
Abstract
The aim of the present study was to compare the benefit to soybean in terms of phosphate solubilization and host plant growth under salt stress of the endophytic fungi Fusarium verticillioides RK01 and Humicola sp. KNU01. Endophyte F. verticillioides RK01 increased phosphate solubilization significantly more than Humicola sp. KNU01, as confirmed by increasing activities of acid phosphatase (1.0 U−1 g fwt), alkaline phosphatase (2.1 U−1 g fwt ) and fungal biomass (4.6 g), with a concurrent decrease in tri-calcium phosphate concentration in the culture medium. In a plant growth evaluation study, 0.1 M NaCl was added to pots containing soybean plants pre-treated with fungi. The salt-affected plants exhibited stunted growth due to a lower concentration of protein and higher level of lipid peroxidation than their controls. F. verticillioides RK01 and Humicola sp. KNU01 isolates significantly increased shoot length (7.3 % and 2.5 %) and protein content (59.1 % and 65.1 %), while inhibiting the lipid peroxidation level in salt stressed plants. Moreover, carotenoids, the activities of superoxide dismutase (SOD) and catalase were lower in plants affected by salinity than in non-stressed plants, whereas Humicola sp. KNU01 fungal association significantly mitigated oxidative stress by enhancing carotenoids (24.1 %) and SOD activity (61.2 %). F. verticillioides RK01 also triggered SOD activity to prevent oxidative damage. In addition, salt stress caused an increase of absicsic acid (ABA) in soybean plants, and F. verticillioides RK01 colonization effectively reduced ABA content in stressed plants. Salicylic acid (SA) is another plant hormone that was present in lower amounts in stressed plants, but the endophytic (F. verticillioides RK01 and Humicola sp. KNU01) fungal association increased SA content in salt-affected plants. The results of the present study indicate that the efficient phosphate solubilizing endophytic fungi F. verticillioides RK01 was more favorable to protect soybean plants against oxidative damage than Humicola sp. KNU01.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Microorganisms have the ability to solubilize insoluble phosphate (Pradhan and Sukla 2005), and also to produce phytohormones. The inoculation of phosphate solubilizing endophytic fungi into soil is a new approach to increase plant growth even under unfavorable salt stress conditions (Khan et al. 2011a). Phosphorous is present in soil at 20–80 % as inorganic forms with calcium, iron and aluminium; or in organic forms (Richardson 2001) that derive from decaying living organisms (Pandey et al. 2008). Moreover, very low quantities of phosphate (0.1 %) are present in a form accessible to plant uptake (Hariprasad and Niranjana 2009). Phosphate is an important macro nutrient, plays a role in plant bioenergetics, and is needed for converting light energy to ATP during photosynthesis, phosphorylation, cell signaling, and synthesis of biomolecules. Phosphate also leads to stimulation of plant growth, flower and seed formation. The existence of phosphate-solubilizing microorganisms in soil has led to the belief that secretion of microbial metabolites such as organic acids plays a key role in converting insoluble phosphate into soluble forms, and to the suggestion that the application of encapsulated soil microorganisms might enhance phosphate solubilization (Fenice et al. 2000; Vassilev et al. 2001). Moreover, Vassilev et al. (2006) documented that Aspergillus and Penicillium effectively solubilize rock phosphate, and that the fungal association enhances the uptake capacity of phosphorous by plants (Alvarez et al. 2009) and, thus, phosphate-solubilizing microorganisms are helpful for sustainable agriculture (Khan et al. 2007).
The mutual interaction of endophytic fungi and plants may differ in different hosts and environmental conditions (Fateh and Fagan 2002). Root-associated endophytes can improve the growth and yield of crop plants by regulating physiological process such as water- and nutrient- uptake (Auge 2001; Wu et al. 2010). Such endophytes are ubiquitous among the large group of soil microorganisms that are known to exist in saline environments (Giri et al. 2003). Soil salinity is one of the important factors limiting plant growth and yield. Endophytic fungi secrete a wide array of bioactive metabolites including gibberellins and indole acetic acid (Khan et al. 2011a), and the role of these plant growth regulators is well known for growth and development of plants under stress conditions. Poss et al. (1985) documented that fungal association with roots improves the salt tolerance of plants.
Soybean is an economically valuable crop, and the yield of this plant is reduced by high salinity. The exogenous application of various agro chemicals and seed pre-treatments may alleviate the oxidative damage induced by salt stress (Radhakrishnan et al. 2012). Recently, several studies have focused on the use of endophytic fungi to prevent salt stress effect in plants, and results suggest that biological organisms can be used as an alternative to chemical fertilizers (Ghorbanli et al. 2004; Khan et al. 2011a; Sohrabi et al. 2012a; Wu et al. 2013). A number of beneficial microorganisms have been isolated from soil and plant roots, and their plant growth promoting ability has also been well demonstrated in crop plants, but still very few studies have documented or elucidated the role in plants of endophytic fungi in stress regulatory metabolism against abiotic stress (Khan et al. 2012). In the present study, we investigated the mitigating effect of endophytic fungi (F. verticillioides RK01 and Humicola sp. KNU01) on oxidative stress-related physiological changes such as plant hormones and antioxidant activities in stressed plants. The aim of the current study was to evaluate the phosphate solubilization ability of two endophytic fungi, and their mitigating effects on soybean plants under salt stress conditions.
Materials and methods
Isolation and identification of endophytes from soybean roots
The endophytic fungi were isolated from roots of soybean plants grown in a greenhouse according to the method described by Khan et al. (2011a). The roots were separated from plants and washed thoroughly with tap water followed by Tween 80 solution, 1 % perchloric acid and sterilized distilled water. The root samples (0.5 cm) were placed on Hagem medium plates (0.5 % glucose, 0.005 % KH2PO4, 0.05 % MgSO4 · 7H2O, 0.05 % NH4Cl, 0.1 % FeCl3, 80 ppm streptomycin and 1.5 % agar; pH 5.6 ± 0.2), and incubated at 25 °C. The fungal colonies were separated and re-inoculated on potato dextrose agar plates. The two fungal colonies were cultured individually on Czapek broth (1 % glucose, 1 % peptone, 0.05 % KCl, 0.05 % MgSO4 · 7H2O, and 0.001 % FeSO4 · 7H2O; pH 7.3 ± 0.2) for 7 days at 30 °C with 120 rpm. For identification of fungi, genomic DNA was isolated from each isolate as described by Khan et al. (2011a), and identified by sequencing the internal transcribed spacer (ITS) rDNA, using universal primers ITS-1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS-4 (5′-TCCTCCGCTTATTGATATGC-3′). The BLAST search program (http://www.ncbi.nlm.nish.gov/BLAST) was used to compare the nucleotide sequence homology of the 18S ITS region of fungi. The two fungal isolates were identified as Fusarium verticillioides isolate RK01 (accession number KC684889) and Humicola sp. KNU01 (accession number KC797230).
Phosphate solubilization ability of fungi
The two fungal strains (F. verticillioides RK01 and Humicola sp. KNU01) were tested for their capacity to solubilize tricalcium phosphate. For estimation of phosphate solubilization activity, 100 mL medium (PD broth, adjusted to pH 7.0) containing 0.5 % of tricalcium phosphate (w/v) was inoculated with 100 μL fungal sample (ca. 30x103 CFU/mL) and incubated at 28 °C for 30 days (Kang et al. 2002). The tricalcium phosphate and pH in culture filtrates, as well as fungal biomass and acid phosphatase and alkaline phosphatase activities were recorded at 10- and 30-day intervals. Briefly, fungal culture was centrifuged at 7,000 rpm for 20 min after measuring the pH, and tricalcium phosphate was estimated in the culture supernatant at 600 nm in a spectrophotometer according to King (1936) with some modifications. The pellet of fungal mycelial samples was transferred to a pre-weighed Whatman No.1 filter paper, and biomass was measured. For the analysis of phosphates activity, fungi were ground separately with 0.1 M acetate buffer (pH 5.0) for acid phosphatase or with 0.1 M Tris-HCl buffer (pH 8.2) for alkaline phosphatase, as a source of crude enzyme. Activity of acid and alkaline phosphatase was measured according to the modified method of Ikawa et al. (1964).
Fungal interaction with soybean plants under salt stress
Two-week-old fungal cultures of F. verticillioides RK01 and Humicola sp. KNU01 were applied to 15-day-old soybean seedlings grown on substrate containing peat moss (13–18 %, w/v), perlite (7–11 %, w/v), coco-peat (63–68 %, w/v) and zeolite (6–8 %, w/v) and ~90 mg/kg NH4 +, ~205 mg/kg NO3 −, ~350 mg/kg P2O5, and ~100 mg/kg K2O. Pots treated with sterile water served as a control. The plants were grown in a greenhouse under a day/night cycle of 16/8 h and at 28/20 °C and 60 % relative humidity. For salt stress induction, 300 mL 0.1 M NaCl was applied to soybean plants at 33 days. Shoot length and biochemical content were analyzed in 38-day-old soybean plants. Six replicates were used in each treatment.
Oxidative damage to lipids
Lipid peroxidation of leaf samples was estimated in terms of amount of malondialdehyde (MDA) content according to the method of Ohkawa et al. (1979). The reaction mixture consisted of 0.2 mL 10 mM phosphate buffer (pH 7.0), leaf extract, 0.2 mL 8.1 % sodium dodecyl sulphate, 1.5 mL 20 % acetic acid (pH 3.5) and 1.5 mL 0.81 % thiobarbituric acid aqueous solution. The mixture was heated in boiling water for 60 min, and 5 mL butanol:pyridine (15:1 v/v) solution was added at room temperature. The absorbance of the upper organic layer was read at 532 nm. Tetramethoxypropane was used as an external standard. The concentration of MDA was calculated from standard graph values and expressed as milligrams of MDA formed per gram of fresh weight.
Protein content
Protein content in leaf samples was determined according to the method of Bradford (1976). Leaf samples were homogenized in 50 mM Tris-HCl buffer (pH 7.0), 3 mM MgCl2, 1 mM EDTA and 1.0 % PVP and centrifuged at 10,000 rpm for 15 min. The protein content of supernatant (or crude enzyme extract) was measured at 595 nm using a spectrophotometer.
Catalase activity
Catalase (CAT) activity was assayed by the method of Aebi (1984); crude enzyme extract was added to 0.5 mL 0.2 M H2O2 in 10 mM phosphate buffer (pH 7.0). CAT activity was estimated by the decrease in absorbance of H2O2 at 240 nm (1 unit of CAT activity was defined as that amount of the enzyme that breaks down 1 μmol H2O2 per minute under assay conditions).
Superoxide dismutase activity
Superoxide dismutase (SOD) activity was estimated by measuring the degree of inhibition of pyrogallol auto-oxidation by crude enzyme extract, according to Marklund and Marklund (1974). The reaction mixture for auto-oxidation consisted of 2 mL Tris–HCl buffer (pH 8.2), 0.5 mL 2 mM pyrogallol and 2 mL distilled water. Initially, the rate of auto-oxidation of pyrogallol was noted every minute for 3 min. This was considered as 100 % auto oxidation. The assay mixture for the enzyme contained 2 mL Tris-HCl buffer pH 8.2, 1.5 mL distilled water, 0.5 mL supernatant and 0.5 mL 2 mM pyrogallol. The absorbance was read at 470 nm. The enzyme activity was expressed in units. One unit of SOD activity was the amount of enzyme needed to inhibit 50 % of auto-oxidation of pyrogallol per minute.
Abscisic acid content
The endogenous abscisic acid (ABA) content was extracted from leaf tissue and estimated according to the method of Qi et al. (1998). The extracts were dried and methylated by adding diazomethane for GC-MS SIM (6890 N network GC system, and 5973 network mass selective detector; Agilent Technologies, Palo Alto, CA) analysis. For quantification of ABA, Lab-Base data system software (ThermoQuset, Manchester, UK) was used to monitor responses to ions of m/e 162 and 190 for Me-ABA and 166 and 194 for Me-[2H6]-ABA.
Salicylic acid content
Salicylic acid (SA) was extracted and quantified by method of Enyedi et al. (1992) and Seskar et al. (1998). Freeze dried plant samples were homogenized with 90 % and 100 % methanol by centrifuging at 10,000 rpm. The combined methanol extracts were vacuum-dried. Dry pellets were re-suspended in 2.5 mL 5 % trichloroacetic acid, and the supernatant was partitioned with ethyl acetate: cyclopentane: isopropanol (100:99:1, v/v/v). The top organic layer containing free SA was dried with nitrogen gas and again suspended in 1 mL 70 % methanol. The samples were injected to C18 reverse-phase high performance liquid chromatography (HPLC) column (HP hypersil ODS, particle size 5 μmol, pore size 120 Å). SA was identified by fluorescence detector (Shimadzu RF-10AXL, excitation and emission at 305–365 nm, respectively). The flow rate was 1.0 mL/min.
Statistical analysis
The experiments were performed in a completely randomized design. Data were subjected to statistical analysis using SPSS software version 11.5 (SPSS, Cary, NC). The significant differences among the mean values of treatments (n = 3) were determined using DMRT test at P < 0.05 level.
Results and discussion
Soil contains a wide range of microbes including fungi; beneficial mycorrhizal fungi, which live in either soil or plant roots as endophytes, play a vital role in improving plant growth. In the current study, two endophytic fungi were isolated from soybean roots and identified as F. verticillioides RK01 and Humicola sp. KNU01 based on their ITS region and phyllogenetic analysis. The phosphate solubilization ability of these endophytic fungi was assessed at 10 and 30 days, and corresponding changes in biomass of fungi, pH and phosphate concentration of broth culture were determined (Table 1). F. verticillioides RK01 isolate exhibited a higher amount of biomass (7.19 g) than Humicola sp. KNU01 isolate. A reduction in pH was observed in 10- and 30-day-old fungal cultures (F. verticillioides RK01 and Humicola sp. KNU01) compared to the initial pH of the medium (pH 7.0). This indicates an increase in gluconic acid production and phosphate solubilization in the medium (Reyes et al. 1999). The higher level of phosphate solubilization ability of the F. verticillioides RK01 isolate was evidenced from the low concentration of tri-calcium phosphate (0.26 mg and 0.001 mg at 10 and 30 days, respectively) in the medium. Secretion of phosphatase enzymes, secondary metabolites and organic acids from fungi is a common mechanism, which induces the conversion of insoluble forms of phosphorous to plant-available forms (Jha et al. 2009). Acid and alkaline phosphatase play significant roles in fungi in the utilization of organic phosphate from the soil. Both acid and alkaline phosphatase activities were higher in endophytic fungus F. verticillioides RK01 than in Humicola sp. KNU01 (Table 1).
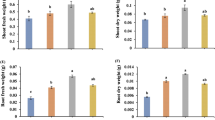
The mutualistic interaction of endophytic fungi favors improvement in the plant growth and development, and can enhance stress tolerance of plants grown under unfavorable abiotic stress environments. Soil salinity limits plant growth, but the host plant is protected by endophytes and their biologically active secondary metabolites (Khan et al. 2011b). Previously, the ameliorative role of mycorrhizal fungi against salt stress in lettuce, citrus, tomato and bajra was documented by Kohler et al. (2009), Wu et al. (2010), Hajiboland et al. (2010) and Borde et al. (2011). In the present study, salt-stress-affected soybean plants showed a significant reduction in their shoot length (7.9 %); this reduction in growth was ameliorated by F. verticillioides RK01 and Humicola sp. KNU01, which increased shoot length by 7.3 % and 2.5 %, respectively (Fig. 1). A remarkable level of stress alleviation was found in F. verticillioides RK01-associated plants. Yates et al. (2005) reported that pretreatment of seeds with F. verticillioides enhanced maize plant growth and yield, under both biotic and abiotic environmental conditions. The important contribution of F. verticillioides RK01 fungal association to the improvement of plant growth might be due to an increase in uptake of phosphorus and other elements by host plants (Wu et al. 2010). Previous findings also revealed that Glomus sp., Diversispora sp. and Exophiala sp. fungal interaction could improve host plant growth under abiotic stresses (Khan et al. 2011c; Sohrabi et al. 2012a; Sheng et al. 2009), suggesting that inoculation of plants with mycorrhizae can enhance plant growth by increasing the water- and nutrient- absorption capacity of plants (Auge 2001). In addition, salt stress significantly diminished the protein content (82.9 %) in non-fungal associated plants, while endophytic fungal (F. verticillioides RK01 and Humicola sp. KNU01)-associated plants exhibited a higher protein content under stress conditions (Fig. 1). Sohrabi et al. (2012b) also observed a higher level of protein in drought-stressed chickpea plants inoculated with Glomus sp. than in stressed plants.

Influence of Fusarium verticillioides RK01 (Fus) and Humicola sp. KNU01 (Hum) on soybean shoot length and protein content in NaCl salt (S)-injured plants. Bars Means plus standard error. Means followed by the same letter are not significantly different (P < 0.05) as determined by DMRT test
Accumulation of ROS in plant cells under environmental stress conditions is a major factor causing damage to lipids, proteins and nucleic acids (Imlay and Linn 1988; Jiang and Zhang 2001). Salt stress causes detrimental effects on plants by increasing lipid peroxidation (MDA content) and reducing antioxidant activities (Fig. 2). High concentrations of MDA in plant cells are considered a stress indicator, damaging membrane lipids and leading to inactivation of receptors, enzymes and ion channels (Gill and Tuteja 2010; Radhakrishnan et al. 2012). Exogenous application of endophytic fungi, F. verticillioides RK01 and Humicola sp. KNU01 reduced the MDA level in salt-stressed plants (Fig. 2), and a significant three-fold reduction of MDA content (0.2 mg/g fwt) was observed in Humicola sp. KNU01 fungi-associated salt-affected plants. These results suggest that endophytic fungi protects host plants by avoiding soil-salinity-induced oxidative stress damage.

Effect of F. verticillioides RK01 (Fus) and Humicola sp. KNU01 (Hum) on lipid peroxidation, carotenoids, superoxide dismutase (SOD) and catalase (CAT) activities in plants exposed to NaCl salt (S) stress. Bars Means plus standard error (n = 3). Means followed by the same letter are not significantly different (P < 0.05) as determined by DMRT test
Next, the mitigating effect of F. verticillioides RK01 and Humicola sp. KNU01 against salinity stress was further assessed by measuring antioxidant activities. It is widely known that ROS metabolism is correlated with antioxidant molecules and enzymes (Radhakrishnan and Lee 2013); however, there is little information on the protective effect of mycorrhizal colonization on plant enzymatic antioxidant defense systems (Ghorbanli et al. 2004). Plants are able to prevent the cytotoxic effects of ROS by using antioxidants such as SOD, CAT and carotenoids (Sharma and Dubey 2005). In the current study, we found a reduction (19.2 %) in the level of antioxidants such as carotenoids, and in the activities of CAT (58.1 %) and SOD (23.7 %) in salt-stressed plants compared to controls (Fig. 2). Under stress conditions, superoxide anion (O2 −) and hydrogen peroxide (H2O2) production are higher in plants, and self defense mechanisms such as SOD and CAT are activated to prevent oxidative damage (Radhakrishnan and Lee 2013). SOD converts O2 − content to H2O2 and water, whereas CAT dismutates H2O2 to water and oxygen. A significant higher SOD activity was found in F. verticillioides RK01- and Humicola sp. KNU01-inoculated plants than in fungal free control plants even under stress conditions (Fig. 2). These results reflect the fact that endophytic fungal association avoids stress effects by stimulating SOD activity to scavenge ROS. However, F. verticillioides RK01 and Humicola sp. KNU01 fungi inhibited CAT activity in salt-stressed plants. The mechanisms of antioxidant systems in plant cells are complex, and fluctuate under an array of stresses. On the other hand, carotenoids are pigment molecules involved in photosynthesis that also act as antioxidants in plants to prevent the photo-oxidative damage generated by biotic and abiotic factors (Siefermann-Harms 1987). We found that Humicola sp. KNU01 fungal association increased the level of carotenoids in salt-stressed plants (Fig. 2), while F. verticillioides RK01 fungal interactions with soybean under salt-stressed conditions showed a lower concentration of carotenoids than stressed plants. Therefore, the elevated content of carotenoids and SOD activity in Humicola sp. KNU01-associated plants might indicate that this fungus had an efficient capacity to avoid oxidative stress.
The response of plants to salt stress is initiated at the receptors in membranes, and results in the production of ROS, ABA and other secondary signaling molecules (Porcel et al. 2012). ABA protects plants from abiotic stresses through the regulation of many stress-responsive genes and stomatal closure to minimize water loss (Zhang et al. 2006). It has been widely documented that accumulation of ABA content in plants increases under salt stress conditions, while fungal association with plants can regulate ABA synthesis (Jahromi et al. 2008; Khan et al. 2012; Porcel et al. 2012). The findings of the current study showed higher accumulation of ABA in plants exposed to salt stress than in controls (Fig. 3). The F. verticillioides RK01 isolate ameliorated the stress effect in soybean by reducing ABA content, while Humicola sp. KNU01 fungal association had no effect on ABA content in plants under salt stress. Khan et al. (2011b) reported a lower concentration of ABA in salt-injured soybean plants colonized by Penicillium minioluteum than in non-fungal treated plants. There is also evidence that levels of ABA altered by fungal association induces salt stress tolerance in soybean. However, the effect of mycorrhizae on endogenous ABA content in plants seems contradictory (Dodd and Perez-Alfocea 2012), and Evelin et al. (2009) suggested that ABA accumulation might respond differentially to the influence of different mycorrhizal fungal species. In our study, we also found that Humicola sp. KNU01 fungal association had no effect on ABA content.

Fusarium verticillioides RK01 (Fus)- and Humicola sp. KNU01 (Hum)-induced changes in abscisic acid (ABA) and salicylic acid (SA) content in soybean plants under NaCl salt (S) stress conditions. Bars Means plus standard error (n = 3). Means followed by the same letter are not significantly different (P < 0.05) as determined by DMRT test
SA is a signaling molecule that plays an important role in plant response to abiotic stresses (Alonso-Ramirez et al. 2009) and that is necessary to generate antioxidants (Sharma et al. 1996). The mutual interaction of fungi and plants increases SA production in induced systemic resistance against abiotic stresses, and high endogenous SA can mitigate the negative effects of ROS accumulation (Khan et al. 2012). Although some studies have documented fungal-induced SA accumulation under abiotic stresses, we compared SA accumulation in soybean upon two endophytic fungal interactions with host plants grown under saline conditions (Fig. 3). Plants exposed to salt stress showed a significantly lower concentration of SA (1.10 μg/g fwt) than controls (2.87 μg/g fwt), but inoculation with endophytic fungi increased the SA level. The increase in SA content varied with F. verticillioides RK01 and Humicola sp. KNU01 treatments. Humicola sp. KNU01 fungi completely reversed the inhibition of SA level decrease in salt-stress-affected plants, while F. verticillioides RK01-associated plants showed a partially rescue of SA reduction induced by salinity. Khan et al. (2011b) also found that the endophytic fungus Penicillium minioluteum had the ability to enhance plant growth under salt stress conditions via increasing SA biosynthesis.
In conclusion, F. verticillioides RK01 and Humicola sp. KNU01 fungi are involved in effective stress avoidance in soybean plants grown in saline soils, as conformed by an increase of shoot length, protein, SOD activity and SA content, and a decrease in lipid peroxidation. Among these two fungal enophytes, F. verticillioides RK01 had a greater phosphate solubilization capacity than the Humicola sp. KNU01 isolate. The results of plant growth under stress conditions, together with the phosphate solubilization ability of F. verticillioides RK01, suggest that exogenous application of F. verticillioides RK01 was more favorable in enhancing soybean plant growth in saline environments compared to Humicola sp. KNU01 application.
References
Aebi H (1983) Catalase. In: Bergmeyer HU (ed) Methods of enzymatic analysis. 3rd edn. Verlag Chemie, Weinheim, pp 273–286
Alonso-Ramirez A, Rodriguez D, Reyes D, Jimenez JA, Nicolas G, Lopez-Climent M, Gomez-Cadenas A, Nicolas C (2009) Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in Arabidopsis seeds. Plant Physiol 150:1335–1344
Alvarez M, Huygens D, Olivares E, Saavedra I, Alberdi M, Valenzuela E (2009) Ectomycorrhizal fungi enhance nitrogen and phosphorus nutrition of Nothofagus dombeyi under drought conditions by regulating assimilative enzyme activities. Physiol Plant 136(4):426–436
Auge RM (2001) Water relations, drought and vesicular arbuscular mycorrhizal symbiosis. Mycorrhiza 11:3–42
Borde M, Dudhane M, Jite P (2011) Growth photosynthetic activity and antioxidant responses of mycorrhizal and non-mycorrhizal bajra (Pennisetum glaucum) crop under salinity stress condition. Crop Prot 30:265–271
Bradford MM (1976) Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254
Dodd IC, Perez-Alfocea F (2012) Microbial amelioration of crop salinity stress. J Exp Bot 63:3415–3428
Enyedi AJ, Yalpani N, Silverman P, Raskin I (1992)Localization, conjugation, and function of salicylic acid in tobacco during the hypersensitive reaction to tobacco mosaic virus.Proc Natl Acad Sci USA 89:2480–2484
Evelin H, Kapoor R, Giri B (2009) Arbuscular mycorrhizal fungi in alleviation of salt stress: a review. Ann Bot 104:1263–1280
Fateh SH, Fagan WF (2002) Fungal endophytes: common host plant symbionts but uncommon mutualists. Integ Comp Biol 42:360–368
Fenice M, Selbman L, Federici F, Vassilev N (2000) Application of encapsulated Penicillium variabile P16 in solubilization of rock phosphate. Bioresour Technol 73:157–162
Ghorbanli M, Ebrahimzadeh H, Sharifi M (2004) Effects of NaCl and mycorrhizal fungi on antioxidative enzymes in soybean. Biol Plant 48:575–581
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Giri B, Kapoor R, Mukerji KG (2003) Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass and mineral nutrition of Acacia auriculiformis. Biol Fertil Soils 38:170–175
Hajiboland R, Aliasgharzadeh N, Laiegh SF, Poschenrieder C (2010) Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 331:313–327
Hariprasad P, Niranjana SR (2009) Isolation and characterization of phosphate solubilizing rhizobacteria to improve plant health of tomato. Plant Soil 316:13–24
Ikawa T, Nisizawa K, Miwa T (1964) Specificities of several acid phosphatases from plant sources. Nature 203:939–940
Imlay JA, Linn S (1988) DNA damage and oxygen radical toxicity. Science 240:1302–1309
Jahromi F, Aroca R, Porcel R, Ruiz-Lozano JM (2008) Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular esponses of mycorrhizal lettuce plants. Microb Ecol 55:45–53
Jha BK, Pragash MG, Cletus J, Raman G, Sakthivel N (2009) Simultaneous phosphate solubilization potential and antifungal activity of new fluorescent pseudomonad strains, Pseudomonas aeruginosa, P. plecoglossicida and P. mosselii. World J Microbiol Biotechnol 25:573–581
Jiang M, Zhang J (2001) Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Pysiol 42:1265–1273
Kang SC, Hat CG, Lee TG, Maheshwari DK (2002) Solubilization of insoluble inorganic phosphates by a soil-inhabiting fungus Fomitopsis sp. PS 102. Curr Sci 82:439–442
Khan AL, Hamayun M, Kim YH, Kang SM, Lee IJ (2011a) Ameliorative symbiosis of endophyte (Penicillium funiculosum LHL06) under salt stress elevated plant growth of Glycine max L. Plant Physiol Biochem 49:852–861
Khan AL, Hamayun M, Ahmad N, Hussain J, Kang SM, Kim YH, Adnan M, Tang DS, Waqas M, Radhakrishnan R, Hwang YH, Lee IJ (2011b) Salinity stress resistance offered by endophytic fungal interaction between Penicillium minioluteum LHL09 and Glycine max L. J Microbiol Biotechnol 21:893–902
Khan AL, Hamayun M, Ahmad N, Waqas M, Kang SM, Kim YH, Lee IJ (2011c) Exophiala sp. LHL08 reprograms Cucumis sativus to higher growth under abiotic stresses. Physiol Plant 143:329–343
Khan AL, Hamayun M, Waqas M, Kang SM, Kim YH, Kim DH, Lee IJ (2012) Exophiala sp.LHL08 association gives heat stress tolerance by avoiding oxidative damage to cucumber plants. Biol Fertil Soils 48:519–529
Khan MS, Zaidi A, Wani PA (2007) Role of phosphate-solubilizing microorganisms in sustainable agriculture—a review. Agron Sustain Dev 27:29–43
King JE (1936) The colorimetric determination of phosphorus. Biochem J 26:292–297
Kohler J, Hernandez JA, Caravaca F, Roldan A (2009) Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ Exp Bot 65:245–252
Marklund S, Marklund G (1974) Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem 47:469–474
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxidation in animal tissues by thiobarbituric acid reaction. Ann Biochem 95:351–358
Pandey A, Das N, Kumar B, Rinu K, Trivedi P (2008) Phosphate solubilization by Penicillium spp. isolated from soil samples of Indian Himalayan region. World J Microbiol Biotechnol 24:97–102
Pradhan N, Sukla LB (2005) Solubilization of inorganic phosphates by fungi isolated from agriculture soil. Afr J Biotechnol 5(10):850–854
Porcel R, Aroca R, Ruiz-Lozano JM (2012) Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron Sustain Dev 32:181–200
Poss JA, Pond E, Menge JA, Harrell WM (1985) Effect of salinity on mycorrhizal onion and tomato in soil with and without additional phosphate. Plant Soil 88:307–319
Qi Q, Rose PA, Abrams GD, Taylor DC, Abrams SR, Cutler AJ (1998) (+)-Abscisic acid metabolism, 3-ketoacyl-coenzyme A synthase gene expression, and very long-chain monounstaurated fatty acid biosynthesis in Brassica napus embryos. Plant Physiol 117:979–987
Radhakrishnan R, Leelapriya T, Ranjithakumari BD (2012) Effects of pulsed magnetic field treatment of soybean seeds on calli growth, cell damage, and biochemical changes under salt stress. Bioelectromagnetics 33:670–681
Radhakrishnan R, Lee IJ (2013) Spermine promotes acclimation to osmotic stress by modifying antioxidant, abscisic acid, and jasmonic acid signals in soybean. J Plant Growth Regul 32:22–30
Reyes I, Bernier L, Simard RR, Tanguay P, Antoun H (1999) Characteristics of phosphate solubilization by an isolate of a tropical Penicillium rugulosum and two UV-induced mutants. FEMS Microb Ecol 28:291–295
Richardson AE (2001) Prospects for using soil microorganism to improve the acquisition of phosphate by plant. Aust J Plant Physiol 28:897–906
Seskar M, Shulaev V, Raskin I (1998)Endogenous methyl salicylate in pathogen-inoculated tobacco plants.Plant Physiol 116:387–392
Sharma P, Dubey RS (2005) Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul 46:209–221
Sharma YK, Leon J, Raskin I, Davis KR (1996) Ozoneinduced responses in Arabidopsis thaliana: the role of salicylic acid in the accumulation of defense-related transcripts and induced resistance. Proc Natl Acad Sci USA 93:5099–5104
Sheng M, Tang M, Chen H, Yang B, Zhang F, Huang Y (2009) Influence of arbuscular mycorrhizae on the root system of maize plants under salt stress. Can J Microbiol 55:879–886
Siefermann-Harms D (1987) The light-harvesting and protective functions of carotenoids in photosynthetic membranes. Physiol Plant 69:561–568
Sohrabi Y, Heidari G, Weisany W, Ghasemi-Golezani K, Mohammadi K (2012a) Changes of antioxidative enzymes, lipid peroxidation and chlorophyll content in chickpea types colonized by different Glomus species under drought stress. Symbiosis 56:5–18
Sohrabi Y, Heidari G, Weisany W, Ghasemi-Golezani K, Mohammadi K (2012b) Some physiological responses of chickpea cultivars to arbuscular mycorrhiza under drought stress. Russ J Plant Physiol 59:708–716
Vassilev N, Vassileva M, Fenice M, Federici F (2001) Immobilized cell technology applied in solubilization of insoluble inorganic (rock) phosphates and P plant acquisition. Bioresour Technol 79:263–271
Vassilev N, Vassileva M, Nikolaeva I (2006) Simultaneous P-solubilizing and biocontrol activity of microorganisms: potentials and future trends. Appl Microbiol Biotechnol 71:137–144
Wu QS, Zon YN, Liu W, Ye XE, Zai HE, Zhao LJ (2010) Alleviation of salt stress in citrus seedlings inoculated with mycorrhiza: changes in leaf antioxidant defense systems. Plant Soil Environ 56:470–475
Wu QS, Zou YN, Huang YM (2013) The arbuscular mycorrhizal fungus Diversispora spurca ameliorates effects of waterlogging on growth, root system architecture and antioxidant enzyme activities of citrus seedlings. Fungal Ecol 6:37–43
Yates IE, Widstrom NW, Bacon CW, Glenn A, Hinton DM, Sparks D, Jaworski AJ (2005) Field performance of maize grown from Fusarium verticillioides-inoculated seed. Mycopathologia 159:65–73
Zhang J, Jia W, Yang J, Ismail AM (2006) Role of ABA in integrating plant responses to drought and salt stresses. Field Crop Res 97:111–119
Acknowledgment
This research work was supported financially by the Eco-Innovation Project, Korea Ministry of the Environment.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Radhakrishnan, R., Khan, A.L., Kang, S.M. et al. A comparative study of phosphate solubilization and the host plant growth promotion ability of Fusarium verticillioides RK01 and Humicola sp. KNU01 under salt stress. Ann Microbiol 65, 585–593 (2015). https://doi.org/10.1007/s13213-014-0894-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13213-014-0894-z