
Abstract
Hurricane Sandy made landfall in the USA on October 29, 2012 and had devastating impacts on human-dominated landscapes in the mid-Atlantic and New England states, but its effects on tidal marsh habitats remain largely undescribed. We evaluated the short-term resilience (a resistance to change or a rapid return to pre-storm conditions) of tidal marshes on Edwin B. Forsythe National Wildlife Refuge, New Jersey by comparing vegetation cover/composition, meadow vole (Microtus pennsylvanicus) relative abundance, and predation rates on artificial bird nests, from three to six months pre-hurricane with seven to 11 months post-hurricane. These three metrics show a high degree of resilience to Hurricane Sandy. Vegetation cover/composition remained similar pre- and post-hurricane, except for five site-specific changes. Although meadow voles were significantly less abundant following the hurricane, we detected a rapid increase from June to July 2013, indicating resiliency via rapid recovery in the population. We also did not observe widespread changes in nest predation rates on artificial nests. Our findings indicate that management actions intended to recover vegetation, small mammals, and nest predators following a large hurricane disturbance may be of less importance than long-term strategies to avoid gradual changes that can reduce ecosystem resilience.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Hurricanes can act as important community-structuring events in many ecosystems, including tidal marshes. Located at the interface of marine and upland environments, tidal marshes absorb hurricane energy and receive both beneficial and detrimental impacts from storms (Michener et al. 1997). Hurricanes can deposit large quantities of sediment, helping marshes grow and accrete in pace with sea-level rise (Rejmánek et al. 1988; Guntenspergen et al. 1995; Nyman et al. 1995; Donnelly et al. 2001), but they can also lower elevations locally through erosion (Morton and Barras 2011). Salinization, and wind and wave energy can cause marsh vegetation loss and the formation of new ponds and channels. In some cases, vegetation recolonizes rapidly (Chabreck and Palmisano 1973; Guntenspergen et al. 1995; Courtemanche et al. 1999), whereas in other locations the altered hydrologic features persist as legacies of the storm event (Morton and Barras 2011). Hurricane impacts on tidal marsh fauna vary based on seasonality, storm severity, and animal mobility. Storm events can reduce population size and growth by direct mortality or by destruction of habitat and food resources, and recovery from such effects is a complex process (Marsh and Wilkinson 1991; Michener et al. 1997; Raynor et al. 2013).
On October 29, 2012, Hurricane Sandy made landfall in New Jersey, USA, and passed over the marsh on Edwin B. Forsythe National Wildlife Refuge (Forsythe NWR). The storm had sustained winds of >100 km/h with >25 cm of precipitation reported in parts of New Jersey (Blake et al. 2013), but the strongest impacts on terrestrial habitats were caused by storm surge and tides (Hall and Sobel 2013). Hurricane Sandy made a nearly perpendicular approach to the coastline, giving it a large fetch over the open ocean and preventing it from being weakened by previous interactions with land surfaces before reaching New Jersey (Hall and Sobel 2013). This trajectory, coupled with full-moon high tides, produced a peak water level of 2.67 m above mean lower low water (MLLW) level at Atlantic City, NJ, ~10 km from Forsythe NWR, causing severe inundation of coastal areas (NOAA / National Ocean Service 2017). Given the potential for an increase in the frequency of large hurricanes due to climate change (Emanuel 2005; Bender et al. 2010), Hurricane Sandy afforded a valuable opportunity to quantify the resilience of coastal ecosystems, enabling more accurate predictions of the effects of future storm events.
Resilience in ecological systems is often defined as the capacity to persist through a disturbance (Gunderson and Holling 2001) or to undergo disturbance without shifting to an alternate stable state (Holling 1973). In forested habitats, measures of resilience to hurricanes have included avian community dynamics, abundance, and survival (Johnson and Winker 2010), as well as tree-stand composition, litterfall, and tree growth rates (Imbert and Portecop 2008). Tidal marsh resilience to hurricanes has been most-widely examined in terms of vegetation, elevation, and sedimentation, but resilience of other ecosystem components and processes has had less attention. For example, vertebrate resilience to hurricanes has largely been described qualitatively and without formal comparison of pre- and post-storm surveys (i.e. Ensminger and Nichols 1958; Gunter and Eleuterius 1971; Cely 1991).
In this study, we evaluated tidal marsh ecosystem resilience on Forsythe NWR to Hurricane Sandy using three metrics: vegetation cover and composition, meadow vole (Microtus pennsylvanicus) abundance, and predation rates on artificial bird nests. We defined resilience as a resistance to change or a rapid return to pre-hurricane conditions, measured over a short temporal scale (three – six months pre-storm compared to seven – 11 months post-storm). By measuring not only structural changes (vegetation) but also alterations in species abundance (meadow voles) and processes (predation rates on artificial nests), we gained a multifaceted perspective of tidal marsh resilience to Hurricane Sandy, enabling us to interpret how interactions between those features influence their resilience. Other studies have reported Hurricane Sandy’s effects on contaminants (Personna et al. 2015; Romanok et al. 2016), barrier beach morphology (Miselis et al. 2016), wetland ecosystem services (Hauser et al. 2015), marsh surface elevation and vegetation (Elsey-Quirk 2016; Rachlin et al. 2017), marsh dieback (Rangoonwala et al. 2016), and regional flood damage (Narayan et al. 2017), but our evaluation provides a novel contribution to this existing body of literature and can help to inform conservation and management actions following future storms.
Methods
Study Area
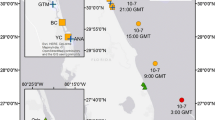
We collected pre-hurricane measurements at Forsythe NWR from June 5 – August 30, 2012 and post-hurricane measurements from June 5 – September 5, 2013. Containing >12,000 ha of tidal marsh, Forsythe NWR supports endemic tidal marsh species of conservation concern and has been designated a Wetland of International Importance under the RAMSAR Convention (U.S. Fish and Wildlife Service 2004; The Ramsar Convention on Wetlands 2014). We sampled at three study sites, which differed in geomorphic setting, landscape context, and management history (Fig. 1). The AT&T site (39.697191° N -74.214032° W) was located in a back-bay marsh along Barnegat Bay, a shallow coastal lagoon with relatively low tidal amplitude, and was historically managed with grid-ditching and open marsh water management (OMWM) for mosquito control. The Oyster Creek site (39.504815° N -74.426283° W) and the Mullica Wilderness site (39.536166° N -74.438021° W) are located within the Great Bay estuary, <20 km from Hurricane Sandy’s landfall. Oyster Creek was historically managed with grid-ditching and OMWM, while Mullica Wilderness contained no hydrologic alterations and is a federally designated wilderness area. Satellite imagery indicated that these three study sites were flooded for at least 12 h and potentially >59 h after the hurricane made landfall (Rangoonwala et al. 2016).

Location of study sites on Edwin B. Forsythe NWR, NJ. Study site abbreviations are: AT = AT&T, MW = Mullica Wilderness, OC = Oyster Creek. Dashed line indicates the path of Hurricane Sandy
Data Collection and Analysis
Vegetation Cover and Composition
We characterized the vegetation cover and composition at a coarse scale and a fine scale at each study site in each year. For the coarse-scale quantification, we surveyed 10 randomly located points, visiting the same points each year. Data were collected from August 16–23, 2012 and July 23 – September 5, 2013. Within a 50-m radius of each point, we ranked six vegetation cover classes (low marsh [Spartina alterniflora], high marsh [S. patens and Distichlis spicata], salt-marsh terrestrial border, invasive species, pannes/pools/creeks, and open water) from zero to six (0 = 0% cover, 0.5 < 1%, 1 = 1–5%, 2 = 6–10%, 3 = 11–25%, 4 = 26–50%, 5 = 51–75%, 6 = 76–100% (Carullo et al. 2007; Neckles et al. 2013)). We used Kruskal-Wallis rank sum tests to compare the six vegetation cover classes between 2012 and 2013 at each site. We controlled for the experiment-wise error rate using the Holm-Bonferroni correction. All analyses throughout this study were conducted using the base package of Program R (version 3.2.0; R Core Team 2014).
For the fine-scale quantification, we surveyed vegetation within 1-m2 quadrats following the methods of Roberts et al. (2017). The quadrats were randomly located within a subplot (14–18 ha) at each site as part of an on-going study of tidal marsh bird demographics (see Kern 2015; Roberts 2016). The number of quadrats was equal to the number of bird nests monitored within each subplot, resulting in 28–66 quadrats surveyed per site per year. Data were collected from June 5 – August 30, 2012 and June 5 – August 2, 2013. Within each quadrat, we estimated the percent of dominant cover types, including low marsh, high marsh, bare ground, wrack (dead vegetation deposited on the marsh by the tide), and open water. We also measured thatch depth (horizontal layer of dead vegetation on the marsh surface) and vegetation height at the center and at the midpoint of each side of the quadrat, and averaged those five measurements to give single estimates of thatch depth and vegetation height for the quadrat. We used t-tests or Kruskal-Wallis rank sum tests when non-normally-distributed data was indicated by D’Agostino’s K-squared tests (Zar 1999) to compare seven fine-scale variables (percent cover of low marsh, high marsh, bare ground, wrack, and open water; thatch depth; and vegetation height) between 2012 and 2013 at each site. We controlled for the experiment-wise error rate using the Holm-Bonferroni correction (Dean and Voss 1999; Zar 1999).
We also examined changes in fine-scale vegetation between pairs of “non-hurricane” years and compared them to the changes from 2012 to 2013. Unlike the other metrics in our study, the fine-scale vegetation measurements were collected over a five-year period (2011–2015), enabling us to analyze three pairs of non-hurricane years (2011–2012, 2013–2014, and 2014–2015) using the methods described above. The number of quadrats surveyed during non-hurricane years ranged from 42 to 162 and the data were collected from June 8 – August 10, 2011; August 14–18, 2014; and June 1 – August 17, 2015. Comparing the changes between non-hurricane years, which approximated background conditions, with those in the year after Hurricane Sandy allowed us to observe whether the hurricane’s impacts stood out from typical inter-annual variation.
Meadow Vole Abundance
We estimated the relative abundance of meadow voles using live-trapping. At each site, we established two 180-m parallel transects, separated by 20 m, forming a 0.36 ha trapping grid (Forys and Dueser 1993; Kruchek 2004; Cameron et al. 2009; Eubanks et al. 2011). In 2012, we used ArcGIS to generate a random location for the trapping grid at each study site, and kept the grids at the same locations in 2013. Along each transect, we placed 10 small Sherman live traps (8 × 9 × 23 cm), spaced at 20-m intervals (20 traps per site). We trapped all sites for three consecutive nights in two rounds per year (round 1 = June 12 – June 21, 2012 and 2013; round 2 = July 24 – August 4, 2012 and July 25 – August 3, 2013), with the exception of round one in 2013 when weather limited trapping to one night at AT&T and two nights at Oyster Creek. We baited traps with a mixture of rolled oats, peanut butter, molasses, and raw apple pieces (Forys and Dueser 1993; Kruchek 2004; Cameron et al. 2009; Eubanks et al. 2011), and placed the bait behind the treadle in the rear of the trap and at the trap entrance. We attached the traps to Styrofoam platforms, secured between two 0.5-m bamboo stakes, to allow flotation during daily tidal changes (Wolfe 1985; DeSa et al. 2012). The duration of each trapping night was 12 h (18:00 to 6:00 h) and we did not trap during heavy rainfall, storms, or lunar (monthly) high tides. All captured animals were identified to species, weighed, and released at the point of capture. Maximum handling time for captured animals was 5 min.
We quantified the landscape context of the three trapping grids using landcover data collected in 2010 by the National Oceanic and Atmospheric Administration’s Coastal Change Analysis Program (National Oceanic and Atmospheric Administration (NOAA) 2014). Using ArcGIS, we calculated the percent of four dominant cover types within a 1000-m radius of each trapping grid. The four cover types were: tidal marsh, vegetated non-marsh (primarily forest), water, and development (roads and buildings). We also calculated the distance from the midpoint of each grid to the nearest non-marsh edge (either vegetated or developed) using the ‘near’ tool in ArcGIS (Table 1).
We examined the effect of Hurricane Sandy on meadow voles by calculating an index of relative abundance for each round, by site and year. The index was created by dividing the total number of vole captures each night by the number of traps set that night (usually 20, but occasionally trap malfunctions occurred). We then averaged the index values across the three nights of each round to give a single relative abundance value (mean captures/trap night) and associated estimate of variance for each round, by site and year. We used a paired t-test to examine differences in relative abundance between years for each round at each site. Within each round of each year, we also tested for differences in relative abundance between sites using a Kruskal-Wallis rank sum test because data were non-normal.
Artificial Nest Predation Rates
To estimate the resilience of the nest predator community, we monitored artificial nests containing clay eggs. Although artificial nests should not be used to estimate actual nest predation rates (Thompson and Burhans 2004), they can be used to identify nest predators and to detect nest predation trends (Wilson et al. 1998; Boulton and Cassey 2006). The artificial nest monitoring occurred immediately prior to the small mammal trapping along the same transects used for the trapping (above). We placed 10 brown, wicker nests (10–12 cm in diameter) along each transect, spaced at 20-m intervals, for a total of 20 nests per site. We deployed and monitored the artificial nests in two rounds per year (round one = June 1–14, 2012 and June 2–14, 2013; round two = July 16–26, 2012 and July 11–26, 2013). Nests were designed to mimic tidal marsh sparrow nests. Each nest was placed 0–15 cm off the marsh surface, concealed in vegetation, and contained two off-white, non-hardening clay eggs. We monitored nests every 2–4 days, for a total of 10–12 days per round. At each nest check, we examined the eggs for evidence of predation, including beak or bite marks.
For each site, we compared the proportion of depredated nests between years using Fisher’s Exact Test of Independence. A nest was considered depredated if one or both of the eggs was found with beak or bite marks at any point during the 10–12 days of monitoring. For sites that showed a difference in predation rates between years, we investigated whether the cause of predation (avian versus mammalian) differed between years using Fisher’s Exact Test of Independence.
Tide Height
To further characterize hurricane versus non-hurricane years, we used daily high tide levels during our study period (January 1, 2011 – August 17, 2015) at the Atlantic City tide station, located 10 km from Forsythe NWR. We examined individual high-water events and the range of daily tidal maxima for each year, relative to the mean lower low water (MLLW) tidal datum (NOAA / National Ocean Service 2017).
Results
The vegetation within the study sites remained largely unchanged pre- and post-Hurricane Sandy. We detected no change of vegetation in the coarse-scale surveys (Table 2). The fine-scale surveys revealed five site-specific changes in low marsh cover, high marsh cover, bare ground cover, and vegetation height (Table 3); however, differences were not consistent across sites and the number of differences was within the range observed between non-hurricane year pairs (two in 2011–2012 to seven in 2014–2015; Fig. 2). Furthermore, differences from 2012 to 2013 were generally of a magnitude similar to those observed between non-hurricane year pairs (Fig. 2). A notable exception to this pattern is Oyster Creek, where low marsh cover was 1.7 times higher and high marsh cover was 2.2 times lower in 2012 than in 2013 (Table 3). These changes were greater in magnitude and opposite in direction from the changes between non-hurricane years at that site (Fig. 2). Summary statistics and results of statistical tests for the non-hurricane year data can be found in Online Resource 1.

Mean differences of fine-scale vegetation variables (low marsh cover, high marsh cover, bare ground cover, wrack cover, water cover, thatch depth, and vegetation height) between year pairs (2011–2012, 2012–2013, 2013–2014, and 2014–2015) at three sites at Edwin B. Forsythe NWR, NJ. Site abbreviations are: AT = AT&T, MW = Mullica Wilderness, OC = Oyster Creek. Asterisks indicate a significant difference in the vegetation variable between years (significance was evaluated using t-tests or Kruskal-Wallis tests followed by the Holms-Bonferroni correction with α = 0.05)
During 2012 and 2013, we trapped for 33 nights (620 total trap nights) and had 206 meadow vole captures. No other species were detected. The trapping effort was similar among sites and years (Table 1). Meadow vole relative abundance decreased dramatically following Hurricane Sandy; we did not detect any voles during the first round of trapping in 2013 (Fig. 3). Abundance was greater in 2012 than in 2013 at all sites for round one (t0.05, 2 = 6.41, P = 0.02) and round two (t0.05, 2 = 5.50, P = 0.03). However, vole abundance in 2013 followed a similar pattern to that observed in 2012; namely, abundance increased from round one to round two, presumably as new individuals were recruited into the population (Fig. 3). Although we did not record the sex of captured voles, we made several observations of meadow vole nests on the study sites in all years (R.A. Longenecker and S.G. Roberts, unpublished data). Therefore, despite the severe decrease in vole abundance following Hurricane Sandy, the population appeared to have begun to recover by late July 2013.

Meadow vole (Microtus pennsylvanicus) relative abundance (mean captures/trap night ±95% confidence intervals) by study site, year, and round at Edwin B. Forsythe NWR, NJ. Study site abbreviations are: AT = AT&T, MW = Mullica Wilderness, and OC = Oyster Creek. Round 1 was conducted from June 12 – June 21, and round 2 was conducted from July 24 – August 4. Confidence intervals are dashed lines for 2012 and solid lines for 2013
Despite site differences in geomorphic setting, management history, and landscape context, patterns of meadow vole relative abundance were similar across AT&T, Mullica Wilderness, and Oyster Creek (Fig. 3). Abundance for each round in each year did not differ among sites (all Kruskal-Wallis tests: χ20.05, 2 > 0.40, P > 0.60).
Predation of artificial nests declined 17.0% at Mullica Wilderness and 38.0% at Oyster Creek pre-Hurricane Sandy (2012) versus post-Hurricane Sandy (2013), but these declines were not statistically significant (P = 0.60 and 0.15, respectively). However, at AT&T artificial nest predation declined 44.3% (P < 0.001; Table 1) during the same period. Despite the decrease at AT&T in the year following the hurricane, the primary cause of nest failure, mammals, remained the same in both years (P = 0.19). Damage to clay eggs suggested that mammals caused four times as many failures than birds in 2012 and ten times as many in 2013.
Based on Atlantic City tide gage data, a single high-tide event on the scale of Hurricane Sandy was not observed in any of the non-hurricane years. The maximum daily high tide in 2012 was 2.67 m (October 29), which was 0.50–0.69 m greater than the maximum in 2011, 2013, 2014, and 2015. Excluding Hurricane Sandy, the range of daily tidal maxima was similar among years (0.89–2.19 m, 0.69–2.17 m, 0.60–2.13 m, 0.68–2.19 m, 0.68–1.98 m, respectively).
Discussion
We found substantial resilience to Hurricane Sandy in all metrics examined, both in the absence of change and in a rapid return to pre-hurricane conditions when change did occur. Daily tidal maxima were relatively consistent across years, excluding Hurricane Sandy, and no other high-tide events on the scale of Hurricane Sandy were observed. We detected only minimal changes in vegetation cover. Although there was a trend of increased bare ground at all sites after the hurricane, indicating that some vegetation may have been removed by wave energy or died from prolonged inundation, bare ground remained uncommon (<10% cover). These findings support those of other field studies in New Jersey tidal marshes, including Elsey-Quirk (2016), who reported no evidence of widespread wrack, sediment deposition, or vegetation removal, and Rachlin et al. (2017), who found no change in vegetation composition. In contrast, several studies using aerial photography (Hauser et al. 2015) or remote sensing methods (e.g., normalized difference vegetation index or NDVI (Rangoonwala et al. 2016) reported large and potentially severe impacts to marsh vegetation on Forsythe NWR, and, specifically, that our study site locations experienced pond/channel creation or expansion, severe natural debris deposition, marsh dieback, and/or large changes in marsh condition (greenness). However, the only vegetation changes detected by our field data were fine-scale and minimal; we did not detect changes of the severity and/or extent as reported by Hauser et al. (2015) or Rangoonwala et al. (2016). This discrepancy between the results of field-based and remote sensing methods warrants further investigation.
We observed resilience in the meadow vole population through its rapid increase in relative abundance after initial decimation following Hurricane Sandy. Because vegetated habitat was not lost, we conclude that inundation from storm surge, high tides, and rainfall was a likely cause of the decline in meadow voles. Other small mammals in North American marshes and beaches, including raccoon (Procyon lotor), beach mouse (Peromyscus polionotus), and Eastern harvest mouse (Reithrodontomys humulis), have been impacted by storm flooding through either direct mortality or alteration of movement patterns (Gunter and Eleuterius 1971; Swilling et al. 1998; Klinger 2006; Pries et al. 2009). Although meadow voles can swim and dive (Harris 1953) and can experience erratic population fluctuations (Getz et al. 2001; Getz et al. 2006), the complete absence of the species from all three sites in June 2013 strongly suggests that prolonged flooding during the hurricane drowned the majority of individuals on the tidal marsh in New Jersey. During a damage assessment immediately following the hurricane, Forsythe refuge staff did not observe any mats of wrack caught in tidal marsh shrubs; instead, all wrack was concentrated along the upland borders of the marshes. Wrack mats can provide refugia for small mammals during a flood, so their absence from the marsh further supports our conclusion that meadow voles were extirpated by high water levels (P.M. Castelli, unpublished data). Although vole abundance at our study sites nine months post-hurricane had not reached pre-hurricane levels, it showed signs of recovery and resilience irrespective of site differences in geomorphic setting, management history, and landscape context. Additional study of meadow vole natural history in tidal marshes is needed.
Artificial nest predation may have been reduced by Hurricane Sandy, but the results are not conclusive at all sites. Following the storm, nest failure rates decreased at AT&T, but did not change at the other study sites. Given the observed decrease in meadow vole abundance, it is plausible that other small mammals that are nest predators, such as Norway rats (Rattus norvegicus) and rice rats (Oryzomys palustris), were similarly impacted, resulting in fewer nest failures at AT&T. Study site differences may have masked or disrupted this effect at Mullica Wilderness and Oyster Creek. Thus, we are not able to conclude whether Hurricane Sandy affected artificial nest predation or nest predators.
Although our study provides insight into the short-term resiliency of marshes to Hurricane Sandy, resilience also operates at other spatial and temporal scales (Carpenter et al. 2001). We defined resilience in terms of persistence – did these three components/processes of the tidal marsh remain similar after the hurricane to the way they were before? – and at a one-year time scale. Further research could evaluate the impact of the hurricane on longer-term processes, such as meadow vole population growth rate, in order to better understand how such stochastic disturbances disrupt or are absorbed over decadal time-spans. Short-term resilience does not necessarily indicate that the hurricane had no detectable impacts; some changes simply may require more time to become evident. For example, hurricanes in tropical forests have been found to indirectly impact vertebrate communities over several years by altering tree fruit production (Klinger 2006). Tidal marsh resilience could also be examined at different spatial scales, in order to capture the impact of the hurricane on metapopulation dynamics, which is important for predicting re-colonization patterns of locally extirpated species.
Finally, although we found resilience in three metrics, caution should be used in generalizing this result to other aspects of the ecosystem or to different tidal marshes. For example, we predict that other resident, small mammal species that use the tidal marsh were extirpated in a similar fashion as meadow voles. However, more mobile species, such as raccoons or red foxes (Vulpes vulpes), may not have been affected as drastically. We also predict that nest predation rates of endemic, tidal marsh birds, such as Saltmarsh (Ammodramus caudacutus) and Seaside (A. maritimus) sparrows, did not dramatically differ after the hurricane because we detected no change in artificial nest predation. However, it does not follow that overall breeding success of tidal marsh birds was similarly unaffected. The hurricane could have altered resources needed for appropriate nest placement, such as thatch availability, or for the successful rearing of nestlings, such as invertebrate food sources. Lastly, our study was conducted in a large, protected, relatively intact tidal marsh complex; tidal marshes in degraded condition may not have demonstrated the same level of resilience. With the exception of grid-ditching and OMWM, our study sites had relatively natural hydrology, which is perhaps the most important physical driver of the ecosystem (Mitsch and Gosselink 2000; U.S. Fish and Wildlife Service 2004). Many tidal marshes elsewhere on the Atlantic Coast, however, have been disconnected from tidal flow and sedimentation by roads, tide gates, or weirs, which fundamentally impact marsh structure and function (Crain et al. 2009), are related to bird population declines (Correll et al. 2017), and may influence the resilience of marshes to hurricanes. Irrespective of marsh management or condition, our study sites were generally representative of other marshes within the region in terms of vegetation composition. Recent remote-sensing analyses, using aerial imagery and elevation data, estimated that high and low marsh were the most common cover types within northeast tidal marshes from Maine to Virginia (36% and 21%, respectively; Correll et al. in review). Using the marsh vegetation classification layers produced by Correll et al. (available at www.tidalmarshbirds.org), we determined that our study sites were largely high marsh (81%), followed by low marsh (12%), which is also true for other marshes in south-central New Jersey (54% and 21%, respectively). The main difference in cover types is terrestrial border habitat, which is more common in south-central New Jersey and the entire Northeast than at our study sites (10%, 24%, and 0.11%, respectively), because our sites were located in the center of large marsh complexes.
Ecosystem resilience may be crucial for maintaining biodiversity and ecological processes in light of global climate change (Mawdsley et al. 2009). Global climate change and sea level rise pose significant threats to the persistence of coastal wetlands and their endemic species (Van De Pol et al. 2010; Bayard and Elphick 2011; Correll et al. 2017; Field et al. 2017). The frequency and intensity of hurricanes in the Atlantic may increase due to global climate change (Emanuel 2005; Webster et al. 2005; IPCC 2014), so quantifying the resilience of tidal marshes to Hurricane Sandy can inform future management and conservation action. Our findings suggest that a future hurricane of similar magnitude, duration, and timing is not likely to cause extensive vegetation loss or to greatly affect nest predation rates on a relatively intact, high-marsh-dominated site, although small mammal abundance may be temporarily reduced. In such a situation, no immediate management actions targeted at recovering vegetation, small mammals, or nest predators would need to be implemented, aside perhaps from clearing human debris deposited by storm surge. Instead, managers should focus on long-term strategies to maintain or increase the resilience of the tidal marsh. Although ecosystem resilience is often measured in response to an abrupt disturbance, it is also affected by gradual changes that occur over long time spans (Scheffer et al. 2001). How resilient a tidal marsh can be to a hurricane may be determined by how greatly it has been weakened by small, on-going changes, such as eutrophication or lack of sedimentation (Scheffer et al. 2001). Thus, increasing the likelihood of tidal marsh persistence through global climate change requires managers, policy-makers, and conservationists to address the gradual changes that influence resilience, rather than simply respond to large disturbances.
References
Bayard TS, Elphick CS (2011) Planning for sea-level rise: quantifying patterns of saltmarsh sparrow (Ammodramus caudacutus) nest flooding under current sea-level conditions. Auk 128:393–403
Bender MA, Knutson TR, Tuleya RE, Sirutis JJ, Vecchi GA, Garner ST, Held IM (2010) Modeled impact of anthropogenic warming on the frequency of intense Atlantic hurricanes. Science 327:454
Blake ES, Kimberlain TB, Berg RJ, Cangialosi JP, Beven JLI (2013) Tropical cyclone report: hurricane Sandy AL182012. National Hurricane Center, Miami
Boulton RL, Cassey P (2006) An inexpensive method for identifying predators of passerine nests using tethered artificial eggs. New Zealand Journal of Ecology 30:377–385
Cameron GN, Williams JM, Kruchek BL (2009) Seasonal dynamics of small mammals and vegetation in a gulf cordgrass community. The Southwestern Naturalist 54:237–247
Carpenter S, Walker B, Anderies JM, Abel N (2001) From metaphor to measurement: resilience of what to what? Ecosystems 4:765–781
Carullo M, Carlisle BK, Smith JP (2007) A New England rapid assessment method for assessing condition of estuarine marshes: a Boston Harbor, cape cod and islands pilot study. Massachusetts Office of Coastal Zone Management, Boston
Cely JE (1991) Wildlife effects of hurricane Hugo. Journal of Coastal Research 8:319–326
Chabreck RH, Palmisano AW (1973) The effects of hurricane Camille on the marshes of the Mississippi River delta. Ecology 54:1118–1123
Correll MD, Wiest WA, Hodgman TP, Shriver WG, Elphick CS, McGill BJ, O'Brien KM, Olsen BJ (2017) Predictors of specialist avifaunal decline in coastal marshes. Conservation Biology 31:172–182
Courtemanche RPJ, Hester MW, Mendelssohn IA (1999) Recovery of a Louisiana barrier island marsh plant community following extensive hurricane-induced overwash. Journal of Coastal Research 15:872–883
Crain CM, Gedan KB, Dionne M (2009) Tidal restrictions and mosquito ditching in New England marshes: case studies of the biotic evidence, physical extent, and potential for restoration of altered tidal hydrology. In: Silliman BR, Grosholz ED, Bertness MD (eds) Human impacts on salt marshes: a global perspective. University of California Press, California, pp 149–170
Dean A, Voss D (1999) Design and analysis of experiments. Springer-Verlag New York, Inc., New York
DeSa MA, Zweig CL, Percival HF, Kitchens WM, Kasbohm JW (2012) Comparison of small-mammal sampling techniques in tidal salt marshes of the central Gulf Coast of Florida. Southeastern Naturalist 11:89–100
Donnelly JP, Roll S, Wengren M, Butler J, Lederer R, Webb T (2001) Sedimentary evidence of intense hurricane strikes from New Jersey. Geology 29:615–618
Elsey-Quirk T (2016) Impact of hurricane Sandy on salt marshes of New Jersey. Estuarine, Coastal and Shelf Science 183:235–248
Emanuel K (2005) Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436:686–688
Ensminger AB, Nichols LG (1958) Hurricane damage to Rockefeller refuge. Proceedings of the annual conference, southeastern association of game and fish commissioners pp 52–56
Eubanks BW, Hellgren EC, Nawrot JR, Bluett RD (2011) Habitat associations of the marsh rice rat (Oryzomys palustris) in freshwater wetlands of southern Illinois. Journal of Mammalogy 92:552–560
Field CR, Bayard TS, Gjerdrum C, Hill JM, Meiman S, Elphick CS (2017) High-resolution tide projections reveal extinction threshold in response to sea-level rise. Global Change Biology 23:2058–2070
Forys EA, Dueser RD (1993) Inter-island movements of rice rats (Oryzomys palustris). The American Midland Naturalist 130:408–412
Getz LL, Hofmann JE, McGuire B, Dolan TW (2001) Twenty-five years of population fluctuations of Microtus ochrogaster and M. pennsylvanicus in three habitats in east-central Illinois. Journal of Mammalogy 82:22–34
Getz LL, Oli MK, Hofmann JE, McGuire B (2006) Vole population fluctuations: factors that initiate and determine intervals between them in Microtus ochrogaster. Journal of Mammalogy 87:387–393
Gunderson LH, Holling CS (2001) Panarchy: understanding transformations in human and natural systems. Island Press, Washington, D.C.
Guntenspergen GR, Cahoon DR, Grace J, Steyer GD, Fournet S, Townson MA, Foote AL (1995) Disturbance and recovery of the Louisiana coastal marsh landscape from the impacts of hurricane Andrew. Journal of Coastal Research 21:324–339
Gunter G, Eleuterius LN (1971) Some effects of hurricanes on terrestrial biota, with special reference to Camille. Gulf Research Reports 3:283–289
Hall TM, Sobel AH (2013) On the impact angle of hurricane Sandy’s New Jersey landfall. Geophysical Research Letters 40:2312–2315
Harris VT (1953) Ecological relationships between meadow voles and rice rats in tidal marshes. Journal of Mammalogy 34:479–487
Hauser S, Meixler MS, Laba M (2015) Quantification of impacts and ecosystem services loss in New Jersey coastal wetlands due to hurricane Sandy storm surge. Wetlands 35:1137
Holling CS (1973) Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4:1–23
Imbert D, Portecop J (2008) Hurricane disturbance and forest resilience: assessing structural vs. functional changes in a Caribbean dry forest. Forest Ecology and Management 255:3494–3501
IPCC (2014) Climate change 2014: impacts, adaptation, and vulnerability. Part a: global and sectoral aspects. Contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University press, Cambridge and New York
Johnson AB, Winker K (2010) Short-term hurricane impacts on a neotropical community of marked birds and implications for early-stage community resilience. PLoS One 5:e15109
Kern RA (2015) Conservation ecology of tidal marsh sparrows in New Jersey. University of Delaware, Dissertation
Klinger R (2006) The interaction of disturbances and small mammal community dynamics in a lowland forest in Belize. The Journal of Animal Ecology 75:1227–1238
Kruchek BL (2004) Use of tidal marsh and upland habitats by the marsh rice rat (Oryzomys palustris). Journal of Mammalogy 85:569–575
Marsh CP, Wilkinson PM (1991) The impact of hurricane Hugo on coastal bird populations. Journal of Coastal Research 8:327–334
Mawdsley JR, O'Malley R, Ojima DS (2009) A review of climate-change adaptation strategies for wildlife management and biodiversity conservation. Conservation Biology 23:1080–1089
Michener WK, Blood ER, Bildstein KL, Brinson MM, Gardner LR (1997) Climate change, hurricanes and tropical storms, and rising sea level in coastal wetlands. Ecological Applications 7:770–801
Miselis JL, Andrews BD, Nicholson RS, Defne Z, Ganju NK, Navoy A (2016) Evolution of mid-Atlantic coastal and back-barrier estuary environments in response to a hurricane: implications for barrier-estuary connectivity. Estuaries and Coasts 39:916–934
Mitsch WJ, Gosselink JG (2000) Wetlands. John Wiley, New York
Morton RA, Barras JA (2011) Hurricane impacts on coastal wetlands: a half-century record of storm-generated features from southern Louisiana. Journal of Coastal Research:27:27–27:43
Narayan S, Beck MW, Wilson P, Thomas CJ, Guerrero A, Shepard CC, Reguero BG, Franco G, Ingram JC, Trespalacios D (2017) The value of coastal wetlands for flood damage reduction in the northeastern USA. Scientific Reports 7:9463
National Oceanic and Atmospheric Administration (NOAA) (2014) Coastal change analysis program regional land cover. http://www.csc.noaa.gov/digitalcoast/data/ccapregional. Accessed 18 Feb 2014
Neckles HA, Guntenspergen GR, Shriver WG, Danz NP, Wiest WA, Nagel JL, Olker JH (2013) Identification of metrics to monitor salt marsh integrity on National Wildlife Refuges in relation to conservation and management objectives. Final report to U.S. Fish and Wildlife Service. Northeast Region. USGS Patuxent Wildlife Research Center, Laurel, p 226
NOAA / National Ocean Service (2017) Center for Operational Oceanographic Products and Services. www.tidesandcurrents.noaa.gov. Accessed 16 Dec 2017
Nyman JA, Crozier CR, DeLaune RD (1995) Roles and patterns of hurricane sedimentation in an estuarine marsh landscape. Estuarine, Coastal and Shelf Science 40:665–679
Personna YR, Geng X, Saleh F, Shu Z, Jackson N, Weinstein MP, Boufadel MC (2015) Monitoring changes in salinity and metal concentrations in New Jersey (USA) coastal ecosystems post-hurricane Sandy. Environmental Earth Sciences 73:1169–1177
Pries AJ, Branch LC, Miller DL (2009) Impact of hurricanes on habitat occupancy and spatial distribution of beach mice. Journal of Mammalogy 90:841–850
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rachlin JW, Stalter R, Kincaid D, Warkentine BE (2017) The effect of superstorm Sandy on salt marsh vascular flora in the New York bight. The Journal of the Torrey Botanical Society 144:40–46
Rangoonwala A, Enwright NM, Ramsey E III, Spruce JP (2016) Radar and optical mapping of surge persistence and marsh dieback along the New Jersey mid-Atlantic coast after hurricane Sandy. International Journal of Remote Sensing 37:1692–1713
Raynor EJ, Pierce AR, Owen TM, Leumas CM, Rohwer FC (2013) Short-term demographic responses of a coastal waterbird community after two major hurricanes. Waterbirds 36:88–93
Rejmánek M, Sasser CE, Peterson GW (1988) Hurricane-induced sediment deposition in a Gulf Coast marsh. Estuarine, Coastal and Shelf Science 27:217–222
Roberts SG (2016) Population viability of seaside and saltmarsh sparrows in New Jersey. University of Delaware, Master’s thesis
Roberts SG, Longenecker RA, Etterson MA, Ruskin KJ, Elphick CS, Olsen BJ, Shriver WG (2017) Factors that influence vital rates of seaside and saltmarsh sparrows in coastal New Jersey, USA. Journal of Field Ornithology 88:115–131
Romanok KM, Szabo Z, Reilly TJ, Defne Z, Ganju NK (2016) Sediment chemistry and toxicity in Barnegat Bay, New Jersey pre- and post-hurricane Sandy, 2012–13. Marine Pollution Bulletin 107:472–488
Scheffer M, Carpenter S, Foley JA, Folke C, Walker B (2001) Catastrophic shifts in ecosystems. Nature 413:591
Swilling WR Jr., Wooten MC, Holler NR, Lynn WJ (1998) Population dynamics of Alabama beach mice (Peromyscus polionotus ammobates) following hurricane opal. The American Midland Naturalist 140:287–298
The Ramsar Convention on Wetlands (2014) The Ramsar list of wetlands of international importance. http://www.ramsar.org. Accessed 24 Mar 2014
Thompson FRI, Burhans DE (2004) Differences in predators of artificial and real songbird nests: evidence of bias in artificial nest studies. Conservation Biology 18:373–380
U.S. Fish and Wildlife Service (2004) Edwin B. Forsythe national wildlife refuge comprehensive conservation plan. U.S. Fish and Wildlife Service, Hadley, Massachusetts
Van De Pol M, Ens BJ, Heg D, Brouwer L, Krol J, Maier M, Exo K, Oosterbeek K, Lok T, Eising CM, Koffijberg K (2010) Do changes in the frequency, magnitude and timing of extreme climatic events threaten the population viability of coastal birds? Journal of Applied Ecology 47:720–730
Webster PJ, Holland GJ, Curry JA, Chang H (2005) Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309:1844–1846
Wilson GR, Brittingham MC, Goodrich LJ (1998) How well do artificial nests estimate success of real nests? Condor 100:357–364
Wolfe JL (1985) Population ecology of the rice rat (Oryzomys palustris) in a coastal marsh. Journal of Zoology 205:235–244
Zar JH (1999) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgements
We thank the staff at Forsythe NWR for their assistance and support, particularly V. Rettig for logistical support, and J. Heise and M. Schvetz for conducting vegetation and small mammal surveys. We also thank A. Anholt, M. Fuirst, M. Grasso, H. Kostick, K. Lewis, A. Mankofsky, N. Pearson, T. Pitcher, M. Rhode, and J. Warner for collecting data. Dr. W. Glanz made critical contributions to the conceptualization and design of this project, and we are grateful for his assistance. We thank K. McCarthy for providing comments on previous versions of this manuscript. Funding was provided by the U.S. Fish and Wildlife Service (State Wildlife Grant # U2-5-R-1) and the National Science Foundation (DEB-1340008). This research was conducted with the following permits and permissions: University of Delaware Animal Care and Use Committee (AUP 1231-2012-0 and 1231-2013-1) and New Jersey Division of Fish and Wildlife Scientific Collecting Permit (SC 2012_097).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Longenecker, R.A., Bowman, J.L., Olsen, B.J. et al. Short-Term Resilience of New Jersey Tidal Marshes to Hurricane Sandy. Wetlands 38, 565–575 (2018). https://doi.org/10.1007/s13157-018-1000-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-018-1000-2