breeding populations for resistance to P. psidii and thus to avoid future costly losses due to rust. However, preservation of native species in natural ecosystems that might be threatened by P. psidii would be much more complex. Serious losses to native biodiversity could ermerge as is now being experienced in Australia.
References
Alfenas AC, Zauza EAV, Mafia RG, Assis TF (2004) Clonagem e doenças do eucalipto. Editora UFV, Viçosa, MG, Brazil
Alfenas AC, Zauza EAV, Wingfield MJ, Roux J, Glen M (2005) Heteropyxis natalensis, a new host of Puccinia psidii rust. Australasian. Plant Pathol 34:285–286
Anonymous (2011) The Montesclaros Declaration. End trade in detrimental ornamental to save forests. IUFRO Spotlight #3. http://www.iufro.org/uploads/media/spotlight3-montesclaros-declaration.pdf
Aparecido CC, Figueiredo MB, Furtado EL (2003) Groups of physiological variability in Puccinia psidii populations. Summa Phytopathol 29:234–238
Biffin E, Lucas EJ, Craven LA, Da Costa IR, Harrington MG, Crisp MD (2010) Evolution of exceptional species richness among lineages of fleshy-fruited Myrtaceae. Ann Bot 106:79–93
Carnegie AJ, Lidbetter JR (2012) Rapidly expanding host range for Puccinia psidii sensu lato in Australia. Australas Plant Path 41:13–29
Carnegie AJ, Lidbetter JR, Walker J, Horwood MA, Tesoriero L, Glen M, Priest MJ (2010) Uredo rangelii, a taxon in the guava rust complex, newly recorded on Myrtaceae in Australia. Australas Plant Path 39:463–466
Carnegie AJ, Kathuria A, Pegg GS, Entwistle P, Nagel M, Giblin FR (2016) Impact of the invasive rust Puccinia psidii (myrtle rust) on native Myrtaceae in natural ecosystems in Australia. Biol Invasions 18:127–144. doi:10.1007/s10530-015-0996-y
Castro de HA, Krügner TL, Ideriha CHF, Cappello MSC, Marchi AB (1983) Cross-inoculation of Eucalyptus, guava (Psidum guajava), and rose apple (Syzygium jambos) with Puccinia psidii. Fitopatol Bras 8:491–497
Coates Palgrave C (2002) Keith Coates Palgraves Trees of Southern Africa, New edn. Struik Publishers, Cape Town, South Africa
Coutinho TA, Wingfield MJ, Alfenas AC, Crous PW (1998) Eucalyptus rust: a disease with the potential for serious international implications. Plant Dis 82:819–825
Dayton L, Higgins E (2011). Myrtle rust “biggest threat to ecosystem”. The Australian, April 09, 2011.
Ferreira FA (1983) Eucalyptus rust. Revista Arvore 7:91–109.
Giblin F (2013) Myrtle rust report: New Caledonia. University of the Sunshine Coast, Maroochydore, Queensland, Australia.
Giblin F, Carnegie AJ (2014) Puccinia psidii (Myrtle Rust) – Global host list. Version current at 24 Sept. 2014. https://www.anbg.gov.au/anpc/resources/Myrtle_Rust.html.
Glen M, Alfenas AC, Zauza EAV, Wingfield MJ, Mohammed C (2007) Puccinia psidii: a threat to the Australian environment and economy – a review. Australas Plant Path 36:1–16
Graça RN, Aun CP, Guimarães LMS, Rodriques BVA, Zauza EAV, Alfenas AC (2011) A new race of Puccinia psidii defeats rust resistance in eucalypt. Australas Plant Path 40:442–447
Graça RN, Ross-Davis AL, Klopfenstein NB, Kim MS, Peever TL, Cannon PG, Aun CP, Mizubuti ESG, Alfenas AC (2013) Rust disease of eucalypts, caused by Puccinia psidii, did not originate via host jump from guava in Brazil. Mol Ecol 22:6033–6047
Junghans DT, Alfenas AC, Maffia LA (2003a) Escala de notas para quantificao da ferrugem em Eucalyptus. Fitopatol Bras 28:184–188
Junghans DT, Alfenas AC, Brommonschenkel SH, Oda S, Mello EJ, Grattapaglia D (2003b) Resistance to rust (Puccinia psidii winter) in eucalyptus: mode of inheritance and mapping of a major gene with RAPD markers. Theor Appl Genet 108:175–180
Kawanishi T, Uemastu S, Kakishima M, Kagiwada S, Hamamoto H, Horie H, Namba S (2009) First report of rust disease on ohia and the causal fungus, Puccinia psidii, in Japan. J genet. Plant Pathol 75:428–431
Liebhold AM, Brockerhoff EG, Garrett LJ, Parke JL, Britton KO (2012) Live plant imports: the major pathway for insect and pathogen invasions of the US. Front Ecol Environ 10:135–143. doi:10.1890/110198
Loope L (2010) A summary of information on the rust Puccinia psidii Winter (Guava Rust) with emphasis on means to prevent introduction of additional strains to Hawaii. USGS Survey Open-File Report, United States Department of Interior, pp. 2010–1082
Machado PS, Alfenas AC, Alfenas RF, Mohammed CL, Glen M (2015) Microsatellite analysis indicates that Puccinia psidii in Australia is mutating but not recombining. Australas Plant Path 44:455–462. doi:10.1007/s13313-015-0364-5
MacLachlan JD (1938) A rust of the pimento tree in Jamaica, BWI. Phytopathology 28:157–170
Mamani EMC, Bueno NW, Faria D, Guimarães LMS, Lau D, Alfenas AC, Grattapaglia D (2010) Positioning of the major locus for Puccinia psidii rust resistance (Ppr1) on the Eucalyptus reference map and its validation across unrelated pedigrees. Tree Genet Genomes 6:953–962
Marlatt RB, Kimbrough JW (1979) Puccina psidii on Pimenta dioica in South Florida. Plant Dis Rep 63:510–512
McTaggart AR, Roux J, Granados GM, Gafur A, Tarrigan M, Santhakumar WMJ (2016) Rust (Puccinia psidii) recorded in Indonesia poses a threat to forests and forestry in southeast Asia. Australas Plant Path 45:83–89. doi:10.1007/s13313-015-0386-z
Morin L, Aveyard R, Lidbetter JR, Wilson PG (2012) Investigating the host-range of the rust fungus Puccinia psidii sensu lato across tribes of the family Myrtaceae present in Australia. PLoS One 7(4):e35434
Palm ME, Rossman AY (2003) Invasion pathways of terrestrial plant-inhabiting fungi. In: Ruiz M and Carleton JT (Eds). Invasive species: vectors and management strategies Washington, DC: Island Press.
Pegg GS, Brawner JT, Lee DJ (2014a) Screening Corymbia populations for resistance to Puccinia psidii. Plant Pathol 63:425–436
Pegg GS, Giblin FR, McTaggart AR, Guymer GP, Taylor H, Ireland KB, Shivas RG, Perry S (2014b) Puccinia psidii in Queensland, Australia: disease symptoms, distribution and impact. Plant Pathol 63:1005–1021. doi:10.1111/ppa.12173
Pérez CA, Wingfield MJ, Altier NA, Simento S, Blanchette RA (2011) Puccinia psidii infecting cultivated Eucalyptus and native Myrtaceae in Uruguay. Mycol Prog 10:273–282
Ross-Davis AL, Graça RN, Alfenas AC, Peever TL, Hanna JW, Uchida JY, Hauff RD, Kadooka CH, Kim MS, Cannon PG, Namba S, Minato N, Simento S, Pérez CA, Rayamajhi MB, Morán M, Lodge DJ, Arguedas M, Medel-Ortiz R, López-Ramirez MA, Tennant P, Glen M, Klopfenstein NB (2013) Tracking the distribution of Puccinia psidii genotypes that cause rust disease on diverse Myrtaceous trees and shrubs. In: Chadwick K Comp Proceedings of the 61st Annual Western International Forest Disease Work Conference; October 6–11, Waterton Lakes National Park, Alberta, Canada.
Roux J (2015) Devastating myrtle rust pathogen spreading in South Africa. The Gardener March 2015:44–45
Roux J, Pegg G (2014) South African Myrtales at risk. The imminent threat of a devastating fungal rust pathogen. Veld and Flora March 2014:12–13
Roux J, Greyling I, Coutinho TA, Verleur M, Wingfield MJ (2013) The myrtle rust pathogen, Puccinia psidii, discovered in Africa. IMA Fungus 4:155–159. doi:10.5598/imafungus.2013.04.01.14
Roux J, Germishuizen I, Nadel R, Lee DJ, Wingfield MJ, Pegg GS (2015) Risk assessment for Puccinia psidii becoming established in South Africa. Plant Pathol 64:1326–1335. doi:10.1111/ppa.12380
Sandhu KS, Karaoglu H, Zhang P, Park RF (2016) Simple sequence repeat markers support the presence of a single genotype of Puccinia psidii in Australia. Plant Pathol 65:1084–1094. doi: 10.111/ppa.12501.
Stevens PF (2012) "Angiosperm Phylogeny Website. Version 12." Retrieved July 2014, from http://www.mobot.org/MOBOT/research/APweb/.
Tobias P, Guest D, Külheim C (2016) A curious case of resistance to a new encounter pathogen: Myrtle rust in Australia. Mol Plant Pathol 17:783–788. doi:10.1111/mpp.12331
Tommerup IC, Alfenas AC, Old KM (2003) Guava rust in Brazil – a threat to Eucalyptus and other Myrtaceae. New zeal. J For Sci 33:420–428
Uchida J, Zhong S, Killgore E (2006) First report of a rust disease on Ohia caused by Puccinia psidii in Hawaii. Plant Dis 90
Wilson PG, O’Brien MM, Heslewood MM, Quinn CJ (2005) Relationships within Myrtaceae Sensu Lato based on a matK phylogeny. Plant Syst Evol 251:2–19
Wingfield MJ, Brockerhoff EG, Wingfield BD, Slippers B (2015) Planted forest health: the need for a global strategy. Science 349:832–836. doi:10.1126/science.aac6674
Winter G (1884) Repertorium. Rabenhorstii fungi europaei et extraeuropaei exsiccati cura Dr. G. Winter, Centuria XXXI et XXXII. Hedwigia 23:164–172
Zauza EAV, Lana VM, Maffia LA, Araujo MMFC, Alfernas RF, Silva FF, Alfenas AC (2015) Wind dispersal of Puccinia psidii urediniospores and progress of eucalyptus rust. Forest Pathol 45:102–110
Zhong S, Yang B, Alfenas AC (2008) Development of microsatellite markers for the guava rust fungus, Puccinia psidii. Mol Ecol Resour 8:348–350
Zhong S, Yang B, Puri KD (2011) Characterization of Puccinia psidii isolates in Hawaii using microsatellite DNA markers. J Gen Plant Pathol 77:178–181
Zhuang J-Y, Wei S-X (2011) Additional materials for the rust flora of Hainan Province, China. Mycosystema 30:853–860
Acknowledgments
We are grateful to Dave and Audrey Woods of Indigiflora, Munster, for providing us with plants to use in the screening trials and to Mr. Jason Batemen of the Manie van der Schijff Botanical Gardern (University of Pretoria) for assistance with sourcing plants. Prof. Braam van Wyk (Department of Plant Sciences, University of Pretoria) provided valuable discussions regarding the taxonomy of Myrtaceae and Dr. Geoff Pegg (Department of Agriculture, Fisheries and Forestry, Horticulture and Forestry Science, Agri-Science Queensland, Australia) is thanked for provision of specimens from Australia. This work is based on research partially supported by the Tree Protection Co-operative Program (TPCP) and the National Research Foundation of South Africa (Grant specific unique reference numbers UID 78566 and UID 83924). The grant holders acknowledge that opinions, findings and conclusions or recommendations expressed in any publication generated by the NRF supported research are that of the authors and that the NRF accepts no liability whatsoever in this regard.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Roux, J., Granados, G.M., Shuey, L. et al. A unique genotype of the rust pathogen, Puccinia psidii, on Myrtaceae in South Africa. Australasian Plant Pathol. 45, 645–652 (2016). https://doi.org/10.1007/s13313-016-0447-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13313-016-0447-y