
Abstract
The probiotic properties and inhibitory effect on Salmonella Typhimurium adhesion on human enterocyte-like HT-29-Luc cells of three Lactobacillus plantarum strains isolated from fermented fish, beach sand and a coastal plant were determined. Compared with the type strain L. plantarum NBRC 15891T, which was isolated from pickled cabbage, L. plantarum Tennozu-SU2 isolated from the acorn of a coastal tree showed high autoaggregation in de Man, Rogosa and Sharpe (MRS) broth and an antagonistic effect against S. Typhimurium in brain heart infusion (BHI) broth. Furthermore, heat-killed L. plantarum Tennozu-SU2 cells inhibited S. Typhimurium adhesion on HT-29-Luc cells. Both live and heat-killed L. plantarum Tennozu-SU2 cells showed an inhibitory effect on gut colonisation in BALB/c mice, as assessed by viable Salmonella count in faecal samples and by invasion into liver and spleen tissues. The properties shown in this study suggest that L. plantarum Tennozu-SU2 is useful as a starter and probiotic bacteria in functional food material.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Salmonella is a well-documented pathogen known to contaminate a wide range of foods, including chicken, egg products [1, 2] and other meats [3], vegetables [4] and dried fish products [5]. Although most cases of human infection with Salmonella enterica subsp. enterica serotype Enteritidis result from the consumption of contaminated foods, the incidence of salmonellosis from Salmonella Typhimurium has remained relatively stable or increased in Europe and other countries [6–8]. Although it is a gram-negative bacterium, S. Typhimurium shows resistance not only to disinfectants but also to dry and acidified environments [9, 10]. Acid tolerance response (ATR) of pathogenic bacteria, such as that to an amino acid decarboxylase, is noted as an adaptive strategy in foods, as well as in humans and other hosts [11]. Additionally, some strains of S. Typhimurium, such as DT104, show multidrug resistance [12].
Various lactic acid bacteria (LAB) strains are used as probiotics. Immune promotive effects, and antagonistic capacities against food-related pathogens are important properties of probiotics. Lactobacillus plantarum has been isolated from plant materials as well as various environments, such as fermented foods (high salinity and acidified), animal intestine and beach cast algae and used as a starter and probiotic [13–15]. There are many reports regarding the immune-promoting activities of LAB, and L. plantarum strains in particular, both in vitro and in vivo [16–20]. Inhibitory effects of several species of LAB, particularly Lactobacillus acidophilus and Lactobacillus casei, on S. Typhimurium infection in mice have also been reported by many researchers [21–24]. However, the inhibitory effect of L. plantarum in mice is not clear, although there are a few studies using human enterocyte-like Caco-2 cells in vitro [25].
Some LAB, including L. plantarum strains isolated from areas near the coast, showed stress resistance against salt, acid and bile, as well as bile acid-lowering capacity [26–29]. These strains can be considered for use as probiotics. In this study, the effect of three L. plantarum strains, isolated from fermented fish, beach sand and a coastal plant, on S. Typhimurium adhesion on human enterocyte-like HT-29-Luc cells was assessed. In addition, the inhibitory effect of a selected strain, administered via drinking water, on S. Typhimurium infection in BALB/c mice was investigated.
Materials and Methods
Bacterial Strains
A bacterial strain S. enterica subsp. enterica serovar Typhimurium NBRC 13245T was pre-incubated in Luria-Bertani (LB) broth (Becton, Dickinson and Co., Sparks, MD) at 37 °C for 20 h (early stationary growth phase). Four L. plantarum strains were used in this study. L. plantarum Sanriku-SU7 (accession no. LC122588) was isolated from fermented fish made in Iwate, Japan [13]. L. plantarum Izu-SU2 (accession no. LC144968) and Tennozu-SU2 (accession no. LC144970) were isolated from beach sand in the Izu Peninsula and from acorns in a coastal park in Tokyo, Japan, respectively [30]. L. plantarum NBRC 15891T isolated from pickled cabbage was used for the typical strain. These L. plantarum strains were pre-incubated in de Man, Rogosa and Sharpe (MRS) broth (Oxoid, Basingstoke, UK) at 37 °C for 24 h.
Effects of Acid and Bile on the Growth of L. plantarum Strains
The effects of acid- and bile-induced stress were determined as previously reported [31], with slight modification. The pre-cultures of four L. plantarum strains (0.03 mL) were inoculated into 3 mL of MRS broth (control broth), MRS broth with pH 4.1 or MRS broth containing 3% (w/v) of oxgall (Wako Pure Chemical, Osaka, Japan) and incubated at 37 °C for 24 h. The increased cell and other insoluble product amounts during the incubation were expressed as the optical density (OD) at 660 nm (OD660) that was measured using a grating microplate reader (SH-1000Lab; Corona, Electric, Hitachinaka, Ibaraki, Japan). OD660 values in the acidified or bile-containing broths were compared with ones from the control broth.
Autoaggregation Assay
Autoaggregation assays were performed according to the method described by Kos et al. [32] with some modification. The strains were cultured in MRS broth for 24 h at 37 °C. The cells were harvested by centrifugation at 5000g for 10 min at 4 °C, washed twice and resuspended in phosphate-buffered saline (PBS, Nissui Pharmaceutical, Tokyo, Japan) to adjust OD600 to 1.0. From each cell suspension, a 4-mL aliquot was mixed by vortexing for 10 s and incubated for 2 h at 37 °C. The autoaggregation (%) was expressed as (1 − (OD600 after 2-h incubation / OD600 before incubation) × 100.
HT-29-Luc Cell Adhesion Assay
The human enterocyte-like HT-29-Luc cells JCRB 1383 were purchased from Japanese Collection of Research Bioresources Cell Bank, Osaka, Japan. The adhesion assay was performed according to a previously reported method, with some modification [33]. The cells were seeded in 48-well plates at a density of 6 log cells/mL in Dulbecco’s modified Eagle’s medium (DMEM, Wako Pure Chemical) containing 10% v/v foetal bovine serum and were incubated at 37 °C in an atmosphere of 5% CO2–95% air for 8 days to facilitate cell differentiation. During this period, the medium was replaced with fresh medium every other day. Alkaline phosphatase activity in the cells was determined by colorimetric assay to confirm the differentiation [28], as previously reported [34].
The L. plantarum grown in MRS broth was centrifuged and resuspended in DMEM (9 log CFU/mL). After HT-29-Luc cell differentiation, 0.05 mL of this L. plantarum cell suspension was added (n = 3) and the mixture was incubated for 1.5 h in a CULTUREPAL™ system (Mitsubishi Gas Chemical, Tokyo, Japan). Subsequently, the wells were washed with PBS thrice to remove non-adherent cells. Gram staining was performed on the attached cells after methanol fixation [35]. The blue-stained cells in three fields of view for each well under a microscope were counted.
Antagonistic Effect Against S. Typhimurium in BHI Broth
The pre-culture of S. Typhimurium was diluted with 1000 times volume of PBS. The undiluted pre-culture (0.03 mL) of L. plantarum strain with the diluted S. Typhimurium culture (0.03 mL) was inoculated into 3 mL of brain heart infusion (BHI) broth (Oxoid). After incubation at 37 °C for 24 h, viable count of Salmonella was enumerated using deoxycholate hydrogen-sulphide lactose (DHL) agar (Eiken Chemical, Tokyo, Japan).
Effect of Heat-Killed L. plantarum Cells on S. Typhimurium Adhesion in HT-29-Luc Cells
The inhibitory effect was determined as reported previously [36]. The L. plantarum cells cultured in MRS broth were centrifuged, resuspended in DMEM (9 log CFU/mL) and then heated in boiling water for 20 min. The heat-killed cell suspension (0.05 mL) was added to the differentiated HT-29-Luc cell culture in a 48-well plate (0.5 mL/well). After incubation for 18 h, 0.05-mL medium containing S. Typhimurium cells (9 CFU/mL) was added (n = 4) and the mixture was incubated for 1.5 h using the CULTUREPAL system. The attached cells were removed as described previously. The attached cells were treated with cold PBS containing 0.1% Triton X-100 (Wako Pure Chemical) for 10 min. Then, the number of viable Salmonella in the detached suspension was determined using DHL agar.
Effect of Live and Heat-Killed L. plantarum Cells on S. Typhimurium Infection in BALB/c Mice
A selected L. plantarum strain was cultured in MRS broth, washed with PBS and suspended in distilled water (DW) at an OD600 of 2.0. Half of the suspension was heated in boiling water. The animal experiments were performed in compliance with the Fundamental Guidelines for Proper Conduct of Animal Experiment and Related Activities in Academic Research Institutions, under the jurisdiction of the Ministry of Education, Culture, Sports, Science and Technology, and approved by the animal experiment committee of Tokyo University of Marine Science and Technology (approval no. H28-4).
Mice were treated with the L. plantarum cell suspension and inoculated with S. Typhimurium using a previously described method [13, 33]. In total, 18 five-week-old male BALB/c mice were obtained from CLEA Japan (Tokyo, Japan). The mice were acclimatised in a negative pressure rack maintained at 20–25 °C with 50–60% relative humidity and were fed CE-2 diet (CLEA Japan) for 5 days prior to the experiment. They were then divided into three groups of six mice each and fed untreated water (control group), live cell suspensions or heat-killed cell suspension as drinking water for the next week. The mice were then anaesthetised using diethyl ether, and as soon as they were mildly sedated, they were administered 0.1 mL S. Typhimurium suspension (8 log CFU/mL in PBS) via a flexible gastric feeding tube (Model 5200, Fuchigami, Kyoto, Japan). The mice inoculated with S. Typhimurium were fed the same diet for the next 7 days.
Before the inoculation and 2 and 4 days after the inoculation, viable Salmonella counts were determined with DHL agar. The typical black colonies were counted as Salmonella. Seven days after the inoculation, the mice were euthanised by CO2, and then, the liver and spleen were aseptically removed and weighed. DHL agar was used to detect Salmonella that had invaded the organs. Data were expressed as mean ± standard error of the mean log CFU of S. Typhimurium per gram of tissue.
Statistical Analysis
Statistical analysis was performed using the software EXCEL statistical Ver. 6.0 (Esumi, Tokyo, Japan). One-way ANOVA was used to assess the differences between the means. Individual mean values were compared by Tukey’s test, with the level of statistical significance set at p < 0.05.
Results and Discussion
Effects of Acid and Bile on the Growth of L. plantarum Strains
The strains grew in the control broth from approximately 6 log CFU/ml (OD660 nm ≒ 0) to 8 log CFU/ml (OD660 nm ≒ 1). As shown in Fig. 1A, all four tested L. plantarum strains grew at pH 4.1, though the OD was lower compared with that at pH 6.5 (control broth). There was no significant difference between the acid tolerance of the type strain and the other strains. In the case of MRS broth containing 3% bile, L. plantarum Izu-SU2 showed the highest tolerance with a growth higher than that in the control broth (Fig. 1B). The tolerance of L. plantarum Sanriku-SU7 and Tennozu-SU2 was slightly higher and lower, respectively, than that of the type strain.

Effect of acid (A pH 4) and bile (B oxgall 3%) on growth of Lactobacillus plantarum Sanriku-SU7 (S-SU7), Izu-SU2 (I-SU2) and Tennozu-SU2 (T-SU2). Values are expressed as mean ± SEM (n = 4). Values with different superscript letters are significantly different with p < 0.05
The growth capacity of L. plantarum strains in the acidified (pH 4) broth and broth containing a high bile concentration is in agreement with previous reports [28, 37]. Probiotic screening for other LAB species, such as Lactococcus lactis, has determined that some of these strains including their type strain cannot grow in the acidified and/or bile-containing broths [28, 31].
Autoaggregation and Adhesion on HT-29-Luc Cells
Autoaggregation of probiotic strains appears to be necessary for adhesion to intestinal epithelial cells [32]. Aggregation and the adhesion capacity of LAB are regarded as factors for the disruption of adhesion of pathogenic bacteria by competing for binding to intestinal epithelial cells, which consequently reduces pathogen colonisation and prevents infection [38].
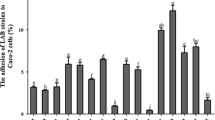
Among the four strains, the autoaggregation was the highest (about 60%) in L. plantarum Tennozu-SU2 (Fig. 2A). The aggregation of L. plantarum Sanriku-SU7 and Izu-SU2 was lower than 10%. In the adhesion assay, the adherent cell count was higher and lower in L. plantarum Izu-SU2 and Tennozu-SU2, respectively, though the difference was not significant (Fig. 2B). This result was not in agreement with result of the autoaggregation assay.

Autoaggregation (A) and adhesion on human enterocyte-like HT-29-Luc cells (B, C) of L. plantarum strains. A, B Values are expressed as mean ± SEM (n = 3). Values with different superscript letters are significantly different with p < 0.05. C A Gram-stained image of HT-29-Luc cells (pale blue) and L. plantarum cells (dark blue) (Colour figure online)
Antagonistic Effects in BHI Broth and HT-29-Luc Cells
S. Typhimurium inoculation size was approximately 3 log CFU/mL BHI broth. None of the four LAB strains could completely suppress the pathogen, which reached 8 log CFU/mL during the 24 h co-incubation. However, the final viable count of the pathogen was reduced to 25% by L. plantarum Tennozu-SU2 and Izu-SU2 (Fig. 3A). This inhibitory effect was higher than that observed for the type strain. Although the antimicrobial activity of various LAB strains with bacteriocin has been reported [39, 40], it is considered that the tested strains have no bacteriocin and have a competitive effect against Salmonella.

Antagonistic effect of L. plantarum strains against Salmonella Typhimurium in BHI broth (A) and adhesion on HT-29-Luc cells (B). Values are expressed as mean ± SEM (n = 4). Values with different superscript letters are significantly different with p < 0.05
A comparison of the effect of heat-killed cells from tested LAB strains on the adhesion and invasion of S. Typhimurium in human enterocyte-like HT-29-Luc cells is shown in Fig. 3B. The levels of pathogen adhesion in intact cells observed in this study, approximately 6 log CFU/well, are in agreement with those in previous reports [33, 41]. Adhesion of S. Typhimurium was clearly inhibited to about 20, 40 and 50% of the control by heat-killed L. plantarum Tennozu-SU2, Izu-SU2 and Sanriku-SU7, respectively. The inhibitory effect was not shown by type strain, though its adhesive activity was not different from other strains (Fig. 3B). The aggregation rate of the type strain was not considerably lower compared with the other tested strains (Fig. 2A).
These results of the in vitro experiments indicate that L. plantarum isolates from the coastal area, rather than the type strain, have an inhibitory effect on the S. Typhimurium infection without a bacteriocin. High molecular compounds, such as some dietary fibres, and heat-killed probiotic cells can affect barrier function as expressed through transepithelial electrical resistance (TEER) [16, 42]. The cell membrane, exopolysaccharides and other heat-treated compounds of the LAB cells in this experiment affect the HT-29-Luc cells and/or S. Typhimurium cells. More detailed studies involving proteomic and genomic experiments to evaluate the effect of intact and heat-treated cell compounds on intestinal cells are needed. From the results of this in vitro experiment, L. plantarum Tennozu-SU2 was selected for the next experiment involving an animal model system.
Effect of Live and Heat-Killed L. plantarum Tennozu-SU2 on the S. Typhimurium Infection in BALB/c Mice
In this animal experiment, all mice survived the entire experimental time course, while faeces were soft in control group after inoculation with the pathogen. Four days after the inoculation, body weight was tended to be high in the mice fed live L. plantarum Tennozu-SU2 cells (Fig. 4A). There was no difference in the liver and spleen weights (data not shown).

Effects of live and heat-killed L. plantarum Tennozu-SU2 cells on drinking water on body weight (A), viable counts in the faeces (B), liver (C) and spleen (D) of S. Typhimurium-infected BALB/c mice. Symbols in A: open circles, open squares and closed squares represent control, live cells and heat-killed cell groups, respectively. Values are expressed as mean ± SEM (n = 6). Values with different superscript letters are significantly different with p < 0.05
Two days after the oral inoculation of 7 log CFU/mouse of S. Typhimurium, 5.0 log CFU/g of Salmonella, expressed as typical black colonies on DHL agar, was detected in faeces of mice that were fed the control diet (Fig. 4B). Subsequently, the viable count of Salmonella cells increased to 6.0 log CFU/g in 4 days. This result suggests that S. Typhimurium colonised the murine intestinal epithelial cells [43]. At 7 days after the inoculation, the Salmonella colonies could not be counted correctly, because there were higher numbers of other Enterobacteriaceae colonies on the DHL agar plate. Both live and heat-killed L. plantarum Tennozu-SU2 cells in drinking water suppressed this colonisation slightly and approximately eightfold at 2 and 4 days.
In all of the mice, Salmonella was detected in the liver tissues. Approximately 4.8 log CFU/g tissue of Salmonella was recovered from the liver of control mice (Fig. 4C). Live and heat-killed L. plantarum Tennozu-SU2 cells reduced the pathogenic counts to 4.4 and 4.6 log CFU/g tissue, respectively. The count in mice fed heat-killed cells differed from counts in other groups; however, the difference was small.
The spleen of mice from all the three groups was also found to be infected by S. Typhimurium. The viable count of S. Typhimurium in the spleen of control mice was approximately 3.9 log CFU/g tissue. The viable counts in the tissues of mice that were fed live or heat-killed L. plantarum Tennozu-SU2 cells were lower, about 3.5 log CFU/g (Fig. 4D). These results are consistent with those obtained from the differentiated HT-29-Luc cells (Fig. 3A, B).
As mentioned earlier, the inhibitory effects of several species of LAB, particularly L. acidophilus and L. casei, on S. Typhimurium infection in mice are known [21–24]. However, the inhibitory effect of L. plantarum on S. Typhimurium infection in mice had been not clear. The results from in vitro and in vivo experiments conducted during this study indicate that both live and heat-killed L. plantarum Tennozu-SU2 cells have an inhibitory effect similar to that of those previous reports about probiotic L. acidophilus and L. casei on S. Typhimurium infection. It is known that L. plantarum has high tolerance against acid, bile and salinity and has activities associated with immune and metabolic improvement [13, 14, 19, 20, 29]. To clarify the mechanisms, future studies concerning immune-related cytokines from enterocyte cultures and in vivo experiments are needed. Thus, we plan to study the synergistic effects of L. plantarum Tennozu-SU2 with other dietary compounds, such as functional dietary fibres, peptides and fatty acids.
Conclusion
In this study, the probiotic properties and inhibitory effect on S. Typhimurium adhesion on human enterocyte-like HT-29-Luc cells of three L. plantarum strains isolated from fermented fish, beach sand and a coastal plant were determined. Compared with the type strain, L. plantarum Tennozu-SU2 isolated from the acorn of a coastal tree showed high autoaggregation in MRS broth and an antagonistic effect against S. Typhimurium in BHI broth. Furthermore, heat-killed L. plantarum Tennozu-SU2 cells inhibited S. Typhimurium adhesion on HT-29-Luc cells. Both live and heat-killed cells showed inhibitory effect on gut colonisation of BALB/c mice, as assessed by a viable Salmonella count in faecal samples, and invasion into liver and spleen tissues. The properties shown in this study suggest that L. plantarum Tennozu-SU2 is useful as a starter and probiotic bacteria in functional food material.
References
Galis AM, Marcq C, Marlier D, Portetelle D, Van I, Beckers Y, André T (2013) Control of Salmonella contamination of shell eggs—preharvest and postharvest methods. Comp Rev Food Sci Food Safety 12:155–182
Zaki HMBA, Mohamed HMH, El-Sheriff AMA (2015) Improving the antimicrobial efficacy of organic acids against Salmonella enterica attached to chicken skin using SDS with acceptable sensory quality. LWT-Food Sci Technol 64:558–564
Gkana E, Lianou A, Nychas GJE (2016) Transfer of Salmonella enterica serovar Typhimurium from beef to tomato through kitchen equipment and the efficacy of intermediate decontamination procedures. J Food Prot 79:1252–1258
Henao-Herreño LX, López-Tamayo AM, Ramos-Bonilla JP, Haas CN, Husserl J (2016) Risk of illness with Salmonella due to consumption of raw unwashed vegetables irrigated with water from the Bogotá River. Risk Anal. doi:10.1111/risa.12656
Kuda T, Yazaki T, Takahashi H, Kimura B (2013) Effect of dried and vinegar flavored squid products on acid resistance of Salmonella Typhimurium and Staphylococcus aureus. Food Control 30:569–574
Gorman R, Adley CC (2004) Characterization of Salmonella enterica serotype Typhimurium isolates from human, food, and animal sources in the Republic of Ireland. J Clin Microbiol 42:2314–2316
Hernandez SM, Keel K, Sanchez S, Trees E, Gerner-Smidt P, Adams JK, Cheng Y, Ray A 3rd, Martin G, Presotto A, Ruder MG, Brown J, Blehert DS, Cottrell W, Maurer JJ (2012) Epidemiology of a Salmonella enterica subsp. enterica serovar Typhimurium strain associated with a songbird outbreak. Appl Environ Microbiol 78:7290–7298
Fàbrega A, Vila J (2013) Salmonella enterica serovar Typhimurium skills to succeed in the host: virulence and regulation. Clin Microbiol Rev 26:308–341
Kuda T, Shibata G, Takahashi H, Kimura B (2015) Effect of quantity of food residues on resistance to desiccation of food-related pathogens adhered to a stainless steel surface. Food Microbiol 46:234–238
Kuda T, Koyanagi T, Shibata G, Takahashi H, Kimura B (2016) Effect of carrot residue on the desiccation and disinfectant resistances of food related pathogens adhered to a stainless steel surfaces. LWT-Food Sci Technol 74:251–254
Ryan D, Pati NB, Ojha UK, Padhi C, Ray S, Jaiswal S, Singh GP, Mannala GK, Schultze T, Chakraborty T, Suar M (2015) Global transcriptome and mutagenic analyses of the acid tolerance response of Salmonella enterica serovar Typhimurium. Appl Environ Microbiol 81:8054–8065
Leekitcharoenphon P, Henderiksen RS, Hello SL, Well FX, Baggesen DL, Jun SR, Ussery DR, Lund O, Crook DW, Wilson DJ, Aerestrup FM (2016) Global genomic epidemiology of Salmonella enterica serovar Typhimurium DT104. Appl Environ Microbiol 82:2516–2526
Nemoto M, Kuda T, Eda M, Yamakawa H, Takahashi H, Kimura B (2016) Protective effects of mekabu aqueous solution fermented by Lactobacillus plantarum Sanriku-SU7 on human enterocyte-like HT-29-luc cells and DSS-induced murine IBD model. Probiotics Antimicro Prot . doi:10.1007/s12602-016-9226-x in print
Eda M, Kuda T, Kataoka M, Takahashi H, Kimura B (2016) Anti-glycation properties of the aqueous extract solutions of dried algae products harvested and made in the Miura Peninsula, Japan, and effect of lactic acid fermentation on the properties. J Appl Phycol . doi:10.1007/s10811-016-0891-7 in print
Kuda T, Sarengaole TH, Kimura B (2016) Alcohol-brewing properties of acid- and bile-tolerant yeasts co-cultured with lactic acid bacteria isolated from traditional handmade domestic dairy products from Inner Mongolia. LWT-Food Sci Technol 63:62–69
Liu WH, Yang CH, Lin CT, Li SW, Cheng WS, Jiang YP, Wu CC, Chang CH, Tsai YC (2015) Genome architecture of Lactobacillus plantarum PS128, a probiotic strain with potential immunomodulatory activity. Gut Pathogens 7:22
Xie J, Yu Q, Nie S, Fan S, Xiong T, Xie M (2015) Effects of Lactobacillus plantarum NCU116 on intestine mucosal immunity in immunosuppressed mice. J Agric Food Chem 63:10914–10920
Tang Y, Dong W, Wan K, Zhang L, Li C, Zhang L, Liu N (2015) Exopolysaccharide produced by Lactobacillus plantarum induces maturation of dendritic cells in BALB/c mice. PLoS One 10:e0143743
Nishimura M, Ohkawara T, Tetsuka K, Kawasaki Y, Nakagawa R, Satoh Y, Sato Y, Nishihara J (2016) Effects of yogurt containing Lactobacillus plantarum HOKKAIDO on immune function and stress markers. J Tradit Complement Med 6:275–280
Xie J, Nie S, Yu Q, Yin J, Xiong T, Gong D, Xie M (2016) Lactobacillus plantarum NCU116 attenuates cyclophosphamide-induced immunosuppression and regulates Th17/Treg cell immune responses in mice. J Agric Food Chem 64:1291–1297
Castillo NA, Perdigón G, de LeBlanc AM (2011) Oral administration of a probiotic Lactobacillus modulates cytokine production and TLR expression improving the immune response against Salmonella enterica serovar Typhimurium infection in mice. BMC Microbiol 11:77
Nakazato G, Paganelli FL, Lago JC, Aoki FH, Mobilon C, Brocchi M, Stehling EG, Silveira WD (2011) Lactobacillus acidophilus decrease Salmonella Typhimurium invasion in vivo. J Food Safety 31:284–289
Chen CY, Tsen HY, Lin CL, Lin CK, Chuang LT, Chen CS, Chiang YC (2013) Enhancement of the immune response against Salmonella infection of mice by heat-killed multispecies combinations of lactic acid bacteria. J Med Microbiol 62:1657–1664
Llana MN, Sarnacki SH, Castañeda MRA, Bernal MI, Giacomodonato BMN, Cerquetti MC (2013) Consumption of Lactobacillus casei fermented milk prevents Salmonella reactive arthritis by modulating IL-23/IL-17 expression. PLoS One 8:e82588
Eom JS, Song J, Choi HS (2015) Protective effects of a novel probiotic strain of Lactobacillus plantarum JSA22 from traditional fermented soybean food against infection by Salmonella enterica serovar Typhimurium. J Microbiol Biotechnol 25:479–491
Kanno T, Kuda T, An C, Takahashi H, Kimura B (2012) Radical scavenging capacities of saba-narezushi, Japanese fermented chub mackerel, and its lactic acid bacteria. LWT-Food Sci Technol 47:25–30
Kuda T, Kawahara M, Nemoto M, Takahashi K, Kimura B (2014) In vitro antioxidant and anti-inflammation properties of lactic acid bacteria isolated from fish intestines and fermented fish from the Sanriku Satoumi region in Japan. Food Res Int 64:248–255
Kawahara M, Nemoto M, Nakata T, Kondo S, Takahashi H, Kimura B, Kuda T (2015) Anti-inflammatory properties of fermented soy milk with Lactococcus lactis subsp. lactis S-SU2 in murine macrophage RAW264.7 cells and DSS-induced IBD model mice. Int Immunopharmacol 26:295–303
Kuda T, Masuko Y, Kawahara M, Kondo S, Nemoto M, Nakata T, Kataoka M, Takahashi H, Kimura B (2016) Bile acid-lowering properties of Lactobacillus plantarum Sanriku–SU3 isolated from Japanese surfperch fish. Food Biosci 14:41–46
Kuda T, Kataoka M, Nemoto M, Kawahara M, Takahashi H, Kimura B (2016) Isolation of lactic acid bacteria from plants of the coastal Satoumi regions for use as starter cultures in fermented milk and soymilk production. LWT-Food Sci Technol 68:202–207
Kuda T, Noguchi Y, Ono M, Takahashi H, Kimura B, Kamita R, Eto T, Kato M, Kawahara M (2014) In vitro evaluation of the fermentative, antioxidant, and anti-inflammation properties of Lactococcus lactis subsp. lactis BF3 and Leuconostoc mesenteroides subsp. mesenteroides BF7 isolated from Oncorhynchus keta intestines in Rausu, Japan. J Funct Foods 11:269–277
Kos B, Šušković J, Vuković S, Šimpraga M, Frece J, Matošić S (2003) Adhesion and aggregation ability of probiotic strain. Lactobacillus acidophilus M92. J Appl Microbiol 94:981–987
Kuda T, Kosaka M, Hirano S, Kawahara M, Sato M, Kaneshima T, Nishizawa M, Takahashi H, Kimura B (2015) Effect of sodium-alginate and laminaran on Salmonella Typhimurium infection in human enterocyte-like HT-29-Luc cells and BALB/c mice. Carbohydr Polymers 125:113–110
Kuda T, Tsuda N, Yano T (2004) Thermal inactivation characteristics of acid and alkaline phosphatase in fish and shellfish. Food Chem 88:543–548
Dimitrov Z, Gotova I, Chorbadjiyska E (2014) In vitro characterization of the adhesive factors of selected probiotics to Caco-2 epithelium cell line. Biotechnol Biotechnol Equip 28:1079–1083
Nakamura S, Kuda T, An C, Kanno T, Takahashi H, Kimura B (2012) Inhibitory effects of Leuconostoc mesenteroides 1RM3 isolated from narezushi, a fermented fish with rice, on Listeria monocytogenes infection to Caco-2 cells and A/J mice. Anaerobe 18:19–24
Giraud E, Lelong B, Raimbaut M (1991) Influence of pH and initial lactate concentration on the growth of Lactobacillus plantarum. Appl Microbiol Biotechnol 36:96–99
Frece J, Kos B, Svetec IK, Zgaga Z, Beganović J, Lebos A, Susković J (2009) Synbiotic effect of Lactobacillus helveticus M92 and prebiotics on the intestinal microflora and immune system of mice. J Dairy Res 76:98–104
Dobson A, Catter PD, Ross PR, Hill C (2012) Bacteriocin production: a probiotic trait? Appl Environ Microbiol 78:1–6
Abedi D, Feizizadef S, Akbari V, Jafarian-Dehkordi A (2013) In vitro anti-bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp bulgaricus on Escherichia coli. Res Pharm Sci 8:260–268
Gagnon M, Berner AZ, Chervet N, Chassard C, Lacroix C (2013) Comparison of the Caco-2, HT-29 and the mucus-secreting HT29-MTX intestinal cell models to investigate Salmonella adhesion and invasion. J Microbiol Methods 94:274–279
Liu HY, Rooms S, Jonsson H, Ahi D, Dicksved J, Lindberg JE (2015) Effects of Lactobacillus johnsonii and Lactobacillus reuteri on gut barrier function and heat shock proteins in intestinal porcine epithelial cells. Physiol Rep 3:e12355
Drago-Serrano ME, Rivera-Aguilar V, Reséndiz-Albor AA, Campos-Rodríguez R (2010) Lactoferrin increases both resistance to Salmonella typhimurium infection and the production of antibodies in mice. Immun Lett 134:35–46
Acknowledgements
The present study was supported in part by the Terrada Warehouse Company, Tokyo, Japan.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The animal experiments were performed in compliance with the Fundamental Guidelines for Proper Conduct of Animal Experiment and Related Activities in Academic Research Institutions, under the jurisdiction of the Ministry of Education, Culture, Sports, Science and Technology, and approved by the animal experiment committee of Tokyo University of Marine Science and Technology (approval no. H28-4).
Conflict of Interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Hirano, S., Yokota, Y., Eda, M. et al. Effect of Lactobacillus plantarum Tennozu-SU2 on Salmonella Typhimurium Infection in Human Enterocyte-Like HT-29-Luc Cells and BALB/c Mice. Probiotics & Antimicro. Prot. 9, 64–70 (2017). https://doi.org/10.1007/s12602-016-9243-9
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12602-016-9243-9