
Abstract
This study was to evaluate the effect of a preparation of Bacillus probiotic (Bacillus licheniformis and B. subtilis, 1:1) on growth and survival rate of Pacific white shrimp, Litopenaeus vannamei larvae. The larvae were fed on Artemia urmiana nauplii and Brachionus plicatilis enriched with the probiotic preparation at 1 × 106 CFU mL−1 rate. The experimental setup was completely randomized design comprised of six treatments, namely solo Artemia nauplii (A) or rotifer (R), Artemia nauplii and rotifer without any enrichment (A + R), Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), rotifer enrichment with probiotic bacilli (R + B) and enriched Artemia nauplii and rotifer (A + R + B). All treatments were performed in triplicate. Chemical parameters of rearing water viz. pH, salinity and temperature were 7.5–8, 30–31 ppt and 31–32 °C, respectively. Photoperiod was 16L:8D. Shrimp larvae were fed Artemia nauplii and rotifers at 5–20 and 10–40 individuals per shrimp larvae four times a day, respectively. Growth and survival rate of larvae were determined at MII, MIII, PL1, PL4, PL7 and PL10 stages. Larvae in A + R + B treatment showed the highest total length (10.89 ± 0.51 mm), weight (674 ± 73 μg) and survival rate (65 % ± 3.5). Lowest total length, weight and survival rate (7.96 ± 0.63 mm, 493 ± 52 μg and 24.5 ± 2.4 %, respectively) were recorded in treatment B larvae. We concluded that Bacillus probiotic can improve growth and survival rate of Pacific white shrimp larvae without conceivably undesirable effects.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Pacific white shrimp, Litopenaeus vannamei, has become one of the most widely cultivated crustacean species worldwide, with production exceeding that of the black tiger shrimp, Penaeus monodon, since 2003 [1]. This is largely due to the advantages in terms of disease management and strain selection. The sustained growth of shrimp aquaculture demands high numbers of good quality hatchery-reared postlarvae and juveniles, especially where captive broodstocks are used. Similar to penaeids, the larval L. vannamei undergoes complicated ontogenic feeding habits [2, 3]. The non-feeding nauplii are followed by the phytoplankton-feeding zoeae and the carnivorous mysis stage which is largely dependent upon a perpetual provision of live food, preferably rotifers and Artemia nauplii.
Intensification is considered the main driving factor for repeated diseases outbreaks in shrimp farms. There is a growing concern regarding extensive application of antibiotics including spread of drug-resistant pathogens and hazards on the food chain and the environment. Innovative aquaculture necessitates keeping healthy environment for the sake of good production practices [4]. The potential of probiotics to replace antibiotics has recently been revealed regarding their growth promoting features including their capacity to halt proliferation of pathogenic microorganisms and other nutritional benefits. The European Union (Regulation (EC) 83/2003) has legalized the use of antibiotics as growth promoters for shrimp producing and exporting farms owing to fears of development of antibiotic resistant strains by the year 2006 [5]. Probiotics can be regarded as live microbes providing a beneficial effect on the host through amending feed utilization capability of host or enhancing nutritional value of feed, stimulating the immune system or even by ameliorating rearing water quality [6, 7]. As a prominent probiont, bacilli are Gram positive, spore forming bacteria used as commercial probiotics mostly owing to the physical and biological characteristics of their spore conferring their convenient storage and cost-effectiveness [6, 8, 9]. Bacillus can positively affect cultured organisms via several modes of action including enhancing survival and growth [10], stimulating the digestive [11] and immune systems [6] and also bioremediating [12, 13].
Several studies investigated the beneficial effects of probiotic bacilli on the larval rearing, and more specifically on survival of the white shrimp: Bacillus firmus ZOU4 [14], Bacillus cereus ZOU8 [15], Bacillus fusiformis [16], Bacillus subtilis [17], Bacillus subtilis E20 [18], several strains of Bacillus spp., including Bacillus licheniformis and Bacillus subtilis [19–21]. The objective of this study was to determine the effect of probiotic bacilli (Bacillus licheniformis and Bacillus subtilis) on the growth, survival rate and microbial number of L. vannamei during larval and postlarval stages fed live food (Artemia urmiana and Brachionus plicatilis).
Materials and Methods
Experimental Design and Larval Rearing System
The study was conducted in Gomishan shrimps research center (Gomishan, Golestan province, Iran). Nauplii of L. vannamei and Chaetoceros muelleri alga were generously provided by the Gomishan shrimp production hatchery. Artemia (A. urmiana), Rotifer (Brachionus plicatilis) and Isochrysis galbana alga were also generously obtained from Urmia Lake research center (Urmia, Iran). Nauplii of L. vannamei were aerobically acclimatized at a density of 100 L−1 in 500-L fiber glass tanks with 30 ppt seawater disinfected with 10 mg L−1 of chlorine until reaching the MI. Prior to the commencing ofexperiments, nauplii were fed with microalgae, Chaetoceros muelleri and Isochrysis galbana at a rate of 106 cell mL−1 each day. The daily water exchange rate was 10 % through water flow rate of 1 L min−1. Weight and length of larvae were also measured before the start of experiment.
This experiment evaluated the effect of Artemia nauplii and rotifer enriched with probiotic and also without any enrichment on larval stages from mysis 2 to PL10. The experiment was performed as a completely randomized design with six treatments including sole Artemia nauplii (A) or rotifer (R), Artemia nauplii and rotifer without any enrichment (A + R), Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), rotifer enrichment with probiotic bacilli (R + B) and enriched Artemia nauplii and rotifer (A + R + B).
Each tank was stocked with mysis stage 1 larvae (N = 1500; 30 mysis L−1). Each treatment was randomly assigned to triplicate glass tanks with a capacity of 50 L. Shrimp larvae were fed Artemia nauplii and rotifers at 5–20 Artemia nauplii and 10–40 rotifers (Table 1) per larvae, four times a day at 6:00, 12:00, 18:00 and 24:00. Salinity and temperature of culture water were maintained at 30–31 ppt and 31–32 °C, respectively. Growth and survival rate determinations and microbial enumeration were carried out at stages MII, MIII, PL1, PL4, PL7 and PL10 (Table 2).
Preparation of the Probiotic Bacilli
The commercial probiotic used in this study (Protexin Aquatech, Probiotics International Ltd, Somerset TA146QE, United Kingdom) contained spores of two species of Bacillus (Bacillus licheniformis and B. subtilis). The spores were rehydrated to vegetative bacteria according to manufacturer’s instructions [11]. Subsequently, the blend of probiotic bacilli were added to Artemia and rotifer culture medium at a density of 1 × 106 CFU mL−1.
Rotifer Enrichment
Brachionus plicatilis was cultured at temperature 26 °C and salinity 24 ppt in 1.0 m3 plastic containers and fed with Chaetoceros muelleri and Isochrysis galbana. Algae were cultured using Guillard f/2 [22] for Chaetoceros muelleri and Conway [23] for Isochrysis galbana, under a standard ambient condition (temperature 22–30 °C, salinity 26–36 ppt, pH 7–9 and light 5000–10,000 lux).
Rotifers were taken from the culture tank, rinsed in seawater and transferred to small glasses (1.5 L) supplied with gentle aeration. The water quality parameters were similar to the previous stage. The probiotic preparation was added at a density of 1 × 106 CFU mL−1 to the containers right after rotifers were transferred. The enrichment was undertaken for 8 h. Samples of rotifers were concentrated on a 55-μm mesh filter and carefully washed with seawater. Afterward, the samples were gently rinsed in distilled water, transferred to Eppendorfs and frozen immediately in liquid nitrogen. Samples were kept at −80 °C until further analysis.
Artemia nauplii Enrichment
Artemia urmiana cysts were hatched at a density of 1.5 g L−1, in natural filtered (55 μm) seawater at a salinity of 30 ppt and 26 °C with strong aeration and illumination. After 24 h, Artemia nauplii were separated, rinsed with seawater and transferred to small containers (1.5 L). Water quality parameters were similar to those applied to rotifers. The nauplii were enriched for 8 h following the same procedure as for the rotifers.
Microbial Enumeration
Water samples, digestive tract samples from shrimp and enriched Artemia and Rotifer were taken to determine counts of total bacteria and probiotic Bacillus. Prior to the homogenization, the shrimp larvae, Rotifer and Artemia nauplii were rinsed with sterilized distilled water, washed with 0.1 % benzalkonium chloride according to Gatesoupe [6], and then rinsed again with sterilized distilled water to ensure all external bacteria were removed. All samples were diluted serially with sterilized normal saline solution (0.85 % w/v NaCl). Total counts of bacteria were determined by plating on tryptic soy agar (with 1 % w/v NaCl), according to Shariff et al. [24]. Bacillus bacteria in water samples were also cultured using a surface drop technique with yeast extract agar. Digestive tract samples were cultured on Bacillus cereus agar (Oxoid CM617) according to the method recommended by Probiotics International Ltd. (Protexin Aquatech, Registration Dossier, unpublished pamphlet). The number of colonies on each plate was counted after incubation for 68 h at 25 °C for water samples and for 72 h at 37 °C for digestive tract samples.
Biochemical Analyses
Dry matter contents of shrimp larvae and live feeds were determined after oven-drying to constant weight at 105 °C [25]. Protein content was determined by the method of Lowry [26], after hydrolysis with NaOH 1.0 M at 95 °C. Carbohydrate was determined by the phenol/sulfuric acid method [27], and lipid was quantified by carbonization at 200 °C [28].
Statistical Analysis
Data on final body weight (BW), survival rate and whole body proximate composition were analyzed using one-way ANOVA (Systat Software Inc., Chicago, IL, USA). Significant differences among treatments were evaluated with Tukey HSD tests [29]. Statistical significance of differences was determined by setting the probability at 5 % (P < 0.05) for each set of comparisons.
Results
Bacterial Study
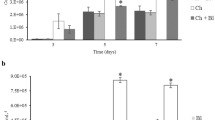
Total count of CFU in Rotifer and Artemia nauplii were determined 2, 4, 6 and 8 h postaddition of probiotic bacilli. Bacterial count was increased as time went by in both Rotifer and Artemia nauplii. The mean Bacillus count in probiotic-enriched Rotifer and Artemia nauplii was 0.12 ± 0.005 × 104 and 0.4 ± 0.02 × 104 CFU Ind−1, respectively (Fig. 1).

Concentration of probiotic Bacillus allocation to the A. urmiana nauplii and Rotifers in different enrichment time (triangle Broth: 1 × 106 CFU mL−1, diamond Artemia and square Rotifers: 1 × 104 CFU Ind−1)
In larval shrimp digestive tract, the total bacterial and probiotic Bacillus increased from 3.6 to 6.7 and 1.1 to 5.1 log CFU larvae−1, respectively, from mysis 2 to PL1 in all groups; however, the total bacterial count and probiotic Bacillus did not increased from PL1 to PL10 (Fig. 2). In shrimp larvae, total bacterial counts of probiotic treatments (A + B, R + B and A + R + B) did not significantly differ from total bacterial counts of other treatments (A, R and A + R) at all larval stages. Number of probiotic Bacillus of probiotic treatments (A + B, R + B and A + R + B) were not significantly different at all larval stages.

Total bacterial count and Bacillus count in digestive tracts of L. vannamei.). First bar Artemia nauplii (A), second bar rotifer (R), third bar Artemia nauplii and rotifer without any enrichment (A + R), fourth bar Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), fifth bar rotifer enrichment with probiotic bacilli (R + B) and sixth bar enriched Artemia nauplii and rotifer (A + R + B)
Survival and Growth
There was significant difference among larvae fed enriched live food containing probiotic bacilli (A. urmiana nauplii and B. plicatilis) and those supplied with unenriched live food regarding larval survival rate (P > 0.05). The Pacific white shrimp, L. vannamei, larvae fed A + R + B treatment showed the highest survival rate (65 %) compared to others (Fig. 3). Lowest survival rate was recorded in treatment fed B. plicatilis alone (24 %).

Survival rate of L. vannamei fed with live food (A. urmiana nauplii and B. plicatilis). open diamond Artemia nauplii (A), closed square rotifer (R), closed triangle Artemia nauplii and rotifer without any enrichment (A + R), closed diamond Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), open square rotifer enrichment with probiotic bacilli (R + B) and open triangle enriched Artemia nauplii and rotifer (A + R + B)
Total length and weight of postlarvae were highest in A + R + B (10.89 mm and 674 μg) and A + R (10.56 mm and 650 μg). There was no significant difference in the L. vannamei larval total length and weight recorded between larvae fed with enriched live foods and unenriched live foods (P > 0.05). Lowest total length (7.96 mm) and weight (493 μg) were recorded in R treatment when larvae fed with B. plicatilis alone (Figs. 4, 5, 6).

Total length (mm) of L. vannamei fed with live food (A. urmiana nauplii and B. plicatilis). Closed square Artemia nauplii (A), open square rotifer (R), open triangle Artemia nauplii and rotifer without any enrichment (A + R), open diamond Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), closed triangle rotifer enrichment with probiotic bacilli (R + B) and closed diamond enriched Artemia nauplii and rotifer (A + R + B)

Total weight (μg) of L. vannamei fed with live food (A. urmiana nauplii and B. plicatilis). First bar Artemia nauplii (A), second bar rotifer (R), third bar Artemia nauplii and rotifer without any enrichment (A + R), fourth bar Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), fifth bar rotifer enrichment with probiotic bacilli (R + B) and sixth bar enriched Artemia nauplii and rotifer (A + R + B)

Biochemical composition (%) of L. vannamei fed with live food (A. urmiana nauplii and B. plicatilis). First bar Artemia nauplii (A), second bar rotifer (R), third bar Artemia nauplii and rotifer without any enrichment (A + R), fourth bar Artemia nauplii enrichment with probiotic bacilli (Bacillus licheniformis and B. subtilis) (A + B), fifth bar rotifer enrichment with probiotic bacilli (R + B) and sixth bar enriched Artemia nauplii and rotifer (A + R + B)
Biochemical Analyses
There was no significant difference in protein, lipid and carbohydrate content of the L. vannamei larval stage MII and PL10 between larvae fed with enriched live food with probiotic bacilli and unenriched live food (P > 0.05). The mean protein, lipid and carbohydrate level in L. vannamei larval stage MII fed with live foods was between 54.6 and 56.2 %, 19.6 and 21.3 %, 6.8 and 8.71 % in different treatment, respectively. In shrimp larvae, the protein, lipid and carbohydrate level decreased from 55.1 to 39.8, 20.6 to 14.1 and 7.7 to 5.05 %, respectively, on larval stages MII to PL10 in all treatments. However, dry matter level increased in all treatments. The protein, lipid and carbohydrate level of L. vannamei larval stage PL10 was highest in treatments fed with Artemia nauplii alone or mixture with Rotifer than treatments fed with Rotifer alone.
Discussion
The survival rate, total length and weight of Litopenaeus vannamei of treatment A + R + B were significantly better in comparison with other treatments. The same results were obtained in Macrobrachium rosenbergii fed with Artemia and Artemia and B. plicatilis (70 + 30 %). Larvae fed Artemia alone showed better survival, MLS and growth and took shorter time to reach the next stage [30]. This is attributed to higher lipid content and consequently the caloric value of Artemia than rotifers [31, 32], besides the presence of higher levels of n-3 HUFA’s in Artemia [33].
Blair et al. [34] reported that larval haddock (Melanogrammus aeglefinus) fed on Artemia nauplii showed better length and weight comparing to those fed rotifer, mainly due to differences in prey size. Since rotifers are smaller than Artemia nauplii, the larvae consuming rotifers would have to spend additional energy to capture a greater number of individual prey organisms before reaching satiation. However, higher lipid and energy content of Artemia would result in such differences. It is apparent that the nutrients provided by a rotifer diet were generally inadequate to meet the nutritional requirements of mud crab larvae, S. serrate, to sustain its survival and development up to the megalopa stage [35]. Godfred et al. [36] reported that in the portunid crab Thalamita crenata, Z1–Z2 larvae had highest survival when fed with B. plicatilis alone, but later stages (Z3–Z5) showed better survival and development on an Artemia–rotifer combination. In addition to prey nutritional value, its size and type of movements would affect its ingestability, feeding success (successful bites to total bites) and foraging efficiency (energy gained from prey to energy cost of food uptake) of the predator [35, 37]. It is also worth mentioning that fish and crustacean larvae have a poor digestive capacity, probably due to an insufficient enzymatic activity and short gut retention time particularly in carnivorous larvae [38–40].
Application of probiotic in the culture water or via Artemia yielded better survival rates [6, 11, 41, 42]. Feeding shrimps with enriched Artemia and rotifers to shrimp [11] and fish [43] is an effective means to deliver the probiotic to host. The results showed that total bacterial and probiotic Bacillus increased from 3.6 to 6.7 and 1.1 to 5.1 log CFU larvae−1, respectively, from larval stages mysis 2 to PL. This gradual increase implied their colonization in the digestive tract. Ziaei-Nejad et al. [11] revealed Bacillus numbers at 104–105 CFU larva−1 (61.5–93.0 % of total bacterial flora) when F. indicus larvae were inoculated with a commercial probiotic product containing 106 CFU mL−1 of Bacillus via culture water or enriched Artemia. In P. monodon, Bacillus, when used as a probiotic, was able to colonize both the culture water and the shrimp digestive tract; the Bacillus also was able to replace Vibrio spp. in the gut of the shrimp, thereby increasing shrimp survival [43], via out-competing other bacteria for nutrients and space by producing antibiotics [7, 13]. It has also been shown to increase shrimp survival by enhancing resistance to pathogens through activating both cellular and humoral immune defenses in shrimp [44, 45]. The results showed that enrichment with Bacillus bacteria did not improve the growth performance of L. vannamei, but a tendency of better values was observed in the experimental group A + R + B for survival. Silva et al. [21] reported that for postlarvae phase, the probiotic did not influence zootechnical parameters (wet weight, total length and survival); however, there is a reduction in presumptive Vibrio counts both in the water and in the shrimp, providing greater security of the system. Ziaei-Nejad et al. [11] also reported that the weight of F. indicus in stages N1-2 to Z3 did not differ significantly when cultured with or without probiotic; however, higher final weight was observed in shrimp treated with probiotic from M1 to PL1-2. Similarly, it is depicted that treating P. monodon and Litopenaeus vannamei with a commercial Bacillus probiotic did not significantly increase the growth [21, 25, 46]. It must be mentioned that many factors such as species composition, application level, frequency of application and environmental conditions would more or less affect the outcome of such studies [10]. The results revealed that protein, lipid and carbohydrate content of the L. vannamei larvae fed enriched preys and those received unenriched preys did not differ. Similarly, Yu et al. [47] reported that the body composition of L. vannamei shrimp did not differ significantly when cultured with or without probiotic. The present result is in disagreement with those reported by Venkat et al. [48], Saad et al. [49] and Fernandez et al. [50].
In conclusion, the results demonstrate that combination of Artemia nauplii and B. plicatilis was an appropriate prey for Litopenaeus vannamei larvae from nutritional requirements point of view. This study also examined the efficiency of the use of probiotic via Artemia nauplii and B. plicatilis, with subsequent satisfactory results for transfer of probiotic to shrimp; however, probiotic did not affect growth parameters, but had beneficial effects on the survival of L. vannamei larvae.
References
FAO (2012) Fishery and aquaculture statistics. Food and Agriculture Organization of the United Nations, Rome
Jones DA, Kumlu M, Le Vay L, Fletcher DJ (1997) The digestive physiology of herbivorous, omnivorous and carnivorous crustacean larvae: a review. Aquaculture 155:285–295
Le Vay L, Jones DA, Puello-Cruz AC, Sangha RS, Ngamphongsai C (2001) Digestion in relation to feeding strategies exhibited by crustacean larvae. Comp Biochem Physiol A 128:623–630
Ai Q, Xu H, Mai K, Xu W, Wang J, Zhang W (2011) Effects of dietary supplementation of Bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker, Larimichthys crocea. Aquaculture 317:155–161
Perreten V (2003) Use of antimicrobials in food-producing animals in Switzerland and the European Union (EU). Mitt Lebensmittel Hyg 94:155–163
Gatesoupe FJ (1999) The use of probiotics in aquaculture. Aquaculture 180:147–165
Verschuere L, Rombaut G, Sorgeloos P, Verstraete W (2000) Probiotics bacteria as biological control agents in aquaculture. Microbiol Mol Biol Rev 64:655–671
Gatesoupe FJ (2008) Updating the importance of lactic acid bacteria in fish farming: natural occurrence and probiotic treatments. J Mol Microb Biotech 14:107–114
Wang YB, Tian ZQ, Yao JT, Li WF (2008) Effect of probiotics, Enteroccus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture 277:203–207
Gomez-Gil B, Roque A, Turnbull JF (2000) The use and selection of probiotic bacteria for use in the culture of larval aquatic organisms. Aquaculture 191:259–270
Ziaei-Nejad S, Rezaei MH, Takami GA, Lovett DL, Mirvaghefi AR, Shakouri M (2006) The effect of Bacillus spp. bacteria used as probiotics on digestive enzyme activity, survival and growth in the Indian white shrimp Fenneropenaeus indicus. Aquaculture 252:516–524
Kennedy SB, Tucker JW, Thoresen M, Sennett DG (1998) Current methodology for the use of probiotic bacteria in the culture of marine fish larvae. Aquaculture 98. World Aquaculture Society, Baton Rouge
Moriarty DJW (1998) Control of luminous Vibrio species in penaeid aquaculture ponds. Aquaculture 164:351–358
Wen C, He H, Xue M, Liu H, Zhou Sh (2006) Effects of Bacillus firmus on metamorphosis of Litopenteus vannamei larvae. J Trop Ocean 25:54–58
Wen C, Xue M, He H, Liu HL, Zhou SN (2007) Effects of bacterium Bacillus cereus on metamorphosis of white leg shrimp Litopenaeus vannamei larvae. Fish Sci 26:440–444
Guo J, Liu K, Cheng S, Chang C, Lay J, Hsu Y, Yang J, Chen T (2009) Selection of probiotic bacteria for use in shrimp larviculture. Aquac Res 40:609–618
Lu J, Li H, Huang X (2010) Effects of Bacillus subtilis on water quality and immunization indicators of larval Penaeus vannamei. South China Fish Sci 6:19–24
Liu KF, Chiu CH, Shiu YL, Cheng W, Liu CH (2010) Effects of the probiotic, Bacillus subtilis E20, on the survival, development, stress tolerance, and immune status of white shrimp, Litopenaeus vannamei larvae. Fish Shellfish Immunol 28:837–844
Luis-Villasenor IE, Macias-Rodriguez ME, Gomez-Gil B, Ascencio-Valle F, Campa-Cordova AI (2011) Beneficial effects of four Bacillus strains on the larval cultivation of Litopenaeus vannamei. Aquaculture 321:136–144
Nimrat S, Suksawat S, Boonthai T, Vuthiphandchai V (2012) Potential Bacillus probiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeus vannamei). Vet Microbiol 159:443–450
Silva EF, Soares MA, Calazans NF, Vogeley JL, Valle BC, Soares R, Peixoto S (2013) Effect of probiotic (Bacillus spp.) addition during larvae and postlarvae culture of the white shrimp Litopenaeus vannamei. Aquac Res 44:13–21
Guillard RRL (1975) Culture of phytoplankton for feeding marine invertebrates. In: Smith WL, Chanley MH (eds) Culture of marine invertebrate animals. Plenum Press, New York, pp 26–60
Walne PR (1966) Experiments in the large scale culture of the larvae of Ostrea edulis. Fish Invest Lond 25:1–53
Shariff M, Yusoff FM, Devaraja TN, Srinivasa Rao SP (2001) The effectiveness of a commercial microbial product in poorly prepared tiger shrimp, Penaeus monodon (Fabricius), ponds. Aquac Res 32:181–187
Azewedo PA, Leeson S, Cho CY, Bureau DP (2004) Growth and feed utilization of size rainbow trout (Onchorhynchus mykiss) and Atlantic salmon (Salmo salar) reared in fresh water: diet and species effects, and responses over time. Aquac Nutr 10:401–411
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with Folin-phenol reagent. J Biol Chem 193:265–275
Kochert G (1978) Carbohydrate determination by the phenol-sulfuric acid method. In: Hellebust JA, Craigie JS (eds) Handbook of phycology methods: physiological and biochemical methods. Cambridge University Press, London, pp 95–97
Marsh JB, Weinstein DB (1966) Simple charring method for determination of lipids. J Lipid Res 7:574–576
Tatsuoka MM (1970) Discriminant analysis: the study of group differences. Institute for Personality and Ability Testing, Champaign
Murthy HS, Yogeeshababu MC, Tejpal CS (2012) Preliminary evaluation of the potential of B. plicatilis for use as a live food for freshwater prawn larvae. Open Access Sci Rep. doi:10.4172/scientificreports.383
Sulkin SD (1975) The significance of diet in the growth and development of larvae of the blue crab, Callinectes sapidus Rathbun, under laboratory conditions. J Exp Mar Biol Ecol 20:119–135
Emmerson WD (1984) Predation and energetics of Penaeus indicus (Decapoda: penacidae). Larvae feeding on Brachionus plicatilis and Artemia nauplii. Aquaculture 38:201–209
Murthy HS (1998) Effect of enriched Artemia on metamorphosis and survival of larvae of the giant freshwater prawn, Macrobrachium rosenbergii. J Aquac Trop 13:215–222
Blair T, Castell J, Neil S, D’Abramo L, Cahu C, Harmon P, Ogunmoye K (2003) Evaluation of microdiets versus live feeds on growth, survival and fatty acid composition of larval haddock (Melanogrammus aeglefinus). Aquaculture 225:451–461
Baylon J, Failaman A (1997) Survival of mud crab Scylla oceanica from zoea to megalopa when fed the rotifer Brachionus sp and brine shrimp Artemia nauplii. UPV J Nat Sci 2:9–16
Godfred J, Ravi A, Kannupandi T (1997) Larval feed preference of the estuarine edible portunid crab Thalamita crenata (Laterille). Indian J Fish 44(1):69–74
Faleiro F, Narciso L (2009) Brachionus vs Artemia duel: optimizing first feeding of Upogebia pusilla (Decapoda: Thalassinidea) larvae. Aquaculture 295:205–208
Hammer HS, Bishop CD, Watts SA (2000) Activities of three digestive enzymes during development in the crayfish Procambarus clarkii (Decapoda). J Crustacean Biol 20(4):614–620
Kolkovski S (2001) Digestive enzymes in fish larvae and juveniles—implications and applications to formulated diets. Aquaculture 200:181–200
D’Abramo LR (2002) Challenges in developing successful formulated feed for culture of larval fish and crustaceans. In: Cruz-Suárez LE, Ricque-Marie D, Tapia-Salazar M, Gaxiola-Cortés MG, Simoes N (eds) Avances en Nutrición Acuícola VI. Memorias del VI Simposium Internacional de Nutrición Acuícola, Cancún, Quintana Roo, México
Agh N, Sorgeloos P (2005) Handbook of protocols and guidelines for culture and enrichment of live food for use in larviculture. Artemia & Aquatic Animals Research Center, Urmia University, Urmia
Decamp O, Moriarty DJW, Lavens P (2008) Probiotics for shrimp larviculture: review of field data from Asia and Latin America. Aquac Res 39:334–338
Rengpipat S, Phianphak W, Piyatiratitivorakul S, Menasveta P (1998) Effects of a probiotic bacterium on black tiger shrimp Penaeus monodon survival and growth. Aquaculture 167:301–313
Rengpipat S, Rukpratanporn S, Piyatiratitivorakul S, Menasaveta P (2000) Immunity enhancement on black tiger shrimp (Penaeus monodon) by a probiont bacterium (Bacillus S11). Aquaculture 191:271–288
Itami T, Asano M, Tokushige K, Kubono K, Nakagawa A, Takeno N, Nishimura H, Maeda M, Kondo M, Takahashi Y (1998) Enhancement of disease resistance of Kuruma shrimp, Penaeus japonicus, after oral administration of peptidoglycan derived from Bifidobacterium thermophilum. Aquaculture 164:277–288
McIntosh D, Samocha TM, Jones ER, Lawrence AL, McKee DA, Horowitz S, Horowitz A (2000) The effect of a commercial bacterial supplement on the high-density culturing of Litopenaeus vannamei with a low-protein diet in an outdoor tank system and no water exchange. Aquac Eng 21:215–227
Yu M, Li Zh, Lin H, Wen G, Ma Sh (2009) Effects of dietary medicinal herbs and Bacillus on survival, growth, body composition, and digestive enzyme activity of the white shrimp Litopenaeus vannamei. Aquac Int 17:377–384
Venkat HK, Narottam PS, Kamal KJ (2004) Effect of feeding Lactobacillus-based probiotics on the gut microflora, growth and survival of postlarvae of Macrobrachium rosenbergii (De Man). Aquac Res 35:501–507
Saad SA, Habashy MM, Sharshar MK (2009) Growth response of the freshwater prawn, Macrobrachium rosenbergii (De Man), to diets having different levels of Biogen. World Appl Sci J 6:550–556
Fernandez R, Sridhar M, Sridhar N (2011) Effect of lactic acid bacteria administered orally on growth performance of Penaeus indicus (H. Milne Edwards) juveniles. Res J Microb 6:466–479
Acknowledgments
The authors wish to thank people in Urmia Lake research center (Urmia, Iran) and the Gomishan shrimp production hatchery (Golestan-Iran) for their invaluable assistance. The authors wish to thank Mr. Pourfaraj (a Ph.D. Candidate at Simon Fraser University) for his assistance in English edition.
Conflict of interest
Hadi Jamali, Ahmad Imani, Daruosh Abdollahi, Reza Roozbehfar and Amin Isari declare that they have no conflict of interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jamali, H., Imani, A., Abdollahi, D. et al. Use of Probiotic Bacillus spp. in Rotifer (Brachionus plicatilis) and Artemia (Artemia urmiana) Enrichment: Effects on Growth and Survival of Pacific White Shrimp, Litopenaeus vannamei, Larvae. Probiotics & Antimicro. Prot. 7, 118–125 (2015). https://doi.org/10.1007/s12602-015-9189-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12602-015-9189-3