
Abstract
The growing popularity of novel psychoactive substances (NPS) has aroused the concerns of public health specialists. The pyrovalerone derivatives are a branch of synthetic cathinones, a very popular group of psychostimulant NPS. Despite numerous case reports of fatal intoxications, little is known about the cytotoxicity of these substances. Therefore, this study was aimed to evaluate the toxic properties of pyrovalerone, its highly prevalent derivative 3,4-methylenedioxypyrovalerone (3,4-MDPV) with its two major metabolites (catechol-MDPV and methylcatechol-MDPV) and the structural isomer 2,3-MDPV, together with newer members of the group, i.e., α-pyrrolidinovalerothiophenone (α-PVT) and α-pyrrolidinooctanophenone (PV9), using model human cell lines for neurons (SH-SY5Y), hepatocytes (Hep G2), and upper airway epithelium (RPMI 2650). We found that the first generation pyrovalerones (pyrovalerone, 3,4-MDPV, and 2,3-MDPV) produced a modest decrease of mitochondrial activity in the three examined cell lines, but were active in lower concentrations than methamphetamine used as a reference psychostimulant compound. Since catechol-MDPV displayed greater toxic potential than the parent compound, we suggest that the toxicity of 3,4-MDPV could be attributed to activity of this metabolite. Strikingly, the two new generation pyrovalerones, α-PVT and PV9, seem to be the most potent cytotoxic compounds: both induced highly pronounced mitochondrial dysfunction; the latter also demonstrated significant damage to cell membranes. The reported in vitro toxic activity of pyrovalerone cathinones against different cell types reinforces existing concerns regarding the health risks associated with the intake of these drugs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Synthetic cathinones and cannabimimetics are regarded as the most prevalent groups of novel psychoactive substances (NPS), and over the last decade have gained popularity as recreational drugs (Glennon 2014; Maciów-Głąb et al. 2014; Zawilska 2015). Synthetic cathinones usually appear on the market in a form of white or yellowish powder or pills (Zawilska and Wojcieszak 2013). Products are often mislabelled as bath salts, plant feeders, research chemicals, jewelry cleaner, ice melter, hookah pipe cleaner and designated not for human consumption (Marinetti and Antonides 2013; Zawilska and Andrzejczak 2015). Depending on the chemical structure and mechanism of action, cathinone derivatives may act as psychostimulants or empathogens/entactogens (Simmler et al. 2013). Pyrovalerone cathinones (α-pyrrolidinophenone derivatives) are an especially popular group within synthetic cathinones due to their strong psychostimulant activity. In fact, 3,4-methylenedioxypyrovalerone (3,4-MDPV), a prominent member of the α-pyrrolidinophenones, was the most widely abused synthetic cathinone in some EU countries and USA in 2013/2014 (EMCDDA 2014; Hasegawa et al. 2014; Rickli et al. 2015).
Pyrovalerone cathinones, including pyrovalerone, 3,4-MDPV, α-pyrrolidinovalerophenone (α-PVP), 3,4-methylenedioxy-α-pyrrolidinopropiophenone (3,4-MDPPP) and 3,4-methylenedioxy-α-pyrrolidinobutiophenone (3,4-MDPBP), exert their psychostimulant effects by selective and very potent inhibition of norepinephrine and dopamine reuptake by inhibition of NET and DAT transporters, respectively (Rickli et al. 2015). The potency of pyrovalerone and 3,4-MDPV to inhibit DAT is at least 10-times greater than that of cocaine (Simmler et al. 2013). None of the aforementioned compounds release monoamines or demonstrate any significant affinity towards serotonin receptors. Unlike other cathinones, such as mephedrone, α-pyrrolidinophenone derivatives have a high DAT:SERT inhibition ratio, resulting in a highly pronounced psychostimulant effect and very high abuse potential. On the other hand, they are devoid of empathogenic/entactogenic properties and have a less pronounced effect on thermoregulation, features that are characteristic of drugs increasing serotoninergic transmission (Glennon 2014; Gregg and Rawls 2014; Paillet-Loilier et al. 2014; Rickli et al. 2015; Simmler et al. 2013).
By analogy to other cathinones, pyrovalerones are most commonly administered intranasally and orally (Liechti 2015; Paillet-Loilier et al. 2014; Zawilska and Andrzejczak 2015). The dosage depends on the specific chemical as well as the user’s experience with drugs. As 3,4-MDPV is considered very potent, its dose is usually lower than those of other psychostimulants, and varies between 5 and 30 mg (http://erowid.org; Paillet-Loilier et al. 2014). Due to its popularity among recreational drugs users and high toxicity accompanied with a significant abuse potential, 3,4-MDPV has been scheduled in USA, Japan and most EU countries (EMCDDA 2014; Gregg and Rawls 2014; Kikura-Hanajiri et al. 2014; Rickli et al. 2015). As designer drugs typically enter the market faster than they can be restricted, new pyrovalerone analogs appeared following the ban of 3,4-MDPV. Among them, there are two new compounds containing pyrrolidine ring, namely α-PVT (α-pyrrolidinovalerothiophenone), which has been identified for the first time in Japan in 2013 in a product called “Fragrance Powder” (Uchiyama et al. 2013), and PV9 (α-POP, α-pyrrolidinooctanophenone) identified in Japan in 2014 in a product labeled “Brown Powder” (Hasegawa et al. 2014; Uchiyama et al. 2014). Doses of new generation pyrovalerones can reach as much as 120–250 mg for α-PVT (http://dopalator.info; http://hyperreal.info/talk/alfa-pvt) and 120 mg of PV9 (http://chemsrus.com; http://hyperreal.info/talk/alfa-pop), as these new drugs are commonly considered “light” or “polite” versions of 3,4-MDPV.
Pyrovalerone cathinones are abused in order to produce psychostimulant effects similar to those of amphetamine derivatives, including euphoria, rush, alertness, talkativeness, sexual arousal, a focused mind and overall positive feeling (Marinetti and Antonides 2013; Paillet-Loilier et al. 2014; Zawilska and Andrzejczak 2015). Due to the long duration of action as well as high potency, recreational use of pyrovalerones, especially 3,4-MDPV, is associated with significant risk of overdosing (Rickli et al. 2015). Moreover, the high lipophilicity of the pyrrolidine ring increases permeability through the blood–brain barrier. This feature, together with the active transport of pyrovalerones, enables higher concentrations to be achieved in the central nervous system than in the blood (Marinetti and Antonides 2013; Rickli et al. 2015; Simmler et al. 2013). There are numerous reports of pyrovalerone cathinones-related intoxications. Liver failure, sympathomimetic toxidrome with aggression, pronounced agitation, prolonged insomnia (which may last for several days), hypertension, chest pain, tachycardia, cardiac arrest, anxiety, fatigue, mydriasis, and combative behavior have been observed in many patients admitted to hospitals after consumption of α-pyrrolidinophenone derivatives. Psychiatric effects, including paranoid psychosis, panic, disorientation, memory loss, blackouts, paranoia, and suicidal ideations, are also common among abusers (Borek and Holstege 2012; Fröhlich et al. 2011; Gregg and Rawls 2014; Liechti 2015; Marinetti and Antonides 2013; Paillet-Loilier et al. 2014; Penders et al. 2012; Rickli et al. 2015; Stevenson and Tuddenham 2014; Zawilska and Andrzejczak 2015).
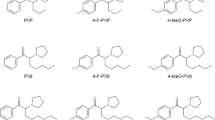
There are several reports of fatal intoxications with α-pyrrolidinophenone derivatives, including 3,4-MDPV, PV9, and α-PVP. These observations suggest that this group of synthetic cathinones is particularly dangerous to human life (EMCDDA 2014, 2015a, b; Hasegawa et al. 2014; Kudo et al. 2015; Nagai et al. 2014). In contrast to numerous case reports of intoxications with pyrovalerone cathinones, limited data exist on the in vitro cytotoxicity of these compounds. Therefore, the aim of this work was to investigate the effects of the following substances: pyrovalerone, 3,4-MDPV and its two main metabolites (catechol-MDPV and methylcatechol-MDPV), along with its structural isomer 2,3-MDPV, and two new generation pyrovalerones, namely α-PVT and PV9 (α-POP) (see Fig. 1), on viability of human cell lines. Based on the properties of the tested drugs and their patterns of use, the following model human cell lines were chosen:

Chemical structures of compounds investigated in this study
-
Neuroblastoma SH-SY5Y as a model to study neurotoxicity (Xie et al. 2010), which may result of the abuse of monoamine reuptake inhibitors;
-
Hep G2 derived from hepatoblastoma (Knasmüller et al. 2004) to examine hepatotoxicity: pyrovalerone derivatives are extensively metabolized in the liver and their concentrations in this organ are found to be the highest within the body (Marinetti and Antonides 2013);
-
RPMI 2650 established from human nasal septum carcinoma (Reichl and Becker 2012) to study cytotoxicity against the human upper airway epithelium: the main administration route of synthetic cathinones is snorting, which may lead to the exposure of epithelial cells to extremely high local concentrations of the drugs.
The cytotoxic effects of pyrovalerones were compared with those of methamphetamine used as a reference compound.
Materials and Methods
Drugs and Reagents
3,4-Methylenedioxypyrovalerone (3,4-MDPV; 1-(1,3-benzodioxol-5-yl)-2-(1-pyrrolidinyl)-1-pentanone), 2,3-methylenedioxypyrovalerone (2,3-MDPV; 1-(1,3-benzodioxol-4-yl)-2-(1-pyrrolidinyl)-1-pentanone), pyrovalerone (1-(4-methylphenyl)-2-(1-pyrrolidinyl)-1-pentanone); demethylenyl-methyl-MDPV (methylcatechol-MDPV; 1-(4-hydroxy-3-methoxyphenyl)-2-(pyrrolidin-1-yl)pentan-1-one), 3,4-dihydroxypyrovalerone (catechol-MDPV; 1-(3,4-dihydroxyphenyl)-2-(pyrrolidin-1-yl)pentan-1-one), α-pyrrolidinopentiothiophenone (α-PVT; 2-(pyrrolidin-1-yl)-1-(thiophen-2-yl)pentan-1-one), and PV9 (α-POP; 1-phenyl-2-(pyrrolidin-1-yl)octan-1-one) were purchased in the form of hydrochloride salts from Cayman Chemical (Ann Arbor, MI, USA). Cell culture media: DMEM, DMEM/F12 and MEM, heat inactivated fetal bovine serum (FBS), phosphate buffered saline (PBS), Trypsin–EDTA, Non-Essential Amino Acids Solution (NEAA), penicillin and streptomycin were purchased from Life Technologies (Warsaw, Poland). Dimethyl sulphoxide (DMSO), methamphetamine, MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide) and Triton X-100 were purchased from Sigma-Aldrich (Poznań, Poland).
Cell Lines
SH-SY5Y, Hep G2, and RPMI 2650 cell lines were purchased from Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany).
Cell Culture
Cells were cultured in DMEM/F12 (SH-SY5Y), DMEM (Hep G2) or MEM with Earle’s salts and 1 × Non-Essential Amino Acids Solution (RPMI 2650) medium supplemented with 10 % fetal bovine serum (FBS) and penicillin (100 U/ml)-streptomycin (100 μg/ml) at 37 °C in a humidified atmosphere enriched with 5 % CO2. Upon reaching 80–90 % confluency, cells were harvested with 0.25 % Trypsin in 1 mM EDTA and transferred into 96-well microplates at a density of 5000 cells/well (Hep G2 cell line) or 10,000 cells/well (SH-SY5Y and RPMI 2650 cell lines) for MTT assay. In the case of LDHe assay, the density was 10,000 cells/well for Hep G2 and RPMI 2650 cell lines, and 20,000 cells/well for SH-SY5Y cell line.
MTT Assay
Cell viability and mitochondrial function were evaluated by measuring the reduction of 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) by mitochondrial dehydrogenases. Following overnight incubation, complete culture medium was removed and replaced by fresh medium without FBS, and working solutions of tested compounds were added into microplate wells. After 24- and 72-h exposure times, a solution of MTT (0.5 mg/ml) was added and cells were incubated for an additional 3 h at 37 °C. After aspiration of culture medium, formazan crystals were dissolved in DMSO and absorbance was measured at 570 nm using a model 680 Bio-Rad microplate reader: the degree of absorbance being proportional to the number of cells with intact mitochondria. The results obtained with the MTT test were further confirmed using the LDHe assay. Each experiment included a positive control of 1 % (v/v) Triton X-100, added 30 min before MTT. The mean values of each group were obtained by subtracting the mean OD of the positive control. Because catechol-MDPV is able to reduce MTT nonenzymatically, all experiments were paralleled by blanks containing solutions of catechol-MDPV (10–300 μM) in culture medium and MTT without cells. The mean values for each group were calculated by subtraction of the corresponding blank.
LDHe Assay
Cellular membrane integrity was assessed by measuring the activity of lactate dehydrogenase released from damaged cells into the culture medium using an LDH Cytotoxicity Assay (ScienCell Research Laboratories, Carlsbad, CA, USA), according to the manufacturer’s instructions. Each experiment included a positive control of 1 % (v/v) Triton X-100, as recommended by the manufacturer.
Data Analysis
Results are expressed as mean ± SEM. Statistical analysis was performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA). The normality of distribution was checked using the Kolmogorov–Smirnov test with the Lilliefors test. Statistical evaluation was performed using analysis of variance (ANOVA). Groups were compared with controls using the Dunnett’s post hoc test in the case of normally distributed data and the Mann–Whitney test in the case of a non-normal distribution. Differences were considered significant when p < 0.05.
Results
Effects of Pyrovalerone on Survival of SH-SY5Y, Hep G2, and RPMI 2650 Cells
Pyrovalerone caused moderate, but statistically significant decreases in viability of SH-SY5Y and RPMI 2650 cells as measured by mitochondrial activity (Fig. 2a, c). The maximal effect was observed at 300 μM, i.e., a decrease in survival of SH-SY5Y cells by 25 and 46 %, and that of RPMI 2650 cells by 24 and 29 % of control values after 24 and 72 h of incubation, respectively. Incubation of Hep G2 cells with 300 μM of pyrovalerone for 72 h resulted in a statistically significant reduction of mitochondrial function by 27 % of control values. A shorter exposure time, 24 h, did not affect viability of Hep G2 cells (Fig. 2b). No cellular damage resulting in an increase of extracellular LDH activity was found in either of the cell lines (data not shown).

Effects of pyrovalerone on viability of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–18 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01, * p < 0.05 versus control group after 24 h incubation; ### p < 0.001, ## p < 0.01 versus control group after 72 h incubation
Effects of 2,3-MDPV on Survival of SH-SY5Y, Hep G2, and RPMI 2650 Cells
After 24-h incubation, 2,3-MDPV produced a concentration-dependent reduction of mitochondrial activity of Hep G2 and RPMI 2650 cells, reaching maximal effect at 300 μM (decreases by 25 and 33 % of control values, respectively; Fig. 3b, c), while no response was observed in SH-SY5Y cells (Fig. 3a). The 72-h exposure to 2,3-MDPV resulted in marked, concentration-dependent reductions of viability of all three cell lines at 300 μM: by 33 % in SH-SY5Y, 38 % in Hep G2, and 46 % in RPMI 2650 cells (Fig. 3). No cellular damage resulting in an increase of extracellular LDH activity was found in either of the cell lines (data not shown).

Effects of 2,3-MDPV on viability of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–18 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01 versus control group after 24-h incubation; ### p < 0.001, ## p < 0.01; # p < 0.05 versus control group after 72-h incubation
Effects of 3,4-MDPV and Its Two Metabolites, Methylcatechol-MDPV and Catechol-MDPV, on Survival of SH-SY5Y, Hep G2, and RPMI 2650 Cells
Incubation of SH-SY5Y cells with 100, 200, and 300 μM of 3,4-MDPV for 72 h resulted in a significant, concentration-dependent reduction in their viability by 21, 39, and 55 % of control values, respectively, whereas 24-h exposure to 300 μM evoked only a slight decrease by 16 % (Fig. 4a). The 3,4-MDPV-evoked decrease in RPMI 2650 cell viability was of similar potency after 24- and 72-h incubation, reaching maximal effect at 300 μM—a reduction by 30 and 26 % of control values, respectively (Fig. 4c). Despite changes in mitochondrial activity, no significant cellular membrane damage was detected using the LDHe assay (data not shown). As observed with pyrovalerone, 3,4-MDPV (300 μM) was cytotoxic to Hep G2 cells only after 72-h incubation (Fig. 4b).

Effects of 3,4-MDPV on viability of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–18 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01 versus control group after 24-h incubation; ### p < 0.001, ## p < 0.01, # p < 0.05 versus control group after 72-h incubation
Methylcatechol-MDPV did not induce any changes of mitochondrial activity nor cellular membrane integrity in any examined cell line, neither after 24- nor after 72-h incubation (data not shown).
Catechol-MDPV produced changes in cell viability that were time-, concentration-, and cell line-dependent. SH-SY5Y cells responded to catechol-MDPV after 24-h incubation at 200 and 300 μM, which resulted in decreases of viability by 22 and 45 % of control values, respectively. The cytotoxic effect was more pronounced after 72-h incubation, resulting in reduced cell viability at concentrations as low as 25 μM, with a maximal reduction by 87 % of control values at 300 μM (Fig. 5, top panel a). Along with disturbances in mitochondrial activity, cell membrane disruptions were observed at 200 and 300 μM, reaching 50 and 53 % of a positive control toxicity, respectively (Fig. 5, bottom panel a). The exposure of Hep G2 cells to catechol-MDPV for 24 h resulted in a significant, concentration-dependent decrease of mitochondrial activity, maximally by 48 % of control values at 300 μM (Fig. 5, top panel b). Extending incubation time to 72 h led to even more potent concentration-dependent decrease in cell survival (maximally by 79 % of control values) accompanied by cell membrane damage equal to 58 % of the positive control at 300 μM (Fig. 5, top panel b and bottom panel b). In RPMI 2650 cells, catechol-MDPV exposure was associated with time- and concentration-dependent decreases of mitochondrial activity by up to 66 % (24 h) and 91 % (72 h) of control values (Fig. 5, top panel c). In addition, a significant disruption of cellular membranes was observed in this cell line reaching 64 % of the positive control at 300 μM (Fig. 5, bottom panel c). It is noteworthy that in SH-SY5Y and RPMI 2650 cells, a significant damage to cellular membranes was observed also after 24-h incubation with 300 μM of catechol-MDPV. These values were equal to 33 % (SH-SY5Y) and 15 % (RPMI) of positive controls (data not shown).

Top Effects of catechol-MDPV on viability of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–18 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01 versus control group after 24-h incubation; ### p < 0.001, ## p < 0.01 versus control group after 72-h incubation. Bottom Effects of catechol-MDPV on the cell membrane integrity of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by LDHe test. Results are given as mean ± SEM of 10–20 values per group and expressed as a percentage of the positive control (1 % v/v Triton-X100). *** p < 0.001, ** p < 0.01 versus negative control group (untreated cells)
Effects of PV9 on Survival of SH-SY5Y, Hep G2, and RPMI 2650 Cells
Exposure to PV9 for both 24 and 72 h resulted in a highly pronounced and concentration-dependent decrease in survival of the three examined cell lines. After 72-h incubation with 300 μM PV9, almost no mitochondrial activity was detected (Fig. 6, top panel). Furthermore, PV9 produced potent increases in cellular membrane permeability. In SH-SY5Y and Hep G2 cells, a significant cell membrane damage was observed at 200 and 300 μM, reaching maximal effect equal to 92 and 77 % of positive controls, respectively. In RPMI 2650 cells, significant disruption of cellular membrane was observed at 100 μM, with damage equal to 71 % of the positive control at 300 μM (Fig. 6, bottom panel).

Top Effects of PV9 on viability of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–18 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01, * p < 0.05 versus control group after 24-h incubation; ### p < 0.001, ## p < 0.01 versus control group after 72-h incubation. Bottom Effects of PV9 on the cell membrane integrity of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by LDHe test. Results are given as mean ± SEM of 10-30 values per group and expressed as a percentage of positive control (1 % v/v Triton-X100). *** p < 0.001, ** p < 0.01 versus negative control group (untreated cells)
Effects of α-PVT on Survival of SH-SY5Y, Hep G2, and RPMI 2650 Cells
α-PVT produced a concentration-dependent decrease of mitochondrial activity of SH-SY5Y cells starting at 50 and 25 μM after 24- and 72-h incubation, respectively, with maximum reductions by 22 % (24 h) and 44 % (72 h) of control values (Fig. 7a). Interestingly, in Hep G2 cells, α-PVT acted at concentrations as low as 2.5 μM (24 h) and 1 μM (72 h), reaching a plateau at 100 μM after 24 h (reduction in cell viability by appr. 65 % of control values) and at 50 μM after 72 h (reduction in cell viability by appr. 55 % of control values) (Fig. 7b). After 24-h incubation with RPMI 2650 cells, α-PVT produced a concentration-dependent decrease of their viability, starting at 25 μM and peaking at 300 μM (reduction by 50 % of control values). On the other hand, 72-h incubation produced a slightly less pronounced effect (onset at 100 μM, maximal decrease by 41 % of control values at 300 μM) (Fig. 7c). Interestingly, even the explicit decrease of mitochondrial activity observed in Hep G2 cells was not accompanied by damage of the cell membranes (data not shown).

Effects of α-PVT on viability of SH-SY5Y, Hep G2 and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–24 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01, * p < 0.05 versus control group after 24-h incubation; ### p < 0.001, ## p < 0.01 versus control group after 72-h incubation
Methamphetamine
Methamphetamine (100–2000 μM), used as a reference compound, induced a time- and concentration-dependent decrease of cell viability. However, the cytotoxic effect of methamphetamine was observed at concentrations markedly higher than these of examined pyrovalerones. In the SH-SY5Y cell line exposed to methamphetamine for 24 h, toxic effects were observed starting from 750 μM and a maximum decrease by 43 % of control values was found at 2000 μM. The toxicity of methamphetamine was higher after 72 h, starting at 300 μM and leading to the death of 77 % cells at 2000 μM (Fig. 8a). In Hep G2 cells exposed to methamphetamine for 24 h, the drug started to exert cytotoxic activity at 750 μM. The maximal reduction in cell viability, by 62 % of control values, was observed at 2000 μM. Incubation of Hep G2 cells with 2000 μM methamphetamine for 72 h resulted in an almost total reduction of viable cells (Fig. 8b). The cytotoxic effect of methamphetamine against RPMI 2650 cells was first observed at 300 and 750 μM after 24 and 72 h, respectively. The exposition to 2000 μM methamphetamine led to a maximal decrease of cell viability by 40 and 82 % of control values after 24 and 72 h, respectively (Fig. 8c). Besides of the distinct changes in mitochondrial activity, methamphetamine at 2000 μM after 24-h incubation did not cause a significant damage to the SH-SY5Y and RPMI 2650 cell membranes, while in Hep G2 cells, the increase of extracellular LDH activity was observed only at 1500 and 2000 μM (by 44 and 142 % of negative control values, which correspond to 9 and 15 % of positive controls, respectively) (data not shown).

Effects of methamphetamine on viability of SH-SY5Y, Hep G2, and RPMI 2650 cells, measured by MTT test. Results are given as mean ± SEM of 6–18 values per group and expressed as a percentage of the respective control (untreated cells). *** p < 0.001, ** p < 0.01, * p < 0.05 versus control group after 24-h incubation; ### p < 0.001, ## p < 0.01 versus control group after 72-h incubation
Discussion
The growing popularity of novel psychoactive substances is a matter of concern for public health specialists. Despite their short presence on the clandestine market, pyrovalerone derivatives, including the very recently introduced PV9, have caused several fatal intoxications (EMCDDA 2014, 2015b; Kudo et al. 2015; Marinetti and Antonides 2013). Therefore, to gain more information on the harmful properties of this emerging group of drugs of abuse, it is crucial to investigate its in vitro toxicity. The present study reports the effects of pyrovalerone derivatives on viability of three human cell lines used as models of neurons, hepatocytes, and the airway epithelium. Pyrovalerone, 3,4-MDPV, and 2,3-MDPV tested at concentrations up to 300 μM caused weak to moderate reductions of mitochondrial activity, but did not produce any significant damage to cell membranes in SH-SY5Y, Hep G2, and RPMI 2650 cells. It is noteworthy that α-PVT and PV9, the new members of the α-pyrrolidinophenone family, were more cytotoxic than the older compounds, i.e., pyrovalerone and 3,4-MDPV, which on the other hand were active in concentrations markedly lower than those of the reference compound, methamphetamine. Out of the two, PV9 desires greater attention, as it evoked not only a potent decrease in mitochondrial activity, but also produced a pronounced damage to cellular membranes. This observation could raise a concern that each new wave of psychostimulant pyrovalerones might be burdened with greater ability to cause cellular damage than the previous ones. Additionally, the cytotoxic potential of designer cathinones seems to be dependent on the presence of 1-pyrrolidinyl ring in their chemical structure, since none of the earlier examined by us nonpyrovalerone cathinone derivatives (i.e., methcathinone, 3-fluoromethcathinone, and 2,3-methylenedioxymethcathinone) applied in concentrations up to 300 μM produced any significant changes in viability of SH-SY5Y, Hep G2, and RPMI 2650 cells (Wojcieszak J., unpublished data).
The concentrations of drugs used in this in vitro study exceeded those found in blood and urine samples collected from abusers (Elliott 2005; Kudo et al. 2015; Marinetti and Antonides 2013). However, it should be emphasized that blood concentrations of α-pyrrolidinophenone derivatives do not directly reflect their organ/tissue levels. Due to their high lipophilicity, these substances have a high volume of distribution, meaning that they reach significantly greater concentrations in lipophilic tissues such as the central nervous system (Rickli et al. 2015). For instance, in postmortem studies, 3,4-MDPV concentrations were found to be up to four times higher in the brain than in peripheral blood, while the highest levels were detected in the liver and bile duct: these concentrations being as much as 23-times higher than in peripheral blood (Marinetti and Antonides 2013). Liver concentrations were also found larger by an order of magnitude for other psychostimulant drugs of abuse, such as methylenedioxymethamphetamine (MDMA) (Araújo et al. 2015). As the liver is the main site of metabolism and particularly high levels of pyrovalerones can be found in this organ, hepatocytes may be considered vulnerable targets for the cytotoxic action of these drugs.
It is well known that snorting is a popular route of administration of psychostimulants (Glennon 2014; Liechti 2015; Paillet-Loilier et al. 2014). Therefore, it is reasonable to assume that exposure to high levels of α-pyrrolidinophenone derivatives may also occur locally in the upper airway epithelial cells. It should be emphasized that a certain lag time following drug intake typically exists before blood samples are collected, which may result in lower concentrations being identified, due to metabolic processes and antemortem/postmortem redistribution of the lipid-soluble compound. Therefore, the blood concentration of a particular drug of abuse does not give much information about the antemortem exposure of crucial organs (Elliott 2005). Finally, in this study, we used cell lines derived from tumors as a model of certain cell types; such cell lines could be less sensitive to harmful actions of various chemical compounds. Thus for example, it has been previously demonstrated that methamphetamine-induced cytotoxicity is much lower in SH-SY5Y neuroblastoma cells than in primary neuronal cultures (den Hollander et al. 2014).
Although little is currently known about the direct mechanism of pyrovalerone toxicity, mitochondrial dysfunction and oxidative stress are considered to be major contributors to the hepatotoxicity induced by amphetamine derivatives (Nakagawa et al. 2009). Similarly, 3,4-MDPV (48-h incubation at 37 and 40.5 °C) was found to produce impairment of mitochondrial activity, disruption of cell membrane, depletion of intracellular glutathione, accumulation of reactive oxygen (ROS), and nitrogen (RNS) species, as well as depletion of intracellular ATP and activation of caspases in primary rat hepatocytes (Valente et al. 2015). At 37 °C cytotoxic effects of 3,4-MDPV were observed at significantly higher concentrations than these found at 40.5 °C, and exposure to 400 μM 3,4-MDPV at 37 °C led only to moderate disturbances of mitochondrial function, cell membrane integrity and total GSH level (Valente et al. 2015).
It has also been suggested that the toxicity of MDMA and related compounds may be attributed to their metabolites and the reactive species generated during enzymatic breakdown (Araújo et al. 2015; den Hollander et al. 2015; Nakagawa et al. 2009). It is postulated that catechols produced in the phase I metabolism can undergo oxidation to ortho-quinones with their semiquinones, which can further react with other molecules, interfere with redox systems and cause oxidative damage within the cells (Araújo et al. 2015; Nakagawa et al. 2009). In line with this, our study revealed that one of the major 3,4-MDPV metabolites, catechol-MDPV (Meyer et al. 2010; Strano-Rossi et al. 2010), was significantly more potent than the parent compound in all examined cell lines. This, in turn, suggests that the catechol-metabolite of 3,4-MDPV is a major contributor to the drug’s toxicity, especially in liver cells, where the parent compound did not produce significant toxicity after 24 h, while the catechol-metabolite caused significant decrease of viability at 50 μM.
Conclusions
Since numerous new cathinone derivatives appear on the market each year, there is an urgent need for a broad screening procedure for their toxic properties. This study provides a comparison of the in vitro cytotoxicity of five α-pyrrolidinophenone derivatives, along with two major metabolites of 3,4-MDPV, against cell lines used as models of neurons, hepatocytes and upper airway epithelial cells. Among the tested compounds, PV9 and α-PVT, the newest members of the pyrovalerone family, exhibited the greatest cytotoxic activity. The reported in vitro toxic activity of pyrovalerone cathinones against different cell types supports the existing concerns regarding the health risks associated with the intake of these drugs. Studies on the molecular mechanisms involved in cytotoxicity induced by α-pyrrolidinophenone derivatives are warranted in order to develop risk management procedures to decrease their morbidity and mortality.
References
Araújo AM, Valente MJ, Carvalho M, Dias da Silva D, Gaspar H, Carvalho F, de Lourdes Bastos M, Guedes de Pinho P (2015) Raising awareness of new psychoactive substances: chemical analysis and in vitro toxicity screening of ‘legal high’ packages containing synthetic cathinones. Arch Toxicol 89(5):757–771. doi:10.1007/s00204-014-1278-7
Borek HA, Holstege CP (2012) Hyperthermia and multiorgan failure after abuse of “bath salts” containing 3,4-methylenedioxypyrovalerone. Ann Emerg Med 60(1):103–105. doi:10.1016/j.annemergmed.2012.01.005
den Hollander B, Sundström M, Pelander A, Ojanperä I, Mervaala E, Korpi ER, Kankuri E (2014) Keto amphetamine toxicity-focus on the redox reactivity of the cathinone designer drug mephedrone. Toxicol Sci 141(1):120–131. doi:10.1093/toxsci/kfu108
den Hollander B, Sundström M, Pelander A, Siltanen A, Ojanperä I, Mervaala E, Korpi ER, Kankuri E (2015) Mitochondrial respiratory dysfunction due to the conversion of substituted cathinones to methylbenzamides in SH-SY5Y cells. Sci Rep 5:14924. doi:10.1038/srep14924
Elliott SP (2005) MDMA and MDA concentrations in antemortem and postmortem specimens in fatalities following hospital admission. J Anal 29(5):296–300
EMCDDA (2014) Report on the risk assessment of 1-(1,3-benzodioxol-5-yl)-2-(pyrrolidin-1-yl) pentan-1-one (3,4-methylenedioxypyrovalerone, MDPV) in the framework of the Council Decision on new psychoactive substances. http://www.emcdda.europa.eu/attachements.cfm/att_228256_EN_TDAK14003ENN.pdf. Retrieved 22 Oct 2014
EMCDDA (2015a) New psychoactive substances in Europe. An update from the EU Early Warning System (March 2015). http://www.emcdda.europa.eu/attachements.cfm/att_235958_EN_TD0415135ENN.pdf. Accessed 23 April 2015
EMCDDA (2015b) α-PVP EMCDDA-Europol joint report on a new psychoactive substance: 1-phenyl-2-(1-pyrrolidinyl)-1-pentanone (α-PVP). http://www.emcdda.europa.eu/attachements.cfm/att_242501_EN_TDAS15001ENN.pdf. Accessed 30 Sept 2015
Fröhlich S, Lambe E, O’Dea J (2011) Acute liver failure following recreational use of psychotropic “head shop” compounds. Ir J Med Sci 180(1):263–264. doi:10.1007/s11845-010-0636-6
Glennon RA (2014) Bath salts, mephedrone, and methylenedioxypyrovalerone as emerging illicit drugs that will need targeted therapeutic intervention. Adv Pharmacol 69:581–620. doi:10.1016/B978-0-12-420118-7.00015-9
Gregg RA, Rawls SM (2014) Behavioral pharmacology of designer cathinones: a review of the preclinical literature. Life Sci 97(1):27–30. doi:10.1016/j.lfs.2013.10.033
Hasegawa K, Wurita A, Minakata K, Gonmori K, Nozawa H, Yamagishi I, Suzuki O, Watanabe K (2014) Identification and quantitation of a new cathinone designer drug PV9 in an “aroma liquid” product, antemortem whole blood and urine specimens, and a postmortem whole blood specimen in a fatal poisoning case. Forensic Toxicol 32(2):243–250. doi:10.1007/s11419-014-0230-0
http://www.chemsrus.com/forum/6-stimulants/55159-pv9-a-pop-roas?start=10
Kikura-Hanajiri R, Kawamura NU, Goda Y (2014) Changes in the prevalence of new psychoactive substances before and after the introduction of the generic scheduling of synthetic cannabinoids in Japan. Drug Test Anal 6(7–8):832–839. doi:10.1002/dta.1584
Knasmüller S, Mersch-Sundermann V, Kevekordes S, Darroudi F, Huber WW, Hoelzl C, Bichler J, Majer BJ (2004) Use of human-derived liver cell lines for the detection of environmental and dietary genotoxicants; current state of knowledge. Toxicology 198(1–3):315–328. doi:10.1016/j.tox.2004.02.008
Kudo K, Usumoto Y, Kikura-Hanajiri R, Sameshima N, Tsuji A, Ikeda N (2015) A fatal case of poisoning related to new cathinone designer drugs, 4-methoxy PV8, PV9, and 4-methoxy PV9, and a dissociative agent, diphenidine. Legal Med (Tokyo) 17(5):421–426. doi:10.1016/j.legalmed.2015.06.005
Liechti M (2015) Novel psychoactive substances (designer drugs): overview and pharmacology of modulators of monoamine signaling. Swiss Med Wkly 145:w14043. doi:10.4414/smw.2015.14043
Maciów-Głąb M, Rojek S, Kula K, Kłys M (2014) “New designer drugs” in aspects of forensic toxicology. Arch Med Sadowej Kryminol 64(1):20–33. doi:10.5114/amsik.2014.44587
Marinetti LJ, Antonides HM (2013) Analysis of synthetic cathinones commonly found in bath salts in human performance and postmortem toxicology: method development, drug distribution and interpretation of results. J Anal Toxicol 37(3):135–146. doi:10.1093/jat/bks136
Meyer MR, Du P, Schuster F, Maurer HH (2010) Studies on the metabolism of the α-pyrrolidinophenone designer drug methylenedioxypyrovalerone (MDPV) in rat and human urine and human liver microsomes using GC-MS and LC-high-resolution MS and its detectability in urine by GC-MS. J Mass Spectrom 45(12):1426–1442. doi:10.1002/jms.1859
Nagai H, Saka K, Nakajima M, Maeda H, Kuroda R, Igarashi A, Tsujimura-Ito T, Nara A, Komori M, Yoshida K (2014) Sudden death after sustained restraint following self-administration of the designer drug α-pyrrolidinovalerophenone. Int J Cardiol 172:263–265. doi:10.1016/j.ijcard.2013.12.262
Nakagawa Y, Suzuki T, Tayama S, Ishii H, Ogata A (2009) Cytotoxic effects of 3,4-methylenedioxy-N-alkylamphetamines, MDMA and its analogues, on isolated rat hepatocytes. Arch Toxicol 83(1):69–80. doi:10.1007/s00204-008-0323-9
Paillet-Loilier M, Cesbron A, Le Boisselier R, Bourgine J, Debruyne D (2014) Emerging drugs of abuse: current perspectives on substituted cathinones. Subst Abuse Rehabil 5:37–52. doi:10.2147/SAR.S37257
Penders TM, Gestring RE, Vilensky DA (2012). Intoxication delirium following use of synthetic cathinone derivatives. Am J Drug Alcohol Abuse 38(6):616–617. doi:10.3109/00952990
Reichl S, Becker K (2012) Cultivation of RPMI 2650 cells as an in vitro model for human transmucosal nasal drug absorption studies: optimization of selected culture conditions. J Pharm Pharmacol 64(11):1621–1630. doi:10.1111/j.2042-7158.2012.01540.x
Rickli A, Hoener MC, Liechti ME (2015) Monoamine transporter and receptor interaction profiles of novel psychoactive substances: para-halogenated amphetamines and pyrovalerone cathinones. Eur Neuropsychopharmacol 25(3):365–376. doi:10.1016/j.euroneuro.2014.12.012
Simmler LD, Buser TA, Donzelli M, Schramm Y, Dieu LH, Huwyler J, Chaboz S, Hoener MC, Liechti ME (2013) Pharmacological characterization of designer cathinones in vitro. Br J Pharmacol 168(2):458–470. doi:10.1111/j.1476-5381.2012.02145.x
Stevenson R, Tuddenham L (2014) Novel psychoactive substance intoxication resulting in attempted murder. J Forensic Leg Med 25:60–61. doi:10.1016/j.jflm.2014.04.007
Strano-Rossi S, Cadwallader AB, de la Torre X, Botrè F (2010) Toxicological determination and in vitro metabolism of the designer drug methylenedioxypyrovalerone (MDPV) by gas chromatography/mass spectrometry and liquid chromatography/quadrupole time-of-flight mass spectrometry. Rapid Commun Mass Spectrom 24(18):2706–2714. doi:10.1002/rcm.4692
Uchiyama N, Matsuda S, Kawamura M, Kikura-Hanajiri R, Goda Y (2013) Two new-type cannabimimetic quinolinyl carboxylates, QUPIC and QUCHIC, two new cannabimimetic carboxamide derivatives, ADB-FUBINACA and ADBICA, and five synthetic cannabinoids detected with a thiophene derivative α-PVT and an opioid receptor agonist AH-7921 identified in illegal products. Forensic Toxicol 31(2):223–240. doi:10.1007/s11419-013-0182-9
Uchiyama N, Matsuda S, Kawamura M, Shimokawa Y, Kikura-Hanajiri R, Aritake K, Urade Y, Goda Y (2014) Characterization of four new designer drugs, 5-chloro-NNEI, NNEI indazole analog, α-PHPP and α-POP, with 11 newly distributed designer drugs in illegal products. Forensic Sci Int 243:1–13. doi:10.1016/j.forsciint.2014.03.013
Valente MJ, Araújo AM, Silva R, Bastos ML, Carvalho F, Guedes de Pinho P, Carvalho M (2015) 3,4-Methylenedioxypyrovalerone (MDPV): in vitro mechanisms of hepatotoxicity under normothermic and hyperthermic conditions. Arch Toxicol. doi:10.1007/s00204-015-1653-z
Xie H, Hu L, Li G (2010) SH-SY5Y human neuroblastoma cell line: in vitro cell model of dopaminergic neurons in Parkinson’s disease. Chin Med J 123:1086–1092
Zawilska JB (2015) “Legal highs”—an emerging epidemic of novel psychoactive substances. Int Rev Neurobiol 120:273–300. doi:10.1016/bs.irn.2015.02.009
Zawilska JB, Andrzejczak D (2015) Next generation of the novel psychoactive substances on the horizon—a complex problem to face. Drug Alcohol Depend 157:1–17. doi:10.1016/j.drugalcdep.2015.09.030
Zawilska JB, Wojcieszak J (2013) Designer cathinones—an emerging class of novel recreational drugs. Forensic Sci Int 231(1–3):42–53. doi:10.1016/j.forsciint.2013.04.015
Acknowledgments
Supported by the National Science Centre (NCN), Cracow, Poland (Grant No. 2014/13/B/NZ7/02237).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wojcieszak, J., Andrzejczak, D., Woldan-Tambor, A. et al. Cytotoxic Activity of Pyrovalerone Derivatives, an Emerging Group of Psychostimulant Designer Cathinones. Neurotox Res 30, 239–250 (2016). https://doi.org/10.1007/s12640-016-9640-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12640-016-9640-6