
Abstract
Data on canopy trees (stems ≥ 15 cm DBH) in riparian wetlands, spanning from headwaters to large river floodplains, were used to test whether forest canopy composition differed among hydrogeomorphic (HGM) riverine subclasses and among physiographic sub-regions (Major Land Resource Areas; MLRA) within a given HGM subclass. Riverine stands (n = 225) were sampled in four MLRA regions of the Southeastern U.S. Atlantic and Gulf Coastal Plain Physiographic Provinces. Composition data were analyzed using Non-metric Multidimensional Scaling and Multiple-Response Permutation Procedures to evaluate differences among HGM subclasses and MLRA regions. Analyses showed that canopy composition differed among three a priori subclasses related to Strahler stream order: headwater complex (along 1st-3rd order streams), mid-gradient floodplain (4th-6th order), and low-gradient floodplain (> 6th order). Further, composition also differed by MLRA region within each subclass. Thus, not only was species composition related to riverine hydrogeomorphology across a wide physiographic area, but differences in composition within HGM subclasses were also related to sub-region. These data could be useful in defining floristic reference standards when evaluating floodplain condition in southeastern USA Coastal Plain stream networks.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forested floodplains of Southeastern U.S. rivers have been studied extensively over the past 30 years, thus much is known about how they function and the species that inhabit them (Wharton et al. 1982; Brinson 1990; Sharitz and Mitsch 1993). Numerous regional studies have also characterized the forested wetlands associated with smaller Southeastern Coastal Plain streams (Gemborys and Hodgkins 1971; Glascock and Ware 1979; Parsons and Ware 1982; Bledsoe 1993; Rheinhardt et al. 1998, 2000, 2007, 2012). Within such a large physiographic region, the composition of mature riverine forests might be expected to vary in consistent ways, both geographically and across the stream network hydrogeomorphic gradient from headwaters to the lowest non-tidal reaches. However, little work has explicitly analyzed variation in canopy composition along the regional upstream/downstream network. Data of this sort would be valuable in understanding patterns of riverine forest composition spanning the stream network, and could provide resource managers with information for evaluating the local conditions of riverine resources. Further, a geographically extensive data set could be used to test the premise of Brinson (1993b) that differences in wetland hydrogeomorphology (HGM type) should influence differences in ecosystem functioning among unaltered wetlands.
Forest structure and canopy composition are important to a suite of key riverine wetland functions. The forest canopy stores > 90 % of aboveground biomass (Rheinhardt et al. 2012), determines biodiversity of arboreal fauna via habitat stratification (Odum 1969; Dickson and Noble 1978), regulates microclimatic regimes from canopy to forest floor (Parker 1995), determines forest floor light regime and its relationship to colonization by shade-intolerant exotic plant species (Loehle 2003), controls the quantity and quality of large detritus used by saprophytic organisms on the forest floor (Laiho and Prescott 1999), affects the amount of particulate and dissolved organic carbon utilized in situ and exported downstream and to hyporheic zones (Hope et al. 1994; Warren et al. 2007; Wipfli et al. 2007), and supplies most of the root exudates that drive microbial nutrient transformations in forest soils (Martin et al. 1999). Canopy composition can influence the type and quality of wildlife forage (mast and soft fruit for animals, nectar for insects) (Wigley and Roberts 1994; Rodewald and Abrams 2002), the distribution of insects on tree surfaces (Robinson and Holmes 1984; Whelan 2001), and the quality and availability of nutrients (Prescott 2002). The age (maturity) and composition of the canopy stratum together provide important information on forest integrity and the capacity of a forest ecosystem to function in a characteristic manner.
Hydrologic regime has repeatedly been shown to be a major factor controlling the distribution of wetland plant species. Light and nutrient availability have also been linked to the composition of floodplain forests (Wharton et al. 1982; Bedford et al. 1999; Denslow and Battaglia 2002). At local scales, forest composition integrates environmental factors and disturbances over long time periods. At larger geographic scales, climate and underlying geology also affect riverine forest composition. Climate affects precipitation patterns, relative humidity, and growing season, while geology affects the type and rates of sediment delivery to floodplains. Therefore, we expected that vegetation would vary among physiographic provinces (e.g., Piedmont vs. Coastal Plain), even within a given HGM type, but we were uncertain how much composition might vary within a single, expansive province like the U.S. Coastal Plain. Understanding this variability would be useful for resource managers evaluating or tracking the condition of riverine wetland resources.
Considering that hydrogeomorphology and geographic region could potentially influence forest canopy composition, we undertook an analysis of forest stand data representing three different riverine subclasses in several recognized physiographic subregions across the extensive Atlantic and Gulf Coastal Plain Physiographic Province of the southeastern USA. Specifically, the study had three primary objectives: (1) obtain comparative quantitative data on canopy composition across the entire Coastal Plain Province, (2) determine if canopy species composition differs significantly among three a priori defined riverine subclasses, and (3) test whether canopy composition differs significantly within the subclasses relative to recognized subregions of the Coastal Plain.
Classification of Riverine Wetlands and Subregions
Given the potential importance of hydrodynamics to forest species composition, we defined riverine wetland subclasses using an HGM classification approach (Brinson 1993b; Brooks et al. 2011; Tiner 2011), which links differences in wetland hydrogeomorphology to differences in ecosystem functions (Brinson 1993a). Three a priori subclasses were defined related to stream order (sensu Strahler 1952): headwater complex, mid-gradient floodplain, and low-gradient floodplain (sensu Tiner 2011). Headwater complexes (1st to 3rd order) are fed primarily by groundwater. They are numerous relative to higher order streams, constituting 70–90 % of stream length in a typical Coastal Plain drainage basin (Rheinhardt et al. 1999), and are critical source areas for organic matter, invertebrates, and wood to downstream reaches (Wipfli et al. 2007). Headwater complexes include bayheads (sensu Monk 1966), which drain organic-rich, nutrient-poor soils of flat, interstream divides. During normal precipitation years, most headwater streams (1st to 3rd order) flow throughout the winter but cease flowing in mid-summer during periods of minimal precipitation and maximum evapotranspiration (Brinson et al. 2006) and under drought conditions. Headwater streams coalesce to form larger, mid-gradient streams (4th to 6th order), which in turn join additional streams to become even larger and more energetic rivers (Strahler 1952). Ten to 30 % of stream length in a Coastal Plain drainage network consists of mid-gradient and larger low-gradient (> 6th order) rivers. In hydrologically unaltered Coastal Plain stream networks, mid- to low-gradient reaches inundate their floodplains in the late winter and early spring of most years, and occasionally in the summer and fall during and after tropical storms.
To define Coastal Plain subregions, we used the USDA (2006) Major Land Resource Areas (MLRA), which are characterized by similar patterns of soils, geology, climate, water resources, and potential vegetation. These MLRA subregions seemed like a reasonable scale at which to examine compositional variability among floodplain forests.
Methods
Study Area
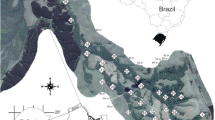
Data were collected from forested stands in the four largest of 12 MLRA subregions of the Coastal plain: Southern Coastal Plain (SCP), Western Coastal Plain (WCP), Southern Mississippi Valley Loess (MVL), and Atlantic Coast Flatwoods (ACF) (Table 1, Fig. 1). Topographic relief across the study area ranges from the extremely broad, flat, low-elevation, coastal terraces of the Atlantic Coastal Plain to the moderately-hilly terrain of north-central Alabama (elevations to 600 ft) in the Southern Coastal Plain. Even across this varied upland landscape, stream gradient is low (< 0.5 % slopes) (Rheinhardt et al. 1998), especially among 4th and higher order streams.

Atlantic and Gulf Coastal Plain physiographic area, and locations of the 225 sampled stands within four major MLRA regions
Forest stand locations could not be located randomly, but potential sampling locations were chosen to represent the variety of settings typical of stream networks in the geographic regions studied. Some locations were suggested by local resource managers; others were known from previous vegetation studies conducted by the authors and from locations identified by the Carolina Vegetation Survey (http://cvs.bio.unc.edu/). Other locations were located using remotely sensed data, primarily the most recent aerial photography available.
Field Data
Field data were collected at the scale of a stand, defined as an area greater than one hectare and homogeneous with respect to vegetation, cover type, and age. Sections of floodplain forests that varied in age (e.g., time since last clearcut) were differentiated as separate stands. The intent was to obtain quantitative data to define and calibrate HGM functional assessment logic models being developed to characterize functional capacity of wetlands associated with riverine ecosystems (Wilder et al. 2013). Besides collecting data on canopy composition, a large amount of additional quantitative data were collected on stream channel characteristics, geomorphic attributes, soils, amount and types of detritus, forest structure and age, the number and cover of strata, and species composition of subcanopy strata. A categorical score ranging from 1 to 5 was also assigned to each stand to represent the stand’s overall condition, based on the type and intensity of a combination of potential alterations listed on data sheets, and best professional judgment (BPJ), as defined below. Both type and relative intensity of alterations, when present, were used to calibrate BPJ scores. The same field data sheets were used for all stands and co-author Wilder spent time in the field with all the other field crews to ensure that data collection protocols were identical, including decision-making used to determine BPJ scores. A determination of BPJ condition was based on the degree to which the following parameters had been altered:
-
(1)
vegetation and forest structure (e.g., condition based on stand age, extent of any missing or altered strata, extent of invasive species cover, and presence and intensity of forest silvicultural management);
-
(2)
surrounding landuse (e.g., condition based on the width and continuity of vegetated buffers and the extent of impervious surfaces and area of non-forest cover in the drainage basin);
-
(3)
hydrologic regime (e.g., condition based on the presence and intensity of hydrologic alterations, including the intensity of channelization or channel incision determined by channel metrics, extent of floodplain drainage due to artificial drainage channels, height of constructed levees, and area of wetland fill or excavation);
-
(4)
soil and substrate (e.g., condition based on soil properties, including the presence and intensity of soil compaction and conversion to non-hydric soils due to drainage);
-
(5)
detrital habitat (e.g., condition based on the prevalence and size distribution of snags and down dead wood).
Old forest (> 75 years old) stands that had none of the above alterations were assigned a BPJ condition score of 1. Stands > 75 years old with only a minor alteration relative to one of the above parameters and unaltered stands 50–75 years old (mature) stands were assigned a BPJ score of 2. Stands assigned BPJ scores of 1 or 2 were considered to be relatively unaltered and potentially useful for examining the variation in canopy composition among stands. Stands with moderate to major alterations and stands younger than 50 years were assigned BPJ scores ranging from 3 to 5, and were not used in this study. Additional quantitative data were examined for each stand to further cull stands. For example, BPJ-2 stands were included only if the mean diameter of the nine largest trees in the sample plots was > 30 cm DBH (diameter at 1.5 m). Using the above criteria, 157 of an initial 340 stands sampled by co-authors Rheinhardt, Wilder, and Williams met the criteria for inclusion. Data from an additional 68 unaltered (BPJ-1) stands were added to the data set, derived from similarly sampled stands by Wilder and Roberts (2002), Klimas et al. (2005), Noble et al. (2007), and Williams et al. (2010). Of the 225 stands used, 120 were BPJ-1 and 105 were BPJ-2.
All stand locations were recorded with a handheld GPS and identified according to MLRA region. Through 2009, canopy trees in each stand were sampled in three fixed, circular plots 11.3-m in radius (0.04 ha). Using this fixed-plot method, tree species and DBH were recorded for every stem > 15 cm DBH within the circular plots. After 2009, the combined Bitterlich-rangefinder-circular quadrat method (sensu Levy and Walker 1971) was used to provide comparable data with less sampling effort. With the Bitterlich method, an angle gauge (Relaskop, prism, or Bitterlich stick) was used to tally stems > 15 cm DBH, by species, while rotating the angle gauge 360° around a given point. The number of canopy trees tallied using an angle gauge was converted to basal area by multiplying the basal area factor (BAF) of the gauge, usually 2 (BAF-2 m), by the number of trees tallied. Tree density, by species, was then obtained by counting stems > 15 cm DBH within a 10-m radius circle (0.0314 ha) centered on the Bitterlich point. Only trees > 15 cm DBH were included in tallies because we had previously determined from extensive field work that almost all stems ≤ 15 cm DBH were in the understory of mature forest stands (i.e., not in the canopy).
Data Analysis
For each stand, basal area of canopy trees (i.e., stems > 15 cm DBH) were converted to relative basal area (RBA), by species. Likewise, the number of canopy trees counted in each circular plot was converted to density (stems/ha) and relativized, by species. Relative basal area and relative density were then averaged to obtain an Importance Value (IV) (maximum IV = 100) for each species. Data were analyzed using PC-ORD software (McCune and Mefford 2011), including applying a Non-metric Multidimensional Scaling (NMS) ordination to the data. Before ordinating, tree species were deleted from the grand data matrix (of all subclasses) if they did not occur in more than 5 % of stands in any of one the three subclasses. This elimination of rare species was performed in order to reduce noise and enhance the detection of ordination structure (McCune and Grace 2002). After species were removed, the data matrices were re-relativized for depiction in tables, so that total stand IV again equaled 100 %. Quantitative, explanatory variables and species data were put in a second matrix, which PC-ORD used to construct joint-plot vectors to show the association between the explanatory variables and species with the ordination axes. Potential explanatory variables for each stand included mean DBH of three largest trees in each plot, stand basal area of all canopy trees, stand density of all canopy trees, stand basal area and density of all midstory trees (stems 10–15 cm DBH), and IV for every species in the ordination. BPJ was also used as a coding variable to determine if there was any relationship between BPJ and the ordination position of stands.
NMS was performed on all stands combined to graphically represent the variation among HGM subclasses, and then on each subclass separately to show the variation among MLRA regions within a subclass. Before ordinating by subclass, rare species were deleted from the data matrix, i.e., those with frequency < 5 % in the subclass. Each NMS analysis was initially run in autopilot mode to determine the optimal number of axes to fit and the best starting configuration. The ordination was then re-run manually with the recommended number of axes and starting configuration to obtain the final solution, using 500 iterations and a stability criterion of 10-6. Final stress values ranged from 16.4 to 18.1, with 3-dimensional solutions identified as optimal in all cases. A joint-plot cutoff threshold value (r2) of 0.30 was used to identify quantitative, explanatory variables or species IV values that were correlated with the ordination axes. Values that met the cutoff threshold were depicted as vectors (arrows) originating at the centroid of each ordination diagram; however, only some species IV values met the threshold, whereas the explanatory variables did not. Ordinations were then rotated until the strongest (longest) vector was parallel to Axis 1 of the ordination diagram.
Multi-response permutation procedure (MRPP) in PC-ORD was used to test for differences in canopy composition among HGM subclasses, with rank-transformed relative Sorensen distances used as the dissimilarity measure. Within each HGM subclass, MRPP analyses were also used to test for differences in composition among MLRA regions. Contrasting comparisons were run by grouping subclass pairs that were not found to be dissimilar (sensu De Cáceres et al. 2010), but grouping was only warranted in one instance.
Results
In the 225 riverine stands sampled, 43 canopy species were recorded with frequency of occurrence > 5 % within any of the three subclasses (Table 2, Supplementary material Table A1). Although varying in relative importance among subclasses, five species tended to dominate or co-dominate stands in at least one of the three HGM subclasses, i.e., they had an IV > 10 in more than 25 % of stands within a subclass: red maple (Acer rubrum), swamp blackgum (Nyssa biflora), sweetgum (Liquidambar styraciflua), sweetbay (Magnolia virginiana), and overcup oak (Q. lyrata). In addition to these five species, tulip poplar (Liriodendron tulipifera) co-dominated mid-gradient stands in the SCP region, laurel oak co-dominated all subclasses in the ACF, water tupelo (Nyssa aquatica) co-dominated mid-gradient stands in the ACF, and Q. nigra co-dominated mid-gradient stands in the MVL. A portion of the remaining 34 canopy species, although widespread and not consistently abundant, sometimes showed high dominance in individual stands. For example, various oaks were locally important: swamp chestnut oak (Q. michauxii), willow oak (Q. phellos), cherrybark oak (Q. pagoda), and Nuttall oak (Q. texana), as were ash species (Fraxinus spp.), bald cypress (Taxodium distichum), American elm (Ulmus americana), sycamore (Platanus occidentalis), loblolly pine (Pinus taeda), and slash pine (P. elliottii). These species, particularly where abundant, are likely responsible for some of the variation in composition revealed by the ordination.
MRPP indicated that the three subclasses supported canopy compositions that significantly differed from one another (p < 0.0000003, t = −26.25). In the NMS ordination (Fig. 2), headwater stands differed from low-gradient stands on Axis 2, with mid-gradient stands intermediate in composition between the two and differentiated along Axis 1. Based on joint-plot vectors (Fig. 2), swamp blackgum (Nb) and sweetbay (Mv) were more important in headwater stands, while sweetgum (Ls) was more important in the mid- and low-gradient stands differentiated by Axis 1.

NMS ordination of canopy composition in three riverine HGM subclasses across the Atlantic and Gulf Coastal Plains. Joint-plot vectors (arrows), show the direction and relative correlation strength between species IV values and the ordination axes, relative to a threshold of r 2 = 0.30. Tick mark intervals represent 12.5 % of the axis range, which were relativized to the maximum ordination score. Abbreviations: Hw headwater complex, Mg mid-gradient floodplains, Lg low-gradient floodplains, Mv Magnolia virginiana, Nb Nyssa biflora, Ls Liquidambar styraciflua
Given that the three HGM subclasses differed in composition, MRPP and NMS were run on each subclass to examine the variation among MLRA regions (Fig. 3) and between BPJ scores (1 vs. 2). Headwater stands sampled in three MLRA regions (SCP, ACF, and WCP) differed significantly from one another (MRPP, p < 0.00015, t = -19.18), but not by BPJ score (p = 0.478, t = 0.155). In the NMS ordination, headwater stands in the ACF region were strongly differentiated from stands in the WCP region on Axis 2 (Fig. 3a), with SCP stands tending to separate along Axis 1. The associated joint plot showed that sweetbay (Mv) had greater importance in WCP stands, whereas swamp blackgum (Nb) and sweetgum (Ls) varied negatively with one another relative to the distribution of stands along Axis 1.

NMS ordinations of canopy composition in each HGM subclass, coded by MLRA region (see Table 1). a Headwater complexes. b Mid-gradient floodplains. c Low-gradient floodplains. MLRA abbreviations: SCP Southern Coastal Plain, ACF Atlantic Coast Flatwoods, WCP Western Coastal Plain. Joint-plot vectors and species abbreviations as in Fig. 2, plus Ar = Acer rubrum
For mid-gradient stands, the same three MLRA regions differed significantly from one another (MRPP, p < 0.00046, t = -11.09), but not by BPJ score (p = 0.065, t = -1.654). The NMS ordination (Fig. 3b) differentiated SCP from WCP stands on Axis 2, with separation of less-numerous ACF stands along Axis 1. The joint-plot vectors suggested that red maple (Ar) had greater importance in SCP stands, while sweetgum (Ls) had greater importance in both SCP and WCP stands and less importance in ACF stands.
Analysis of low-gradient stands encompassed four sub-regions. An initial MRPP analysis did not distinguish composition between the SCP (n = 28) and ACF (n = 7) stands (p = 0.47, t = 0.07), so the two regions were grouped for subsequent tests. The grouped SCP/ACF region differed from the WCP and MVL regions (p < 0.0057, t = -14.2) and by BPJ score (p = 0.0003, t = -5.492). In the NMS ordination (Fig. 3c), the SCP/AFC group was differentiated from WCP stands, with less clear separation of MVL stands. The joint-plot vector suggested that sweetgum (Ls) had greater importance in WCP and MVL stands on the right end of Axis 1.
Discussion
This study comprises the largest compilation of quantitative data describing the canopy composition of riverine forests across the southeastern U.S. Coastal Plain. Our conclusion that the forest canopy composition varies with HGM riverine subclass, even across such a spatially expansive physiographic region, supports the premise that hydrogeomorphology is an important factor controlling ecological processes (sensu Brinson 1993a). To our knowledge, no prior studies have explicitly tested the extent to which various HGM subclasses differ in species composition over such an extensive geographic region, or to what extent composition differs within an HGM subclass among sub-regions within a particular physiographic province. To the extent that HGM subclass and MLRA region represent variations in climate, hydrologic regime, and soils, we can infer how these differences might be related to the compositional patterns we observed.
The stream networks we sampled range from groundwater-driven headwater complexes, where surficial soils tend to be saturated for much of the year, to overbank-flow dominated floodplains of low-gradient (> 6th order) rivers, where floodplains tend to flood for long durations in the spring, but are dry throughout much of the remaining growing season. These hydrogeomorphically-mediated differences can explain the separation of headwater (1st to 3rd order) and low-gradient stands in the full ordination of Coastal Plain stands. The intermediate ordination positions of mid-gradient (4th–6th order) stands may reflect their intermediate hydrologic status, i.e., by the effects of groundwater saturation and shorter-duration overbank flooding regime. These differences in hydrologic regime are likely a primary reason for the variation among HGM types across the broader Coastal Plain region. Overlap among HGM types in composition likely reflects overlap in hydrologic regime among hydrogeomorphic classes, i.e., hydrologic regime varies within all three HGM types, but geomorphology restricts the extent of this variation.
Within an HGM subclass, the variation among MLRA regions may reflect the criteria used to classify the MLRA regions: soils, climate, and potential vegetation. One might expect a wide range of edaphic variation to occur across such an extensive physiographic region and composition within an HGM type did vary relative to regional differences. Even so, stands within a given HGM type were remarkably similar in composition relative to other HGM types.
For the most part, our results of canopy species affiliations within hydrogeomorphically different reaches of a stream network both support and build upon prior studies. For example, although laurel oak occurs throughout the Coastal Plain from Virginia to East Texas (Little 1977; Burns and Honkala 1990), the available literature has not generally indicated that the species is more prevalent in the Atlantic Coast Flatwoods region (where it co-dominated in all HGM subclasses) than elsewhere in the Southeast. In fact, we found that laurel oak was relatively unimportant in other MLRAs of the Coastal Plain. In contrast to laurel oak, we found that some species commonly co-dominated only particular subclasses and/or regions. For example, sweetbay never co-dominated stands in overbank-flow dominated floodplains (mid- and low-gradient subclasses), but it commonly co-dominated headwater stands in all MLRA regions, except the Flatwoods region. Similarly, overcup oak was a common co-dominant (IV > 10) only in low-gradient stands in the Western Coastal Plain, rarely co-dominated stands in other mid- and low-gradient stands in other regions, and never co-dominated stands in the headwater subclass of any MLRA region (Supplementary material Table A1). Likewise, tulip poplar occasionally co-dominated headwater stands of the Atlantic Coast Flatwoods and headwater and mid-gradient stands of the Southern Coastal Plain; however, it rarely or never co-dominated stands elsewhere (Supplementary material Table A1).
Swamp blackgum (N. biflora) is common in Southeastern swamps and often overwhelmingly dominates stands with long hydroperiods. It is generally considered to be important in shallow backswamps of alluvial forests and in non-alluvial depressions and flats (Burns and Honkala 1990). In this study, swamp blackgum was important in mid-gradient reaches of the Atlantic Flatwoods and Western Coastal Plain regions and particularly prevalent (and important) in headwater reaches of all Coastal Plain regions. Locations of stands were not randomly chosen, but neither were blackgum stands preferentially sought for sampling. Thus, it is possible that swamp blackgum is more important in headwater reaches than is generally recognized.
Water tupelo (N. aquatica) is associated with even longer hydroperiods than swamp blackgum (Hook and Brown 1973; Burns and Honkala 1990). The species is generally associated with backswamps and abandoned channels of large river systems. However, although water tupelo was more prevalent in low-gradient stands (21 %) in this study, it occurred in all three subclasses (Supplementary material Table A1), often as an overwhelming dominant. Again, although stand locations were not randomly chosen, our results suggest that water tupelo may be more prevalent in headwater and mid-gradient reaches than generally acknowledged.
There were complementary advantages and limitations to the analytical approaches used in the study. Mean IVs (Table 2) provided a characterization of central tendencies, but could not adequately describe the more complex underlying variation in IV among all species (see Supplementary material Table A1). MRPP tested whether pre-defined groupings exhibited major compositional differences, which to some extent obscures the overlapping variation that might be expected along continuous gradients. In contrast, the NMS ordinations were useful for visualizing the more continuous relative similarities and dissimilarities within stream networks and among geographic regions. Taken together, these tools were useful in characterizing the relationships in canopy composition among floodplains in stream networks in the Coastal Plain.
Management Applications
Because the canopy stratum comprises > 90 % of the biomass of a floodplain forest, it is not unreasonable to presume that canopy species composition is important to the functioning of floodplain forests, particularly biogeochemical and habit functioning. The canopy composition of mature forests also integrates environmental conditions that have occurred over long time periods. For these reasons, canopy composition and forest maturity are often identified as potentially important indicators of forest health, productivity, and habitat potential. This suggests that a quantitative measure of canopy composition would be useful in reference-based assessments such as the HGM approach (Brinson 2009; Smith et al. 1995), where site condition is determined relative to a range of reference wetland conditions exhibited by a regional subclass. In this study, data on the canopy component of mature and older floodplain forests (> 50 y old) were useful for quantifying the variation in composition across a large geographic area and for defining floristic standards that could be useful in evaluating ecosystem condition.
The geographic ranges for species associated with HGM subclass and MLRA region should be taken into account when evaluating forest condition in HGM assessments. Likewise, species that are widespread, but only locally important, should also be given consideration when determining reference conditions. For example, bald cypress is locally abundant where hydrologic conditions are suitable. Similarly, depending on local environmental conditions, overcup oak, willow oak, and swamp chestnut oak are sometimes locally abundant in the canopy. The overlap exhibited by stands in the ordination diagrams shows how widely canopy composition varies across the Coastal Plain, even within a subclass and MLRA region, and so any evaluation of condition based on floristic reference conditions must be flexible enough to integrate this variability.
In this study, nine canopy species could be associated as potential co-dominant species for particular subclasses and/or MLRA regions. The relative importance of these species could be used to help characterize floristic condition, particularly when setting standards for regional subtypes within a hydrogeomorphic subclass. For example, laurel oak would not be useful for defining standards in the Western Coastal Plain region because it was not found in any of the stands sampled there. Likewise, sweetbay might not be an appropriate indicator in headwater reaches of the Atlantic Flatwoods region (except for pocosin drainages) because it rarely co-dominated there. Nonetheless, results of this study could be used to customize standards for evaluating canopy floristic condition by MLRA region and HGM subclass, as long as potential variations in composition are integrated. In designing restoration plans, the entire suite of species identified in this study, relative to HGM subclass and MLRA region, should be considered when planning floodplain forest restoration projects.
References
Bedford BL, Walbridge MR, Aldous A (1999) Patterns in nutrient availability and plant diversity of temperate North American wetlands. Ecology 80:2151–2169
Bledsoe B (1993) Vegetation along hydrologic and edaphic gradients in a North Carolina coastal plain creek bottom. North Carolina State University, Raleigh, M.S. Thesis
Brinson MM (1990) Riverine forests. In: Lugo A, Brinson M, Brown S (eds) Forested wetlands: ecosystems of the world, 15. Elsevier, New York, pp 87–141
Brinson MM (1993a) Changes in the functioning of wetlands along environmental gradients. Wetlands 13:65–74
Brinson, MM (1993b) A hydrogeomorphic classification for wetlands. Technical Report RP-DE-4, Vicksburg, MS, USA. Available via http://el.erdc.usace.army.mil/wetlands/procedure.html
Brinson MM (2009) The United States HGM (Hydrogeomorphic) approach. In: Maltby E, Barker T (eds) The wetlands handbook. Wiley-Blackwell, London, Oxford, pp 486–512
Brinson MM, Miller KH, Rheinhardt R, Christian R, Meyer G, O’Neal J (2006) Developing reference data to identify and calibrate indicators of riparian ecosystem condition in rural coastal plain landscapes in North Carolina. Report to the Ecosystem Enhancement Program, North Carolina Department of Environment and Natural Resources. Raleigh, NC, USA. (http://www.nceep.net/pages/resources.htm)
Brooks RP, Brinson MM, Havens K, Hershner C, Rheinhardt RD, Wardrop DH, Whigham DF, Jacobs AD, Rubbo JM (2011) Proposed hydrogeomorphic classification for wetlands of the Mid-Atlantic region, USA. Wetlands 31:207–219
Burns RM, Honkala BH (1990) Silvics of North America: 2. Hardwoods. Agriculture Handbook 654. U.S. Department of Agriculture, Forest Service, Washington, DC, USA
De Cáceres M, Legendre P, Moretti M (2010) Improving indicator species analysis by combining groups of sites. Oikos 119:1674–1684
Denslow JS, Battaglia LL (2002) Stand composition and structure across a changing hydrologie gradient: Jean Lafitte National Park. Wetlands 22:738–752
Dickson JG, Noble RE (1978) Vertical distribution of birds in a Louisiana bottomland hardwood forest. Wilson Bulletin 90:19–30
Gemborys SR, Hodgkins EJ (1971) Forests of small stream bottoms in the coastal plain of southwestern Alabama. Ecology 52:70–84
Glascock S, Ware SA (1979) Forests of small stream bottoms in the Peninsula of Virginia. Virginia Journal of Science 30:17–21
Hook D, Brown CL (1973) Root adaptations and relative flood tolerance of five hardwood species. Forest Science 19:225–229
Hope D, Billett MF, Cresser MS (1994) A review of the export of carbon in river water: fluxes and processes. Environmental Pollution 84:301–324
Klimas CV, Murray EO, Pagan J, Langston H, Foti T (2005) A regional guidebook for applying the hydrogeomorphic approach to assessing wetland functions of forested wetlands in the west Gulf Coastal Plain region of Arkansas. ERDC/EL TR-05-12, U.S. Army Engineer Research and Development Center, Ecosystem Management and Restoration Research Program, Vicksburg, MS, USA. Available via http://el.erdc.usace.army.mil/wetlands/guidebooks.cfm
Laiho R, Prescott CE (1999) The contribution of coarse woody debris to carbon, nitrogen and phosphorus cycles in three Rocky Mountain coniferous forests. Canadian Journal of Forest Research 29:1592–1603
Levy GF, Walker SW (1971) The combined Bitterlich-rangefinder-circular quadrat method in phytosociological studies. Jeffersonia 5:37–39
Little EL, Jr. (1977) Atlas of United States trees, volume 4, minor Eastern hardwoods. U.S. Department of Agriculture Publication 1342. http://esp.cr.usgs.gov/data/atlas/little/querlaur.pdf
Loehle C (2003) Competitive displacement of trees in response to environmental change or introduction of exotics. Environmental Management 32:106–115
Martin TL, Kaushik NK, Trevors JT, Whiteley HR (1999) Review: denitrification in temperate climate riparian zones. Water, Air, and Soil Pollution 111:171–186
McCune B, Grace JB (2002) Analysis of ecological communitries. MjM Software Design, Gleneden Beach, Oregon, USA
McCune B, Mefford MJ (2011) PC-ORD: multivariate analysis of ecological data. MjM Software. Version 6. Gleneden Beach, Oregon, USA
Monk CD (1966) An ecological study of hardwood swamps in north-central Florida. Ecology 47:649–654
Noble CV, Wakeley JS, Roberts TH, Henderson C (2007) Regional guidebook for applying the hydrogeomorphic approach to assessing the functions of headwater slope wetlands on the Mississippi and Alabama Coastal Plains. ERDC/EL TR-07-9, U.S. Army Engineer Research and Development Center, Vicksburg, MS, USA
Odum EP (1969) The strategy of ecosystem development. Science 164:262–270
Parker GG (1995) Structure and microclimate of forest canopies. In: Lowman MD, Nadkarni NM (eds) Forest canopies: a review of research on a biological frontier. Academic, San Diego, pp 73–106
Parsons SE, Ware SA (1982) Edaphic factors and vegetation in Virginia Coastal Plain swamps. Bulletin of the Torrey Botanical Club 109:365–370
Prescott CE (2002) The influence of the forest canopy on nutrient cycling. Tree Physiology 22:1193–1200
Rheinhardt RD, Rheinhardt MC, Brinson MM, Faser K (1998) Forested wetlands of low order streams in the inner coastal plain of North Carolina, USA. Wetlands 18:365–378
Rheinhardt RD, Rheinhardt MC, Brinson MM, Faser K (1999) Application of reference data for assessing and restoring headwater ecosystems. Restoration Ecology 7:241–251
Rheinhardt RD, Whigham KH, Brinson MM (2000) Floodplain vegetation of headwater streams in the inner coastal plain of Virginia and Maryland. Castanea 65:21–35
Rheinhardt RD, McKenney-Easterling M, Brinson MM, Rubbo J, Brooks R, Whigham D, O’Brien D, Hite J, Armstrong A (2007) Canopy composition and forest structure as indicators of habitat condition in headwater riparian ecosystems. Restoration Ecology 17:51–59
Rheinhardt R, Brinson M, Miller K, Myer G (2012) Carbon storage of headwater riparian zones in an agricultural landscape. Carbon Balance and Management. Available via http://www.cbmjournal.com/content/7/1/4
Robinson SK, Holmes RT (1984) Effects of plant species and foliage structure on the foraging behavior of forest birds. Auk 101:672–684
Rodewald AD, Abrams MD (2002) Floristics and avian community structure: implications for regional changes. Forest Science 48:267–272
Sharitz RR, Mitsch WJ (1993) Southern floodplain forests. In: Martin WH, Boyce SG (eds) Biodiversity of the southeastern United States: lowland terrestrial communities. John Wiley and Sons, New York, pp 311–372
Smith RD, Ammann A, Bartoldus C, Brinson MM (1995) An approach for assessing wetland functions using hydrogeomorphic classification, reference wetlands, and functional indices. Technical Report WRP-DE-9, U.S. Army Engineer Waterways Experiment Station,, Vicksburg, MS, USA
Strahler AN (1952) Dynamic basis of geomorphology. Geological Society of America Bulletin 63:923
Tiner RW (2011) Dichotomous Keys and Mapping Codes for Wetland Landscape Position, Landform, Water Flow Path, and Waterbody Type Descriptors: Version 2.0. U.S. Fish and Wildlife Service, National Wetlands Inventory Program, Northeast Region, Hadley, MA. 51 pp. Available via http://digitalmedia.fws.gov/cdm/singleitem/collection/document/id/1324/rec/19
USDA (2006) MLRA Geographic Database, version 4.2. Land Resource Regions and Major Land Resource Areas of the United States, the Caribbean, and the Pacific Basin. United States Department of Agriculture Handbook 296. Available via ftp://ftp-fc.sc.egov.usda.gov/NSSC/Ag_Handbook_296/Handbook_296_low.pdf
Warren DR, Bernhardt ES, Hall RO, Likens GE (2007) Forest age, wood and nutrient dynamics in headwater streams of the Hubbard Brook Experimental Forest, NH. Surface Processes and Landforms 32:1154–1163
Wharton CH, Kitchens WM, Pendleton EC, Sipe TW (1982) The ecology of bottomland hardwood swamps of the Southeast: a community profile. FWS/OBS-81/37
Whelan CJ (2001) Foliage structure influences foraging of insectivorous forest birds: an experimental study. Ecology 82:219–231
Wigley TB, Roberts TH (1994) A review of wildlife changes in southern bottomland hardwoods due to forest management practices. Wetlands 14:41–48
Wilder TC, Roberts TH (2002) A regional guidebook for applying the hydrogeomorphic approach to assessing wetland functions of low-gradient riverine wetlands in western Tennessee. ERDC/EL TR-02-6, U.S. Army Engineer Research and Development Center, Vicksburg, MS
Wilder TC, Rheinhardt RD, Noble CV (2013) A regional guidebook for applying the hydrogeomorphic approach to assessing wetland functions of forested wetlands in alluvial valleys of the Coastal Plain of the Southeastern United States. ERDC/EL TR-13-1. Vicksburg, MS: U.S. Army Engineer Research and Development Center, Vicksburg, MS
Williams H, Miller, AJ, McNamee RS, and Klimas CV (2010) A regional guidebook for applying the Hydrogeomorphic Approach to the functional assessment of forested wetlands in alluvial valleys of east Texas, ERDC/EL TR-10-17, U.S. Army Engineer Research and Development Center, Vicksburg, MS. Available via
Wipfli MS, Richardson JS, Naiman RJ (2007) Ecological linkages between headwaters and downstream ecosystems: transport of organic matter, invertebrates, and wood down headwater channels. Journal of the American Water Resources Association 43:72–85
Acknowledgments
Funding was provided by the U.S. Army Corps of Engineers, Engineer Research and Development Center (ERDC). Field data were collected by many individuals besides the authors, Elizabeth Murray in Arkansas, Jill Clancy in Alabama, Keith Daily, Darinda Dans, Levi Gibson, Penny Gibson, Rachael McNamee Erwin, and Adam Miller in Texas and Louisiana. Identification of data to be collected and methods of collection were jointly decided upon by the authors and a panel of wetland experts, including Mark Brinson (East Carolina University), Tom Roberts (Tennessee Technological University), Tad Zebryk (U.S. Corps of Engineers-Mobile District), Richard Darden (U.S. Corps of Engineers-Charleston District), Les Parker (U.S. Corps of Engineers-Charleston District), Mike Zeman (U.S. Natural Resources Conservation Service-Tennessee), Bill Ainslie (U.S. Environmental Protection Agency-Atlanta), Dave Lekson (U.S. Corps of Engineers-Wilmington District), and Justin Hammonds (U.S. Corps of Engineers-Savannah District). Thanks are also extended to two anonymous reviewers who helped improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(XLS 169 kb)
Rights and permissions
About this article
Cite this article
Rheinhardt, R., Wilder, T., Williams, H. et al. Variation in Forest Canopy Composition of Riparian Networks from Headwaters to Large River Floodplains in the Southeast Coastal Plain, USA. Wetlands 33, 1117–1126 (2013). https://doi.org/10.1007/s13157-013-0467-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-013-0467-0