
Abstract
This study was designed to examine the effects of intracerebroventricular injection of DL-AP5 (N-methyl-D-aspartate (NMDA) receptor antagonist) and glutamate on ghrelin-induced feeding behavior in 3-h food-deprived (FD3) broiler cockerels. At first, guide cannula was surgically implanted in the right lateral ventricle of chickens. In experiment 1, birds were intracerebroventricularly injected with 0, 2.5, 5, and 10 nmol of DL-AP5. In experiment 2, chickens received 5 nmol DL-AP5 prior to the injection of 0.6 nmol ghrelin. In experiment 3, birds were administered with 0.6 nmol ghrelin after 300 nmol glutamate, and the cumulative feed intake was determined at 3-h postinjection. The results of this study showed that the intracerebroventricular injection of DL-AP5 increased food consumption in FD3 broiler cockerels (P ≤ 0.05), and this increase occurs in a dose-dependent manner. Moreover, the decreased food intake induced with the intracerebroventricular injection of ghrelin was additively enhanced by pretreatment with glutamate, and this effect was attenuated by DL-AP5 administration(P ≤ 0.05).These results suggest that there is an interaction between ghrelin and glutamatergic system (through NMDA receptor) on food intake in broiler cockerels.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The appetite regulatory network is modulated at the hypothalamic level by the interaction of hormonal and neuronal signaling [7]. Aminergic neurotransmitters have long been implicated in feeding control at the hypothalamic level, and it has previously been shown that peptide hormones such as Leptin, Orexins A and B, and ghrelin differently modulate hypothalamic dopamine, norepinephrine, and serotonin release, which could partially explain their appetite regulatory activity [15].
Ghrelin, an acylated peptide originally identified in rat stomach as the endogenous ligand for the growth hormone secretagogue receptor (GHS-R), regulates growth hormone secretion, food intake, and energy homeostasis [11]. It is known that, in rats, central and peripheral ghrelin increases food intake mainly through activation of neuropeptide Y and agouti-related protein [3, 24]. In chicken, ghrelin messenger RNA (mRNA) expression is detected at the highest level in the proventriculus [6]. Systematic and intracerebroventricular administration of ghrelin or other growth hormone secretagogues like GHRP-2 strongly inhibit food intake in a dose-dependent manner in chickens [9, 19]. This effect of ghrelin is contrary to that of mammals. The mechanism underlying the ghrelin-induced hypophagia in birds is still unclear, although the anorexic effect of ghrelin was reported to be mediated by corticotropin-releasing factor but not neuropeptide Y [19].
Glutamate, a major excitatory neurotransmitter in the central nervous system (CNS), has been suggested to be an endogenous agent involved in the neural control of food intake and body weight in mammals [14]. Systematic, intracerebroventricular, or local administration of glutamate or its agonists to the lateral hypothalamus can evoke dose-related stimulation of food intake [21] that can be mediated by N-methyl-D-aspartate (NMDA) receptor subtypes. Glutamate receptors have been found to be widely distributed in the avian CNS [16], and can be involved in learning and memory processes as well as neuroendocrine control mechanism in chicken [18]. In opposition to mammals, it has been shown that intracerebroventricular injection of glutamate in 24-h food-deprived pigeons and broiler cockerels decreases food intake in a dose-dependent manner [28, 29].
On the basis of these findings and considering the same effects of ghrelin and glutamate on feeding behavior in birds, we hypothesized that glutamatergic system possibly mediates ghrelin signaling in the hypothalamus of birds. Thus, the present study was designed for the first time to investigate whether blocking or stimulating glutamate NMDA receptor can influence ghrelin-induced feeding response in 3-h food-deprived broiler cockerels.
Materials and methods
Animals
Broilers cockerels (Iran-Germany Co., Iran) were reared in heated batteries with continuous lighting until 3 weeks of age. Birds were provided with a mash diet (21% protein and 2,869 kcal/kg of metabolic energy) and water ad libitum. At approximately 2 weeks of age, the birds were transferred to individual cages. The temperature and relative humidity of the animal room was maintained at 22 ± 1°C and 50%, respectively, in addition to the continuous lighting condition.
Drugs
Drugs used in the experiment included DL-AP5 (NMDA receptor antagonist), glutamate, and ghrelin which were purchased from Tocris Cookson Co. (Bristol, UK) and diluted in pyrogen-free 0.9% NaCl solution (saline) that served as control. Doses of drugs were chosen on the basis of preliminary experiments and our previous studies [22, 28].
Surgical preparation
At 3 weeks of age, broilers were anesthetized with Sodium Pentobarbital (Sagatal, Rhone Merieux; 25 mg/kg body weight, IV), and a 23-gauge thin-walled stainless steel guide cannula was stereotaxically implanted into the right lateral ventricle, as described by Denbow et al. [4]. The stereotaxic coordinates were: AP = 6.7, L = 0.7, H = 3.5–4 mm below the dura mater with the head oriented as described by Van Tienhoven and Juhaz [27]. The cannula was secured with three stainless steel screws placed in the calvaria surrounding each guide cannula, followed by acrylic dental cement (Pars acryl) being applied to both the screws and guide cannula. An orthodontic #014 wire (American Orthodontics) trimmed to the exact length of the guide cannula and was inserted into it while the chicks were not being used for the experiments. Lincospectin (Razak) was applied to incision to prevent infection. The birds were allowed a minimum of 5 days recovery prior to injection.
Experimental procedures
To determine the involvement of glutamate NMDA receptor in the brain in ghrelin-induced eating response, effects of centrally administered DL-AP5 and glutamate on ghrelin-induced eating response were determined in chickens. Injections were made with a 29-gauge, thin-walled stainless-steel injection cannula which extends 1.0 mm beyond the guide cannula. This injection cannula was connected to a 10-μl Hamilton syringe connected to a 60 cm length of PE-20 tubing. Solutions were injected over a period of 60 s. Another 60-s period was allowed to permit the solution to diffuse from the tip of the cannula into the ventricle. All experimental procedures were performed between 9 a.m. and 2 p.m. Before the injections, the birds were removed from their individual cages, restricted by hand, and after injections were put back into their cages. Birds were handled and mock-injected daily during the 5 days recovery period to habituate them to the injection procedure. Three hours before the beginning of the experiments, animals were deprived of food but with water ad lib. Immediately after injections, the birds were returned to their cages. Fresh food was supplied at the time of injection, and cumulative feed intake (grams) was recorded at 15, 30, 60, 120, and 180 min after injection. Placement of the guide cannula into the ventricle was verified by the presence of cerebrospinal fluid and intracerebroventricular injection of methylene blue and anatomically slicing the frozen brain tissue at the end of the experiments.
Experiment 1 was designed to examine the effect of intracerebroventricular injections of different DL-AP5 doses on cumulative feed intake in 3-h food-deprived (FD3, n = 8 for each group) chickens. For this purpose, the birds received 0, 2.5, 5, and 10 nmol of DL-AP5 in a volume of 10 μl. Control group was injected with 10 μl of 0.9% NaCl solution.
In experiment 2, birds of each group received two injections. The first injection consisted of either 0 or 5 nmol DL-AP5 in a volume of 5 μl. The second injection consisted of either 0 or 0.6 nmol ghrelin in a volume of 5 μl, 15 min after the first injection as described in Table 1 (n = 7–9 for each group).
Experiment 3 was conducted similarly to the first experiment except that the chicks were injected with 0 or 300 nmol glutamate instead of DL-AP5.
The dosages of drugs were selected based on our previous studies [22, 28] and preliminary experiments. During the injection of higher doses of ghrelin and glutamate, the birds were very active, excited, vocal, and sometimes convulsive.
Statistical analysis
Cumulative feed intake is presented as mean ± SE and analyzed using a one-way analysis of variance (ANOVA) at each time period. For treatment showing a main effect by ANOVA, means have been compared by post hoc Tukey test. The values were considered significantly different when P ≤ 0.05.
Results
The food intake response to intracerebroventricular injection of DL-AP5, glutamate, and ghrelin in broiler chickens is presented in Figs. 1, 2, and 3.

Effect of intracerebroventricular injection of different doses of AP-5 on cumulative feed intake in 3-h food-deprived chickens. Data are presented as mean ± SE; a indicates significant difference from control group (saline) within each time point (P ≤ 0.05)

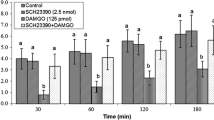
Effect of intracerebroventricular injection of AP-5 (5 nmol) followed by ghrelin (0.6 nmol) on cumulative feed intake in 3-h food-deprived chickens. Data are presented as mean ± SE; a indicates significant difference from control group (saline + saline) within each time point (P ≤ 0.05) and b indicates significant difference from ghrelin group (saline + ghrelin) within each time point (P ≤ 0.05)

Effect of intracerebroventricular injection of glutamate (300 nmol) followed by ghrelin (0.6 nmol) on cumulative feed intake in 3-h food-deprived chickens. Data are presented as mean ± SE; a indicates significant difference from control group (saline + saline) within each time point (P ≤ 0.05) and b indicates significant difference from ghrelin group (saline + ghrelin) within each time point (P ≤ 0.05)
In experiment 1, the injected DL-AP5 into the lateral ventricle of 3-h food-deprived chickens caused a dose-dependent increase in food consumption which was significant for 5 and 10 nmol doses (Fig. 1; f(3,25) = 12.43, P ≤ 0.05). The 5 nmol DL-AP5 dose was selected for the following experiment because it was found to induce an increase in food intake in FD3 animals without affecting other non-ingestive behavioral parameters.
In experiment 2, intracerebroventricular injection of 0.6-nmol ghrelin alone decreased food intake, and this effect of ghrelin on food intake was significantly attenuated by DL-AP5 pretreatment in 3-h food-deprived chickens until 120 min after injections (Fig. 2; f(3,25) = 14.08, P ≤ 0.05).
The results of experiment 3 showed that the inhibitory effect of ghrelin on cumulative food intake was significantly increased by 300 nmol glutamate pretreatment, probably in an additive manner (Fig. 3; f(3,25) = 18.56, P ≤ 0.05).
Discussion
The present study was designed for the first time to investigate the possible involvement of glutamatergic circuits in ghrelin control mechanisms of feeding behavior in broiler cockerels. The results obtained from experiment 1 (Fig. 1) which show that the blockage of NMDA receptors dose-dependently increases food intake in 3-h food-deprived chickens is in line with earlier findings of Zeni et al. [29] on pigeons. According to that report, pretreatment with MK-801 (NMDA receptor antagonist) before intracerebroventricular injection of glutamate induced a partial suppression of food intake in 24-h food-deprived pigeons. Furthermore, these findings and the results obtained from experiment 3 confirm our previous study which showed that intracerebroventricular infusion of 300 nmol glutamate inhibits food intake in 24-h food-deprived broiler cockerels [28].
The results obtained from experiment 2 are consistent with previous studies reported in which systematic or intracerebroventricular administration of ghrelin or GHRP-2 strongly inhibits food intake in a dose-dependent manner in chickens [9, 19]. However, in mammals, central and peripheral administration of ghrelin increase food intake mainly through activation of neuropeptide Y and agouti-related protein [3, 24]. Such discrepancies may be due to species properties, the affected site in the brain, or 3-h deprivation of food in this experiment. These results indicate that species properties are important in the aminergic feeding regulatory systems.
In experiment 2, the decreased food consumption induced by the intracerebroventricular injection of ghrelin was attenuated by pretreatment with DL-AP5 (Fig. 2). It showed that the inhibitory effect of ghrelin on food intake is modulated by the pathway(s) linked to the NMDA receptor. In this regard, co-administration of glutamate and ghrelin additively enhanced inhibitory effect of ghrelin on food intake (Fig. 3). These results indicate that both glutamate and ghrelin are involved in regulatory centers of feeding and energy balance.
Several lines of evidence suggest that glutamatergic system may be involved in the ghrelin-induced regulation of feeding behavior. Ghrelin has been identified as an endogenous high-affinity ligand for GHS-R, a G-protein-coupled receptor highly expressed in the hypothalamus arcuate nucleus [23]. Additionally, neurons of the arcuate nucleus contain the orexigenic neuropeptide Y and the anorectic pro-opiomelanocortin-derived peptide α-melanocyte stimulating hormone [5]. GHS-R is highly expressed in the arcuate nucleus with 94% of all neuropeptide Y containing neurons and 8% of pro-opiomelanocortin neurons co-expressing GHS-R mRNA [17]. In the mammals, it seems that two major hypothalamic pathways are the predominant mediators of ghrelin’s influence on energy balance [20, 25]; one involves the neuropeptide Y neurons [8], and the other involves the α-melanocyte stimulating hormone-containing neurons [12]. It is well known that the direct ghrelin-induced activation of neuropeptide Y neurons in the ventromedial arcuate nucleus is paralleled by an inhibitory synaptic effect on α-melanocyte stimulating hormone neurons of the ventrolateral arcuate nucleus through an increase of GABAergic neurotransmission [10]. In this regard, our previous study revealed an interaction between glutamate and GABA neurons on feeding behavior in broiler cockerels [28]. We showed that inhibitory effect of glutamate on food intake was significantly amplified with pretreatment of bicuculline (GABAA receptor antagonist), and this effect of glutamate attenuated with muscimol (GABAA receptor agonist).
Formerly, Saito and colleagues showed that inhibitory effect of ghrelin on food consumption is mediated by the corticotropin-releasing factor in neonatal chicks [19]. It has been suggested that glutamate mediates corticotropin-releasing factor release, and studies have shown that direct application of glutamate to paraventricular nucleus resulted in corticotropin-releasing factor release, a rise in ACTH, and an increase in corticosterone levels. This effect of glutamate on corticotropin-releasing factor release is mediated through NMDA receptors, and pretreatment with an NMDA receptor antagonist reduces corticosterone release [13]. In this regard, our finding is consistent with the reports of Saito et al. [19].
It has been revealed that phenotypes of many neurons in the arcuate nucleus are glutamatergic, and glutamate is involved in the innervation of arcuate neurons [26], a key site for the interaction between circulating hormones and the central nervous system [14]. The arcuate nucleus, which has a weak blood–brain barrier [1], contains two neuronal populations which integrate signals of nutritional status and influence energy homeostasis [2]. Moreover, Meister [14] reported that cell bodies in the ventrolateral part of the arcuate nucleus that stimulate feeding behavior contain neuropeptide Y/agouti-related peptide, but are also predominantly GABAergic in nature. Some arcuate neurons in the ventrolateral part of the arcuate nucleus that have an inhibitory influence on food intake are glutamatergic, GABAergic, and/or cholinergic. Neurons of the lateral hypothalamic area containing hypocretin/orexin are to a large extent glutamatergic whereas some lateral hypothalamic MCH neurons appear to be GABAergic. Therefore, it seems that glutamate, GABA and ghrelin may interact with each other to modulate food intake by excitatory or inhibitory effects on orexigenic and anorexigenic neurons of arcuate nucleus in hypothalamus. In this study, we showed that intracerebroventrical co-administration of ghrelin and glutamate amplified the hypophagic response compared with that induced by ghrelin or glutamate individually. These results suggest that ghrelin and glutamate may inhibit neuropeptide Y/agouti-related peptide neurons through GABAA receptor and excite pro-opiomelanocortin neurons through NMDA receptor causing a decrease in food intake in chickens. In addition, DL-AP5 may attenuate ghrelin-induced hypophagia through reduction in release of neuropeptide Y and agouti-related protein.
References
Broadwell RD, Brightman MW (1976) Entry of peroxidase into neurons of the central and peripheral nervous systems from extracerebral and cerebral blood. J Comp Neurol 166(3):257–283
Cone RD, Cowley MA, Butler AA, Fan W, Marks DL, Low MJ (2001) The arcuate nucleus as a conduit for diverse signals relevant to energy homeostasis. Int J Obes Relat Metab Disord 25(5):63–67
Date Y, Murakami N, Toshinai K, Matsukara S, Niijima A, Matsuda Kangawa K, Nakazato M (2002) The role of the gastric afferent vagal nerve in ghrelin-induced feeding and growth hormone secretion in rats. Gastroenterology 123:1120–1128
Denbow DM, Cherry JA, Siegel PB, Van Kery HP (1981) Eating, drinking and temperature response of chicks to brain catecholamine injections. Physiol Behav 27:265–269
Hagan MM, Rushing PA, Schwartz MW, Yagaloff KA, Burn P, Woods SC, Seeley RJ (1999) Role of the CNS melanocortin system in the response to overfeeding. J Neurosci 19:2362–2367
Kaiya H, Vander Geyten S, Kojima M, Hosoda H, Kitajima Y, Matsumoto M, Geelissen S, Darras VM, Kangawa K (2002) Ghrelin: purification cDNA cloning and biological activity. Endocrinol 143:3454–3463
Kalra SP, Dube MG, Pu S, Xu B, Horvath TL, Kalra PS (1999) Interacting appetite-regulating pathways in the hypothalamic regulation of body weight. Endocr Rev 20:68–100
Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H, Wakabayashi I (2001) Chronic central infusion of ghrelin increases hypothalamic neuropeptide Y and agouti-related mRNA levels and body weight in rats. Diabetes 50:2438–2443
Khan MSI, Dodo K, Yahata K, Nishimoto S, Ueda H, Taneike T, Kitazawa T, Hosaka Y, Bungo T (2006) Intracerebroventricular administration of growth hormone releasing peptide-6 ( GHRP-6) inhibits food intake, but not food retention of crop and stomach in neonatal chicks. J Poult Sci 43:35–40
Kiss J, Csaba Z, Csaki A, Halasz B (2005) Glutamatergic innervation of neuropeptide Y and pro-opiomelanocortin-containing neurons in the hypothalamic arcuate nucleus of the rat. Eur J Neurosci 21:2111–2119
Kojima M, Kangawa K (2005) Ghrelin: structure and function. Physiol Rev 85:495–522
Marks DL, Cone RD (2001) Central melanocortins and the regulation of weight during acute and chronic disease. Recent Prog Horm Res 56:359–375
Mathew SJ, Coplan JD, Schoepp DD, Smith EL, Rosenblum LA, Gonman JM (2001) Glutamate–hypothalamic–pituitary–adrenal axis interactions: implications for mood and anxiety disorders. CNS Spectr 6(555–556):561–564
Meister B (2007) Neurotransmitters in key neurons of the hypothalamus that regulate feeding behavior and body weight. Physiol Behav 92:263–271
Orlando G, Brunetti L, Di Nisio C, Michelotto B, Recinella L, Ciabattoni G, Vacca M (2001) Effects of cocaine- and amphetamine-regulated transcript peptide, leptin, and orexins on hypothalamic serotonin release. Eur J Pharmacol 430:269–272
Ottiger HP, Gerin-Moser A, Del Principe F, Dutly F, Streit P (1995) Molecular cloning and differential expression patterns of avian glutamate receptor mRNAs. J Neurochem 64:2413–2426
Reidiger T, Traebert M, Schmid HA, Scheel C, Lutz TA, Scharrer E (2003) Site-specific effects of ghrelin on the neuronal activity in the hypothalamic arcuate nucleus. Neurosci Lett 341:151–155
Riters LV, Bingman VP (1994) The NMDA receptor antagonist MK-801 impairs navigational learning in homing pigeons. Behav Neural Biol 62:50–59
Saito ES, Kaiya H, Tachibana T, Tomonage S, Denbow DM, Kangawa K, Furuse M, Furuse M (2005) Inhibitory effects of ghrelin on food intake is mediated by the corticotropin-releasing-factor system in neonatal chicks. Regul Pept 125:201–208
Shintani M, Ogawa Y, Ebihara K, Aizawa-Abe M, Miyanaga F, Takaya K, Hayashi T, Inoue G, Hosoda K, Kojima M, Kangawa K, Nakao K (2001) Ghrelin, an endogenous growth hormone secretagogue, is a novel orexigenic peptide that antagonizes leptin action through the activation of hypothalamic neuropeptide Y/Y1 receptor pathway. Diabetes 50:227–232
Stanley BG, Butterfield BS, Grewal RS (1997) NMDA receptor coagonist glycine site: evidence for a role in lateral hypothalamic stimulation of feeding. Am J Physiol 273:790–796
Taati M, Nayebzadeh H, Khosravinia H, Cheraghi J (2010) The role of the histaminergic system on the inhibitory effect of ghrelin on food intake in broiler chickens. IJVR 11(1):38–45
Tannenbaum GS, Lapointe M, Beaudet A, Howard AD (1998) Expression of growth hormone secretagogue-receptors by growth hormone-releasing hormone neurons in the mediobasal hypothalamus. Endocrinology 139:4420–4423
Toshinai K, Date Y, Murakami N, Shimada M, Mondal MS, Shimbara T, Guan JL, Wang QP, Funahashi H, Sakurai T, Shioda S, Matsukura S, Kangawa K, Nakazato M (2003) Ghrelin-induced food intake is mediated via the orexin pathway. Endocrinology 144:1506–1512
Tschop M, Statnick MA, Suter TM, Heiman ML (2002) GH-releasing peptide-2 increases fat mass in mice lacking NPY: indication for a crucial mediating role of hypothalamic agouti-related protein. Endocrinology 143:558–568
Van den Pol AN, Trombley PQ (1993) Glutamate neurons in hypothalamus regulate excitatory transmission. J Neurosci 13:2829–2836
Van Tienhoven A, Juhaz LP (1962) The chicken telencephalon, diencephalons and mesencephalon in stereotaxic coordinates. J Comp Neural 118:185–197
Zendehdel M, Baghbanzadeh A, Babapour V, Cheraghi J (2009) The effects of bicuculline and muscimol on glutamate-induced feeding behavior in broiler cockerels. J Comp Physiol A 195:715–720
Zeni LA, Seidler HB, De Carvalho NA, Freitas CG, Marino-Neto J, Paschoalini MA (2000) Glutamatergic control of food intake in pigeons: effects of central injections of glutamate, NMDA, and AMPA receptor agonists and antagonists. Pharmacol Biochem Behav 65(1):67–74
Acknowledgment
This research was supported by a grant from Research Council of the College of Veterinary Medicine, Lorestan University and a grant from Research Council of the Faculty of Veterinary Medicine, University of Tehran. Animal handling and experimental procedures were performed according to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication NO.85-23, revised 1996) and also with the current laws of Iran.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Taati, M., Nayebzadeh, H. & Zendehdel, M. The effects of DL-AP5 and glutamate on ghrelin-induced feeding behavior in 3-h food-deprived broiler cockerels. J Physiol Biochem 67, 217–223 (2011). https://doi.org/10.1007/s13105-010-0066-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13105-010-0066-y