
Abstract
Cold-adapted, complex polysaccharide-degrading marine bacteria have important implications in biogeochemical processes and biotechnological applications. Bacteria capable of degrading complex polysaccharide substrates, mainly starch, have been isolated from various cold environments, such as sea ice, glaciers, subglacial lakes, and marine sediments. However, the total diversity of polysaccharide-degrading culturable bacteria in Kongsfjorden, Arctic Ocean, remains unexplored. In the study reported here, we tested 215 cold-adapted heterotrophic bacterial cultures (incubated at 4 and 20 °C, respectively) isolated from Kongsfjorden, for the production of cold-active extracellular polysaccharide-degrading enzymes, including amylase, pectinase, alginase, xylanase, and carboxymethyl (CM)-cellulase. Our results show that 52 and 41% of the bacterial isolates tested positive for extracellular enzyme activities at 4 and 20 °C, respectively. A large fraction of the bacterial isolates (37% of the positive isolates) showed multiple extracellular enzyme activities. Alginase and pectinase were the most predominantly active enzymes, followed by amylase, xylanase, and CM-cellulase. All isolates which tested positive for extracellular enzyme activities were affiliated to microbial class Gammaproteobacteria. The four genera with the highest number of isolates were Pseudomonas, followed by Psychrobacter, Pseudoalteromonas, and Shewanella. The prevalence of complex polysaccharide-degrading enzymes among the isolates indicates the availability of complex polysaccharide substrates in the Kongsfjorden, likely as a result of glacial melting and/or macroalgal load. In addition, the observed high functional/phenotypic diversity in terms of extracellular enzyme activities within the bacterial genera indicates a role in regulating carbon/carbohydrate turnover in the Kongsfjorden, especially by reducing recalcitrance.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Kongsfjorden is an open glacial fjord located on the west coast of Spitsbergen, an island in the Svalbard archipelago, situated in the Arctic Ocean between mainland Norway and the North Pole (79°N, 12°E). It has been identified as a key European site for Arctic biodiversity monitoring (Hop et al. 2002) and is also suitable for exploring the impacts of possible climate changes (Włodarska-Kowalczuk and Węsławski 2001; Hop et al. 2002). Earlier studies in this fjord have focused on the physical characteristics of the environment (Svendsen et al. 2002), bacterial abundance and biomass (Jankowska et al. 2005), phytoplankton and zooplankton diversity (Hop et al. 2002), benthic microbial community (Srinivas et al. 2009), and marcobenthos (Wlodarska-Kowalczuk and Pearson 2004). However, in the recent past there have been reports on culturable (Srinivas et al. 2009; Choidash et al. 2012; Prasad et al. 2014) and non-culturable (Zeng et al. 2009; Piquet et al. 2010) bacterial diversity of the Kongsfjorden. These studies revealed that exceptionally diverse bacterial communities thrive in the perennially cold (<5 °C) water column of the Kongsfjorden. To date, cultivation-based studies have focused primarily on the bio-prospecting potential of the Kongsfjorden bacterial community with respect to the production of various cold-active enzymes, mostly amylases, lipases, and proteases (Srinivas et al. 2009; Chattopadhyay et al. 2013; Prasad et al. 2014). Despite these findings, however, little is known of the ability of the culturable Kongsfjorden bacterial community to produce extracellular cold-active polysaccharide-degrading enzymes other than amylases.
Kongsfjorden receives 5–10% of its total organic carbon of terrestrial origin from glacier melting, and about 90–95% of the organic carbon in the Kongsfjorden is contributed by primary production (Kuliński et al. 2014). However, Buchholz and Wiencke (2016) reported that macroalgae, in particular kelps, also contribute significantly to the pool of organic matter in the Kongsfjorden. It is important to note that high-molecular-weight carbohydrates or polysaccharides constitute a large proportion of the particulate organic matter (POM) and dissolved organic matter (DOM) in the ocean (Benner et al. 1992; lee et al. 2004). Marine microbial communities possess extracellular enzymes for hydrolyzing these high-molecular-weight substrates to yield sufficiently smaller substrates (<600 Da) for microbial uptake and to make them available for higher trophic levels (Azam et al. 1994; Arnosti 2011). Therefore, there is a strong possibility that complex polysaccharide-degrading cold-adapted bacteria will be present in the waters of Kongsfjorden.
The ability to produce cold-active enzymes is important for the growth and survival of microorganisms living in the permanently cold marine environment. Bacterial activity at low temperature requires higher levels of DOM compared to warmer environments (Pomeroy and Wiebe 2001). In addition, low temperature decreases the rate of chemical reactions and increases protein compactness, thereby affecting enzyme catalysis (Rasmussen et al. 1992). To overcome these challenges, cold-active enzymes have evolved high specific activity and a lower optimum temperature for activity (Feller et al. 1996). Thus, the detection of cold-adapted bacteria with cold-active polysaccharide-degrading enzymes is important for elucidating the mechanism of organic matter transformation in cold marine environments as well as to fulfill the needs of various industries (Georlette et al. 2004; Kumar et al. 2011). The aim of the study reported here was, therefore, to explore both the phylogenetic and functional/phenotypic (in terms of enzyme activity) diversity among polysaccharide-degrading cold-adapted culturable bacteria isolated from the Kongsfjorden in order to decipher their possible role in organic matter transformation.
Material and methods
Study area and sampling
Six different stations located in the Kongsfjorden were sampled, starting at the mouth of Kongsbreen and Kronebreen glaciers and continuing to the open ocean waters (Fig. 1). A Conductivity Temperature Depth (CTD) instrument was deployed at each station to determine depthwise variations in temperature, salinity, and fluorescence. Depthwise distribution of bacteria in Kongsfjorden is affected by the hydrological stratification and/or phytoplankton distribution. Moreover, bacterioplankton and phytoplankton generally tend to accumulate in the layers around the pycnocline in the Kongsfjorden (Jankowska et al. 2005). Therefore, in order to increase the probability of encountering diverse polysaccharide-degrading bacteria we collected 5-l water samples from the deep chlorophyll maxima (DCM) depths using Niskin bottles in June and July 2015. Immediately after collection, the water samples were filtered through a 0.22-μm pore-size polycarbonate membrane (Merck Millipore, Billerica, MA).

Map of Kongsfjorden showing locations of the sampling stations (red stars 1–6)
Isolation of heterotrophic bacteria
The bacterial population retained on the 0.22-μm pore-size polycarbonate membrane was cultivated. Specifically, immediately after filtration of a water sample, the filter paper was immersed in a flask containing 25 ml of Zobell Marine Broth (ZMB; Hi-Media, Mumbai, India) and incubated at 4 °C until visible bacterial growth, identified as increased turbidity, was recorded. At the end of the incubation period, 500 μl of the medium was preserved and transported back to the National Centre for Antarctic and Ocean Research (NCAOR) using standard protocols where individual bacterial cultures were then isolated. For this purpose all 500 μl of the preserved medium was emptied into a flask containing 10 ml of ZMB and incubated at 4 °C for 7 days. At the end of the incubation period 100 μl of the medium was appropriately serially diluted and spread-plated onto Zobell Marine Agar (ZMA). The plates were incubated at 4 °C until visible bacterial colonies formed. Morphologically distinct colonies were handpicked using a sterile loop and transferred onto ZMA slants.
Screening of bacterial isolates for cold-active extracellular polysaccharide-degrading enzyme activities
To study the activities of extracellular polysaccharide-degrading enzymes, such as amylase, pectinase, alginase, xylanase and CM-cellulase, we spot-inoculated bacterial isolates onto modified basal salt medium (MBM) supplemented with 0.5% (w/v) starch, 0.2% (w/v) pectin (poly-D-galacturonic acid methyl ester), 0.2% (w/v) sodium alginate, 0.5% (w/v) xylan from beechwood, and 0.5% (w/v) carboxylmethyl-cellulose (CMC). All polysaccharide substrates were procured from Hi-Media. The MBM contained (per liter) 2 g yeast extract, 0.3 g peptone, 15 g agar, 25 g NaCl, 0.75 g KCl, 7 g MgSO4, 0.5 g NH4Cl, 0.2 g CaCl2, 7 ml 10% w/v K2HPO4, 3 ml 10% w/v KH2PO4, and 1 ml trace metal solution (Bhosle 1981). Prior to sterilization the pH of the MBM was adjusted to 7.2 using 1 N NaOH. After spot-inoculation of the bacterial isolates, the plates were incubated at 4 °C and 20 °C, respectively, for 7 days. The zone of clearance around bacterial colonies due to extracellular polysaccharide-degrading enzyme activities was observed by flooding the plates with appropriate reagents. Xylan- and CMC-containing plates were flooded with 1% Congo Red for 30 min followed by rinsing with 1.5 M NaCl (Khandeparker et al. 2011). Gram’s iodine solution was used for detecting bacterial isolates which could hydrolyze starch, pectin (Sunnotel and Nigam 2002), and alginate (Sawant et al. 2015).
Genomic DNA extraction and PCR assay
Genomic DNA from bacterial cultures which tested positive for enzyme activity was extracted using the Purelink Genomic DNA Extraction kit (Invitrogen, Carlsbad, CA) following the manufacturer’s protocol. The extracted DNA was stored in Purelink elution buffer at −20 °C until analyzed. The 16S rRNA gene from the genomic DNA was amplified using universal bacterial 16S primers 27 F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) (Lane 1991). PCR assays were carried out in 50-μl reaction mixtures containing 20 pmol of each primer, 1.5 mM MgCl2, 200 μM dNTPs, and 2.5 U Taq DNA polymerase. Amplifications were carried out in a thermocycler (model CFX 96; BioRad, Hercules, CA) with the following cycling program: an initial denaturation at 95 °C for 2 min followed by 29 cycles of 30 s at 95 °C, 30 s at 45 °C, and 2 min at 72 °C, with final extension for 10 min at 72 °C. Amplification was confirmed by 1% agarose gel electrophoresis. The amplified products were purified using The Purelink PCR Purification kit (Invitrogen).
Amplified ribosomal DNA restriction analysis and DNA sequencing
The PCR-amplified 16S rRNA gene products were digested separately with the restriction enzymes AluI and HaeIII. The resulting fragments were electrophoresed on 3% polyacrylamide gel, and the gel then stained with ethidium bromide and visualized under UV light. Amplified ribosomal DNA restriction analysis (ARDRA) profiles obtained on different gels were matched using a 100-bp ladder DNA (Invitrogen) as reference. On the basis of the ARDRA profiles, bacterial isolates were clustered into groups, with those isolates sharing similar restriction profiles placed together in a single group, and those isolates with distinct restriction patterns categorized into separate groups. Depending upon the phenotypic pattern (enzyme activities), a variable number of isolates, but always more than one isolate, were selected from each ARDRA group for 16S rRNA gene sequencing. The purified 16S rRNA gene fragments of each representative isolate were sequenced using an automated DNA sequencing system (Applied Biosystems, Foster City, CA).
Sequence analysis
The obtained 16S rRNA gene sequences were checked for the presence of chimera using the DISCPHER software toolset (http://decipher.cee.wisc.edu/; Wright et al. 2012). Non-chimeric sequences were compared with the NCBI database sequences using the MegaBLAST option of the BLAST (https://blast.ncbi.nlm.nih.gov/) algorithm (Altschul et al. 1990). In order to reliably identify the bacterial isolates to the genus level, we also used SeqMatch and classifier tools from Ribosomal Database Project Release 10 (http://rdp.cme.msu.edu/) (Wang et al. 2007). A total of 75 sequences were submitted to GenBank under accession numbers KX417120 to KX417194.
Phylogenetic tree
A phylogenetic tree of the representative sequences (1 sequence per ARDRA group) was constructed to visualize their respective relationship and affiliation with the closest relative type strain sequences from the NCBI database. The tree was constructed in MEGA-5 software using maximum composite likelihood as the substitution model, and bootstrap values were calculated using the neighbor-joining method with a resampling size of n = 500 (Tamura et al. 2011).
Results
Hydrography
Water column temperature decreased from 6.5 °C in the open ocean station (stn1) to 3.0 °C close to the glacier (stn6) (Table 1), and salinity ranged from 29 to 34.2 psu between the inner and outer fjord stations, respectively. The fluorescence signal decreased gradually from 6.7 mg m−3 at stn1 to 0.62 mg m−3 at stn6. In addition, the depth of the DCM decreased gradually from stn 1 (35 m) to stn 6 (5 m) in the Kongsfjorden (Table 1).
Screening for extracellular enzyme activities
A total of 215 bacterial isolates obtained from the six different stations located along the fjord axis were tested for various extracellular enzyme activities using the plate assay method. Of the total number of bacterial isolates assayed, 58% (125 isolates) were found to test positive for polysaccharide-degrading enzymes. At an incubation temperature of 4 °C, 52% (112 isolates) of all bacterial isolates tested positive for extracellular enzyme activities, whereas at 20 °C, 41% (88 isolates) of the total tested positive for extracellular enzyme activities. Among the bacterial isolates isolated, 17.8, 28.7, 41.3, 6.5, and 5.6% were able to hydrolyze starch, pectin, alginate, xylan and CM-cellulose, respectively, when incubated at 4 °C (Fig. 2a); at 20 °C, 22.6, 22.6, 28.8, and 26% of the total isolates were able to hydrolyze starch, pectin, alginate, and xylan, respectively (Fig. 2b). A relatively high percentage of bacterial isolates was able to degrade alginate, pectin, and starch, while the ability to hydrolyze xylan and CM-cellulose was restricted to few isolates. Interestingly, CM-cellulase activity in isolates was recorded exclusively at the 4 °C incubation temperature. These results indicate that the production of alginase and pectinase represent the main enzyme activities of isolates recovered from the Kongsfjorden.

Percentage of bacterial isolates from the total number of isolates that were able to hydrolyze starch, pectin, alginate, xylan, and carboxymethyl (CM)-cellulose at an incubation temperature of 4 °C (a) and 20 °C (b)
Variation in enzyme activities among bacterial isolates according to location
The ability of the bacterial isolated to hydrolyze starch, pectin, alginate, xylan, and CM-cellulose varied with the location of the station (Fig. 3a, b). At the 4 °C incubation temperature, the ability to degrade pectin and alginate was significantly higher among isolates from stn3, stn4, and stn5 (Fig. 3a), while the ability to degrade xylan and CM-cellulose was expressed exclusively in bacterial isolates from stn6 (located near to Kongsbreen glacier; Fig. 3a). At the 20 °C incubation temperature, the ability to degrade xylan was expressed in isolates from stn1, stn4, stn5, and stn6 (Fig. 3b), which otherwise was exclusively observed in isolates from stn6 when incubated at 4 °C.

Location-wise distribution of bacterial isolates degrading a specific substrate at an incubation temperature of 4 °C (a) and 20 °C (b). The percentage of bacterial isolates degrading particular substrate was calculated separately for each station (stn)
Multiple enzyme activities among bacterial isolates
Many bacterial isolates produced more than one extracellular cold-active polysaccharide-degrading enzyme. For example, at the 4 °C incubation temperature, one isolate (identified based on 16S rRNA gene similarity as Pseudomonas extremaustralis) tested positive for all five of the enzymatic activities screened, and two tested positive for four of these enzymatic activities; 36 isolates produced three of the hydrolytic enzymes tested, and 36 tested positive for two of these hydrolytic enzymes. At the 20 °C incubation temperature, none of the isolates tested positive for all five enzymatic activities screened, while nine tested positive for four of these; 21 isolates produced three of the hydrolytic enzymes tested, and eight tested positive for two of these. The percentage of isolates degrading additional substrates within the groups of isolates producing a particular enzymatic activity was calculated according to Tropeano et al. (2012), and the results are shown in Fig. 4a, b. More specifically, all of the isolates that tested positive for amylase were considered to have a value of 100%, and then the percentages of isolates capable of utilizing each one of the other substrates (pectin, alginate, xylan, CM-cellulose) were calculated. This analysis showed that the ability to degrade pectin and alginate was the most frequent enzymatic activity within the groups, followed by the ability to degrade starch, while the ability to degrade xylan and CM-cellulose was the least frequent among the isolated microbial groups. However, at the 4 °C incubation temperature, a large proportion of the isolates producing xylanase were also able to degrade CM-cellulose and vice-versa (Fig. 4a).

Percentage of bacterial isolates able to degrade additional substrates within the groups of isolates producing a particular enzymatic activity at an incubation temperature of 4 °C (a) and 20 °C (b)
Diversity of polysaccharide-degrading bacterial isolates
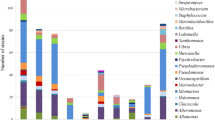
The distribution of the 125 bacterial isolates which tested positive for extracellular enzyme activities among the different genera is shown in Table 2. The ARDRA analysis of these 125 bacterial isolates with two restriction enzymes showed 23 distinct cleavage patterns. A total of 75 representative isolates from all ARDRA groups were selected for 16S RNA sequencing (Table 2), and the results revealed that all of the sequences shared a high similarity to their nearest neighbor type strain sequences deposited in the databases. Most of the closest relative type strains are cold adapted and were isolated from spring and natural mineral water, Antarctic pond water, Antarctic coastal marine environment, diatoms, seaweed, seafood, Arctic sea-ice, and seawater (Table 2). The overall identity of 92% of the bacterial isolates with respect to their closest type strains was >98.8% (Table 2), while approximately 7.6% of the isolates showed a <98.7% similarity to the type strain. Previous studies reported that 98.7% of 16S rRNA gene sequence similarity can be used as the threshold for differentiating two species (Wayne et al. 1987; Stackebrandt and Ebers 2006; Kim et al. 2014). Thus, the isolates in our study most likely represent novel complex polysaccharide-degrading bacterial species. A phylogenetic tree of probable novel bacterial isolates is shown in Fig. 5. It should be noted that multiple ARDRA profiles were observed for the strains belonging to the same species (Table 2), possibly due to intra-species diversity (Chang et al. 2005). Interestingly, high functional/phenotypic diversity in terms of enzyme activities was observed among strains belonging to same genus and/or species (Table 2). Taxonomic affiliation of the representative isolates revealed that all strains belonged to the class Gammaproteobacteria. Four genera were identified, with the highest number of isolates belonging to genus Pseudomonas (50 representative isolates), followed by Psychrobacter (19 isolates), Pseudoalteromonas (8 isolates), and Shewanella (2 isolates). Our results clearly indicate that among all four of these genera, those isolates belonging to Pseudomonas genus were predominant in terms of demonstrating all extracellular enzyme activities tested at 4 °C and 20 °C. A large fraction the isolates in the Kongsfjorden which produce amylase, pectinase, alginase, xylanase and CM-cellulase belong to the genus Pseudomonas.

Phylogenetic tree (maximum likelihood) showing evolutionary relationship of probable novel bacterial isolates with the type strains. Bootstrap values of >50% are given at the nodes (calculated with the neighbor-joining method). Scale bars Percentage of sequence divergence. The isolates are represented by the laboratory reference number along with genbank accession number (in parenthesis)
Discussion
The observed decrease in temperature in the upper layer water column from the outer fjord to the near glacier station indicates cooling of the inner Kongsfjorden. This cooling could be induced by the large influx of cold glacier meltwater into the inner fjord during the summer (Hop et al. 2002). The sharp lateral gradient in salinity decreasing towards near glacier locations indicate that glacier melt induced freshening of the inner fjord. The decrease in magnitude of fluorescence from the outer to inner Kongsfjorden could be attributed to reduced light penetration into the water column as a result of the increased input of turbid freshwater from glacier melting (Svendsen et al. 2002).
Extracellular enzyme activities of cold-adapted bacteria contribute essentially to the processes of nutrient turnover, biomass production, and litter decomposition (Hatha et al. 2013). Thus, the presence of a large number of bacterial isolates capable of producing various cold-active extracellular polysaccharide-degrading enzymes in our samples highlights their importance in carbohydrate/carbon turnover in the Kongsfjorden. In addition, the higher proportion of isolates showing extracellular enzyme activities at the 4 °C incubation temperature compared to the 20 °C incubation temperature suggests that the bacterial community adapted to the permanently cold waters of Kongsfjorden with lower temperature optima of their extracellular enzymes (Huston et al. 2000) and/or temperature-dependent production of extracellular enzymes (Buchon et al. 2000). Bacterial extracellular enzyme activities in most of aquatic environments depend upon the presence or concentration of corresponding substrates (polymers) (Münster et al. 1992). Moreover, it has been widely accepted that hydrolytic enzymes are induced by the presence of polymeric substrates (Chróst 1991; Vetter and Deming 1999; Yu et al. 2009). Thus, the predominance of xylanase and CM-cellulase activities in bacterial isolates from the near glacier location could be due to the presence of lignocellulose-rich organic matter of terrigenous origin, most likely contributed by glacial melting in the summer. Kuliński et al. (2014) reported that glacier melting contributes 5–10% of the total organic carbon of terrestrial origin into the Kongsfjorden. In addition, the dominance of alginate- and pectin-degrading bacteria in the middle and inner fjord locations could be associated with the degradation of macro-algal derived organic matter in the Kongsfjorden. Brown algae are the major source of dissolved and particulate alginate in ocean waters. Hop et al. (2012) reported the occurrence of endemic Arctic brown algae Laminaria solidungula in the middle and inner part of the Kongsfjorden. Recently, Buchholz and Wiencke (2016) estimated that brown alga Alaria esculenta loses its blade gradually at a rate of 3 ± 0.8% of the blade area per day, thereby contributing significantly to the pool of POM in the Kongsfjroden. Consequently, location-wise variation in the ability to degrade polysaccharide substrates among bacterial isolates signifies the presence of a wide variety of organic matter in these waters. It has been reported that the salinity of the sampling location has a negative impact on microbial extracellular enzyme activities (Park et al. 2006; Khandeparker et al. 2011). However, in the present study we did not observed any negative correlation between percentage abundance of polysaccharide-degrading bacteria and the salinity of the sampling location. Nevertheless, Liu and Boone (1991) reported that increased concentrations of NaCl inhibit the growth of lignocellulose-degrading bacterial population. Therefore, it is possible that in this study decreased salinity due to glacial melting might have also contributed to the predominance of xylan- and CM-cellulose degrading bacteria in near glacier location.
The high proportions of isolates capable of producing multiple polysaccharide-degrading enzymes indicate the exceptional adaptation of the Kongsfjorden microbiota to flourish in a cold environment. The ability to produce multiple enzymes provides microorganisms with a higher chance to adapt and survive, which is particularly useful in the rapidly changing marine environment of the Kongsfjorden in terms of uncertainty in the availability of substrates for bacterial metabolism (Tropeano et al. 2012). In addition, the ability of several isolates to produce both xylanase and CM-cellulase has a huge potential for the conversion of lignocellulolytic biomass to bioethanol (Agustini et al. 2012). Interestingly, we observed high phenotypic variation, in terms of enzyme activities, among bacterial isolates belonging to a particular genus and/or species. This phenomenon is known as microbial phenotypic heterogeneity and is a non-genetic variation which does not involve changes in the genome level, but rather is a result of random alterations in the rates of protein synthesis (Veening et al. 2008; Davidson and Surette 2008) modulated by external environmental factors (Rossignol et al. 2009). Phenotypic heterogeneity allows a genetically identical bacterial population to cope with substrate limitations and fluctuations, thus ensuring their survival under highly variable environmental conditions (Ackermann 2015).
Our phylogenetic analyses of the polysaccharide-degrading bacteria obtained in this study reveal that they all belong to the class Gammaproteobacteria. This result is in accordance with earlier reports of a link between carbohydrate degradation in the marine environment and the dominance of members of the class Gammaproteobacteria (Edwards et al. 2010; Gifford et al. 2014). In our study, members of genera Pseudomonas and Psychrobacter were particularly abundant among the isolates which tested positive for the extracellular enzyme activities. This result not only suggests their role in carbohydrate degradation in the cold marine environment but also indicates the bio-prospecting potential of Pseudomonas and Psychrobacter isolates from Kongsfjorden for all of the extracellular enzymatic activities tested. The members of genus Pseudomonas demonstrate a great deal of metabolic diversity and consequently are able to colonize a wide range of niches (Palleroni 1984, 1992). They are found predominantly in the cultured bacteria community from various marine environments due to their ease of culture in vitro (Mancinelli 1984). Members of Pseudomonas and Psychrobacter genera have been reported to produce mainly cold-active lipase (Choo et al. 1998; Rashid et al. 2001; Alquati et al. 2002; Zhang and Zeng 2008; Dang et al. 2009; Dey et al. 2014) and protease (Martınez-Rosales and Castro-Sowinski 2011). In this regard, our results are significant in that we demonstrate the potential of Kongsfjorden bacterial isolates belonging to these genera to produce cold-active complex polysaccharide-degrading enzymes. The closest type strains within the Pseudomonas genus (P. orientalis, P. antarctica, P. grimontii, P. extremaustralis, P. mandellii) were isolated from freshwater environments. Earlier studies from Svalbard, Arctic also reported the dominance of bacterial isolates belonging to Pseudomonas genus, particularly P. orientalis in the glacier ice core (Singh et al. 2015) and glacier meltwater stream (Reddy et al. 2009). Therefore, it appears that increased terrestrial and glacial inputs during the summer might have contributed to the observed dominance of members of Pseudomonas genus in our samples from Kongsfjorden. In addition, the presence of a few isolates in our samples having < 98.7% sequence similarity with the type strain sequence in the database indicates the possibility of having novel and/or unexplored culturable bacterial diversity, which probably contributes to complex carbohydrate degradation in the Kongsfjorden.
In conclusion, our results clearly demonstrate that Kongsfjorden bacterial isolates possess multiple enzyme activities as well as phenotypic heterogeneity for complex polysaccharides degradation, thereby ensuring their survival and growth in an Arctic environment characterized by the variability and availability of different polymeric or complex substrates. Enhanced terrestrial and glacial inputs may also contribute complex carbohydrate-degrading bacteria to the Kongsfjorden. These results have significant implications for understanding carbohydrate/carbon turnover in a glacially influenced, permanently cold marine coastal environment. Further, Kongsfjorden bacterial isolates could be harnessed for the production of cold-active polysaccharide-degrading enzymes for various industrial applications.
References
Ackermann M (2015) A functional perspective on phenotypic heterogeneity in microorganisms. Nat Rev Microbiol 13:497–508
Agustini SL, Efiyanti L, Faulina SA, Santoso E (2012) Isolation and characterization of cellulase- and xylanase producing microbes isolated from tropical forests in Java and Sumatra. Int J Environ Bioener 3(3):154–167
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
Alquati C, De Gioia L, Santarossa G, Alberghina L, Fantucci P, Lotti M (2002) The cold-active lipase of Pseudomonas fragi: heterologous expression, biochemical characterization and molecular modeling. Eur J Biochem 269(13):3321–3328
Arnosti C (2011) Microbial extracellular enzymes and the marine carbon cycle. Annu Rev Mar Sci 3:401–425
Azam F, Steward GF, Smith DC, Ducklow HW (1994) Significance of bacteria in carbon fluxes in Arabian Sea. Proc Indian Acad Sci (Earth Planet Sci) 103(2):341–351
Benner R, Pakulski JD, McCarthy M, Hedges JI, Hatcher PG (1992) Bulk chemical characteristics of dissolved organic matter in the ocean. Science 255(5051):1561–1564
Bhosle NB (1981) Microbial degradation of petroleum hydrocarbons. PhD thesis. University of Bombay, Bombay
Buchon L, Laurent P, Gounot AM, Guespin-Michel JF (2000) Temperature dependence of extracellular enzymes production by psychrotrophic and psychrophilic bacteria. Biotechnol Lett 22(19):1577–1581
Buchholz CM, Wiencke C (2016) Working on a baseline for the Kongsfjorden food web: production and properties of macroalgal particulate organic matter (POM). Polar Biol 39 (11): 2053–2064
Chang HC, Wei YF, Dijkshoorn L, Vaneechoutte M, Tang CT, Chang TC (2005) Species-level identification of isolates of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex by sequence analysis of the 16S-23S rRNA gene spacer region. J Clin Microbiol 43(4):1632–1639
Chattopadhyay AN, Singh P, Dey A, Roy P, Chatterjee S, Saha P, Mukhopadhyay SK (2013) Study of a psychrotolerant amylolytic Paenibacillus sp. isolated from Arctic region. J Microbiol Biotechnol Res 3(4):24–31
Choidash B, Begum Z, Shivaji S (2012) Bacterial diversity of Ny-Alesund, Arctic Archipelago Svalbard. Monogr J Biol Sci 10(1–2):67–72
Choo DW, Kurihara T, Suzuki T, Soda K, Esaki N (1998) A cold-adapted lipase of an Alaskan psychrotroph, Pseudomonas sp. strain B11-1: gene cloning and enzyme purification and characterization. Appl Environ Microbiol 64(2):486–491
Chróst RJ (1991) Environmental control of the synthesis and activity of aquatic microbial ectoenzymes. In: Chróst RJ (ed) Microbial enzymes in aquatic environments. Springer, New York, pp 29–59
Dang H, Zhu H, Wang J, Li T (2009) Extracellular hydrolytic enzyme screening of culturable heterotrophic bacteria from deep-sea sediments of the Southern Okinawa Trough. World J Microbiol Biotechnol 25(1):71–79
Davidson CJ, Surette MG (2008) Individuality in bacteria. Annu Rev Genet 42:253–268
Dey A, Chattopadhyay A, Mukhopadhyay SK, Saha P, Chatterjee S, Maiti TK, Roy P (2014) Production, partial purification and characterization of an extracellular psychrotrophic lipase from Pseudomonas sp. ADT3. J Bioremed Biodegr 5:242
Edwards JL, Smith DL, Connolly J, McDonald JE, Cox MJ, Joint I, Edwards C, McCarthy AJ (2010) Identification of carbohydrate metabolism genes in the metagenome of a marine biofilm community shown to be dominated by Gammaproteobacteria and Bacteroidetes. Genes 1:371–384
Feller G, Narinx E, Arpigny JL, Aittaleb M, Baise E, Genicot S, Gerday C (1996) Enzymes from psychrophilic organisms. FEMS Microbial Rev 18(2–3):189–202
Georlette D, Blaise V, Collins T, D’Amico S, Gratia E, Hoyoux A, Marx JC, Sonan G, Feller G, Gerday C (2004) Some like it cold: biocatalysis at low temperatures. FEMS Microbiol Rev 28(1):25–42
Gifford SM, Sharma S, Moran MA (2014) Linking activity and function to ecosystem dynamics in a coastal bacterioplankton community. Front Microbiol 5(185). doi: 10.3389/fmicb.2014.00185
Hatha AA, Rahiman MK, Krishnan KP, Saramma AV, Saritha G, Deepu L (2013) Characterisation and bioprospecting of cold adapted yeast from water samples of Kongsfjord, Norwegian Arctic. Indian J Geol Mar Sci 42(4):458–465
Hop H, Pearson T, Hegseth EN, Kovacs KM, Wiencke C, Kwasniewski S, Eiane K, Mehlum F, Gulliksen B, Wlodarska-Kowalczuk M, Lydersen C, Weslawski JM, Cochrane S, Gabrielsen GW, Leakey RJG, Lønne OJ, Zajaczkowski M, Falk-Petersen S, Kendall M, Wängberg SÅ, Bischof K, Voronkov AY, Kovaltchouk NA, Wiktor J, Poltermann M, di Prisco G, Papucci C, Gerland S (2002) The marine ecosystem of Kongsfjorden, Svalbard. Polar Res 21:167–208
Hop H, Wiencke C, Vögele B, Kovaltchouk N (2012) Species composition, zonation, and biomass of marine benthic macroalgae in Kongsfjorden, Svalbard. Bot Mar 55:399–414
Huston AL, Krieger-Brockett BB, Deming JW (2000) Remarkably low temperature optima for extracellular enzyme activity from Arctic bacteria and sea ice. Environ Microbiol 2(4):383–388
Jankowska K, Włodarska-Kowalczuk M, Wieczorek P (2005) Abundance and biomass of bacteria in two Arctic glacial fjords. Pol Polar Res 26(1):77–84
Khandeparker R, Verma P, Meena RM, Deobagkar DD (2011) Phylogenetic diversity of carbohydrate degrading culturable bacteria from Mandovi and Zuari estuaries, Goa, west coast of India. Estuar Coast Shelf Sci 95:359–366
Kim M, Oh HS, Park SC, Chun J (2014) Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol 64:346–351
Kuliński K, Kędra M, Legeżyńska J, Głuchowska M, Zaborska A (2014) Particulate organic matter sinks and sources in high Arctic fjord. J Mar Syst 139:27–37
Kumar PS, Ghosh M, Pulicherla KK, Sambasiva Rao KRS (2011) Cold active enzymes from the marine psychrophiles: biotechnological perspective. Adv Biotechnol 10:16–20
Lane DJ (1991) 16S/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M (eds) Nucleic acids techniques in bacterial systematics. John Wiley & Sons, Chichester, pp 115–147
Lee C, Wakeham S, Arnosti C (2004) Particulate organic matter in the sea: the composition conundrum. Ambio 33(8):565–575
Liu Y, Boone DR (1991) Effects of salinity on methanogenic decomposition. Bioresour Technol 35(3):271–273
Mancinelli RL (1984) Population-dynamics of alpine tundra soil bacteria, Niwot Ridge, Colorado Front Range, USA. Arct Alp Res 16(2):185–192
Martınez-Rosales C, Castro-Sowinski S (2011) Antarctic bacterial isolates that produce cold-active extracellular proteases at low temperature but are active and stable at high temperature. Polar Res 30:7123
Münster U, Einiö P, Nurminen J, Overbeck J (1992) Extracellular enzymes in a polyhumic lake: important regulators in detritus processing. Hydrobiologia 229:225–238
Palleroni NJ (1984) Genus I Pseudomonas Migula 1894. In: Krieg NR, Holt JG (eds) Bergey’s manual of systematic bacteriology. Williams and Wilkins, Baltimore, pp 141–199
Palleroni NJ (1992) Introduction to the family pseudomonadaceae. In: Balows A, Truper H, Dworkin M, Harder W, Schleifer KH (eds) The prokaryotes–2. Springer, New York, pp 3071–3079
Park JS, Choi DH, Hwang GJ, Park BC (2006) Seasonal study on ectoenzyme activities, carbohydrate concentrations, prokaryotic abundance and production in a solar saltern in Korea. Aquat Microb Ecol 43(2):153–163
Piquet AMT, Scheepens JF, Bolhuis H, Wiencke C, Buma AGJ (2010) Variability of protistan and bacterial communities in two Arctic fjords (Spitsbergen). Polar Biol 33(11):1521–1536
Pomeroy LR, Wiebe WJ (2001) Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat Microb Ecol 23(2):187–204
Prasad S, Manasa P, Buddhi S, Tirunagari P, Begum Z, Rajan S, Shivaji S (2014) Diversity and bioprospective potential (cold-active enzymes) of cultivable marine bacteria from the Subarctic Glacial Fjord, Kongsfjorden. Curr Microbiol 68(2):233–238
Rashid N, Shimada Y, Ezaki S, Atomi H, Imanaka T (2001) Low-temperature lipase from psychrotrophic Pseudomonas sp. strain KB700A. Appl Environ Microbiol 67(9):4064–4069
Rasmussen BF, Stock AM, Rings D, Petsko GA (1992) Crystallin ribonuclease a loses function below the dynamical transition at 220 K. Nature 357(6377):423–424
Reddy PVV, Rao SSSN, Pratibha MS, Sailaja B, Kavya B, Manorama RR, Singh SM, Srinivas TNR, Shivaji S (2009) Bacterial diversity and bioprospecting for cold-active enzymes from culturable bacteria associated with sediment from a melt water stream of Midtre Lovenbreen glacier, an Arctic glacier. Res Microbiol 160(8):538–546
Rossignol G, Sperandio D, Guerillon J, Duclairoir Poc C, Soum-Soutera E, Orange N, Feuilloley MG, Merieau A (2009) Phenotypic variation in the Pseudomonas fluorescens clinical strain MFN1032. Res Microbiol 160(5):337–344
Sawant SS, Salunke BK, Kim BS (2015) A rapid, sensitive, simple plate assay for detection of microbial alginate lyase activity. Enzym Microb Technol 77:8–13
Singh P, Singh SM, Roy U (2015) Taxonomic characterization and the bio-potential of bacteria isolated from glacier ice cores in the High Arctic. J Basic Microbiol 56(3):275–285
Srinivas TNR, Nageswara Rao SSS, Vishnu Vardhan Reddy P, Pratibha MS, Sailaja B, Kavya B, Hara Kishore K, Begum Z, Singh SM, Shivaji S (2009) Bacterial diversity and bioprospecting for cold-active lipases, amylases and proteases, from culturable bacteria of Kongsfjorden and Ny-Ålesund, Svalbard, Arctic. Curr Microbiol 59(5):537–547
Stackebrandt E, Ebers J (2006) Taxonomic parameters revisited: tarnished gold standards. Microbiol Today 33(4):152–155
Sunnotel O, Nigam P (2002) Pectinolytic activity of bacteria isolated from soil and two fungal strains during submerged fermentation. World J Microbiol Biotechnol 18(9):835–839
Svendsen H, Beszczynska-Mollar A, Hagen JO, Lefauconnier B, Tverberg V, Gerland S, Orbaek JB, Bischof K, Papucci C, Zajaczkowski M, Azzolini R, Bruland O, Wiencke C, Winther JG, Dallmann W (2002) Physical environment of Kongsfjorden, an Arctic fjord system in Svalbard. Pollut Res 21(1):133–166
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28(10):2731–2739
Tropeano M, Coria S, Turjanski A, Cicero D, Bercovich A, Cormack WM, Vazquez S (2012) Culturable heterotrophic bacteria from Potter Cove, Antarctica, and their hydrolytic enzymes production. Pollut Res 31:18507
Veening JW, Smits WK, Kuipers OP (2008) Biostability, epigenetics, and bet-hedging in bacteria. Annu Rev Microbiol 62:193–210
Vetter YA, Deming JW (1999) Growth rates of marine bacterial isolates on particulate organic substrates solubilized by freely released extracellular enzymes. Microb Ecol 37(2):86–94
Wayne LG, Brenner DJ, Colwell RR, Grimont PAD, Kandler O, Krichevsky MI, Moore LH, Moore WEC, Murray RGE, Stackebrandt E, Starr MP, Truper HG (1987) Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int J Syst Bacteriol 37:463–464
Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 73(16):5261–5267
Wlodarska-Kowalczuk M, Pearson T (2004) Soft-bottom macrobenthic faunal associations and factors affecting species distributions in an Arctic glacial fjord (Kongsfjord, Spitsbergen). Polar Biol 27:155–167
Włodarska-Kowalczuk M, Węsławski JM (2001) Impact of climate warming on Arctic benthic biodiversity: a case study of two Arctic glacial bays. Clim Res 18:127–132
Wright ES, Yilmaz LS, Noguera DR (2012) DECIPHER, a search-based approach to chimera identification for 16S rRNA sequences. Appl Environ Microbiol 78(3):717–725
Yu Y, Li H, Zeng Y, Chen B (2009) Extracellular enzymes of cold-adapted bacteria from Arctic sea ice, Canada Basin. Polar Biol 32:1539–1547
Zeng Y, Zheng T, Li H (2009) Community composition of the marine bacterioplankton in Kongsfjorden (Spitsbergen) as revealed by 16S rRNA gene analysis. Polar Biol 32:1447–1460
Zhang JW, Zeng RY (2008) Molecular cloning and expression of a cold adapted lipase gene from an Antarctic deep sea psychrotrophic bacterium Pseudomonas sp. 7323. Mar Biotechnol 10:612–621
Acknowledgments
The authors thank the Director, ESSO-NCAOR, for his support and encouragement. This work was carried out as a part of Indian Scientific Expeditions to the Arctic–2015. We also thank the two anonymous reviewers for their constructive comments, which helped us to improve the manuscript. This is NCAOR contribution number 42/2016.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jain, A., Krishnan, K.P. A glimpse of the diversity of complex polysaccharide-degrading culturable bacteria from Kongsfjorden, Arctic Ocean. Ann Microbiol 67, 203–214 (2017). https://doi.org/10.1007/s13213-016-1252-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13213-016-1252-0