
Abstract
The influence of constant temperatures on biological parameters of German strains of Trichogramma aurosum Sugonjaev & Sorokina (Hymenoptera: Trichogrammatidae) was evaluated in the laboratory on eggs of Cydia pomonella L. (Lepidoptera: Tortricidae). Development time and longevity of all strains were decreased as temperature increased. Development time of the strains differed significantly only when exposed to 15°, 20°, and 25°C. Cumulative fertility and longevity differed significantly at 15° and 20°C. Realized fertility differed significantly at all constant temperatures. Emergence rates of all strains were less than 65% and were decreased even further as temperature increased. Female-biased sex ratio ranged from 65% to 100% at all constant temperatures. The low temperature threshold for T. aurosum was 10°C and the mean number of degree-days at 15°, 20°, 25° and 30°C was 175, 183, 173 and 185, respectively. The Bavarian strain tolerated high temperatures and had the highest parasitization capability, while the Hessian strain had the lowest parasitization at all temperatures. Fertility life table analysis revealed a major effect of temperature on the population growth parameters. Net reproductive rate was highest at intermediate constant temperatures in all strains, with the highest rate recorded for the Bavarian strains at all constant temperatures. Mean cohort generation time, and population doubling time decreased as temperature increased. The daily intrinsic rate of increase and finite rate of increase were positively correlated with temperature. The relevance of our results is discussed in the context of climatic adaptation, intraspecific variability and biological control.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae), is a major pest of apple, pear, peach, plum, cherry and walnut worldwide (Madsen and Morgan 1970). Infestation rates have been reported to reach 95% when no adequate control methods were applied (Ahmad and Abul-Hab 1977). Control of the codling moth has been tried in commercial orchards by pheromone trapping, trunk banding, sanitation, application of pesticides (Barnes 1957; Madsen and Morgan 1970), mating disruption (Bloem et al. 1999; Wang et al. 2001), and biological control (Unruh and Lacey 2001). At present, the management of the codling moth has shifted from application of chemical insecticides to integrated management that includes horticultural practices such as pruning and thinning. Although these practices are aimed primarily at producing a healthy, productive fruit tree, they may also improve control of the pest through habitat management (Ahmad and Abul-hab 1977). Emphasis is placed on the minimum use of pesticides that have a disruptive impact on the beneficial arthropods present in apple orchards (Blomefield et al. 1997; Unruh and Lacey 2001).
The gregarious egg parasitoids of the genus Trichogramma (Hymenoptera: Trichogrammatidae) are the most widely used natural enemies worldwide with several species being mass-produced and sold by a number of commercial companies (Smith 1996). Five species of Trichogramma have been reported to be potentially useful for control of codling moth. They include T. platneri Nagarkatti (Mills et al. 2000), T. minutum Riley and T. pretiosum Riley (Yu et al. 1984) in the Nearctic region, while T. dendrolimi Matsumura and T. cacoeciae (Marchal) (also erroneously identified as T. embryophagum Hartig) have been tested in the palearctic region (Hassan 1993). Mills et al. (2000) reported 60% reduction of damage in California walnut and apple orchards through releases of T. platneri, and Hassan (1993) suggested T. dendrolimi as a promising biocontrol agent.
Reduction in pest damage, however, does not necessarily mean that moth population deceased below the economic threshold at the end of the growing period. Hence, there is a necessity to seek additional candidate species. For instance, Pinto et al. (2002) reported 11 species of Trichogramma attacking tortricid eggs in apple and pear orchards in the USA. One of them, Trichogramma aurosum Sugonjaev and Sorokina, is a holarctic species that occurs naturally in Middle Europe (Samara 2005), the USSR (Livshits and Mitrofanov 1986; Lopatina 1983) and the USA (Pinto et al. 2002). It was collected in Germany for the first time in 2000. In host preference experiments it was shown that this species prefers eggs of the codling moth, C. pomonella, to other lepidopteran eggs. It may therefore be a potential candidate for the control of C. pomonella in apple orchards (Samara et al. 2008a, b). From 2001 to 2003, a wide collection of this species was carried out in the German Federal Republic from eggs of Nematus tibialis Newman (Hymenoptera: Tenthredinidae) on Robinia pseudoacacia (L.), in order to obtain strains that could be used for pre-introductory research.
Temperature plays a major role in the activity and metabolic processes of poikilothermic organisms such as insects (Suverkropp et al. 2001). Suitability of Trichogramma spp. for their use as bioagents is dependent on their ability to tolerate and adjust to adverse abiotic conditions (G.A. Pak, 1988, thesis, Agricultural Univ. of Wageningen). According to Pak and van Heiningen (1985), climatic adaptability is one of the criteria for the comparative selection of a strain through laboratory experiments. Many studies were conducted at constant temperatures to assess the biological characteristic of Trichogramma species (Maceda et al. 2003; H. E. Sakr, 2003, thesis, Univ. of Hohenheim; Schöller and Hassan 2001).
The impact of constant temperatures was investigated on the biology, parasitization potential and population growth parameters of different German strains of T. aurosum, in order to obtain suitable candidates that can possibly be used against C. pomonella in different parts of the country.
Materials and methods
Insect rearing
A stock culture of the codling moth C. pomonella was maintained in the laboratory according to Bathon et al. (1991). Strains of T. aurosum collected from the field (Table 1) were reared and maintained on eggs of the factitious host Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). The factitious host was reared following the procedure described in Cerutti et al. (1992). Strains were placed in culture tubes (70 × 20 mm) closed with a plastic lid, which had a small hole for aeration. The tubes were kept at ca 25°C, 65 ± 5% r.h. and a photoperiod of 18:6 h (L:D) in a climate cabinet during pupal development of the parasitoids. To feed the wasps, a droplet of honey was placed in the tube prior to or upon their emergence. Emerged parasitoids were provided with fresh host eggs on an ‘egg card’. Egg cards were prepared by sprinkling host eggs on a drop of gum arabic on a piece of paper index card (50 × 15 mm). Females used for the experiments were 24 h old, mated, fed on honey, and had no experience with hosts prior to the tests.
Effect of preimaginal development, emergence, and mortality
Temperature-dependent development of T. aurosum from egg to adult emergence was determined by allowing females to oviposit on 24-h-old C. pomonella eggs for 24 h. Females of each strain were placed singly (n = 20) in glass test tubes (70 × 20 mm) with 10–20 C. pomonella eggs to follow individual development and emergence. Development duration was calculated from the time when the test females were removed. Each group of test tubes was placed in a separate cabinet at constant temperature regimes of 15°, 20°, 25° and 30°C (all ±0.5°C) with a 16:8 h (L:D) photoperiod, and a relative humidity of 65 ± 5%. Development time from oviposition to adult emergence was determined by two visual controls for each parasitized egg every 12 h (07:00, 19:00). Degree-days and mean development threshold were determined according to Smith and Hubbes (1986).
Fertility and longevity
Mean cumulative female fertility and relative cumulative parasitized eggs per day were determined under the same conditions as given in the previous paragraph. Test females were obtained from egg cards with parasitized E. kuehniella eggs in a small glass vial (40 mm long, 15 mm diameter), 3–6 h before the experiments. A single mated female, maximum 16 h old, was placed on a circular filter paper that covered the bottom of a glass petri dish (70 mm diameter, 10 mm deep). One minute drop of undiluted honey was provided in the middle of the petri dish to feed the adult during this test. Fresh codling moth eggs (10–20 eggs per dish per day) were supplied daily until the natural death of the female wasps. Twenty replications were performed for each wasp strain and for each temperature treatment. Longevity (number of days) until death of the adults was also recorded. Hatched C. pomonella larvae from unparasitized eggs were removed daily from the vials to avoid parasitized eggs being consumed in the vials. Cumulative female fertility is defined as the total number of successfully parasitized eggs by a female over the full life span (as evidenced by black coloration of the eggs). The number of parasitized host eggs as well as the number of adults emerging from the host eggs and the number of unhatched eggs were counted. We refer to the number of live female progeny per female in each age interval as age-specific fertility (mx) (Southwood 1978). Cumulative realized fertility is the number of eggs successfully parasitized by a female wasp over the first 3 days of adult life (Kuhlmann and Mills 1999), since most Trichogramma sp. die within this period in nature if they are not able to find a food source. The parameters usually estimated from fertility life tables are the net reproductive rate (R0); the intrinsic rate of increase (rm), which is a measure of the growth rate of a population per female (Pak and Oatman 1982); the mean cohort generation time (Tc); the doubling time (Dt) (the time required for a population to double its numbers); and the finite rate of increase (λ). All were estimated according to Southwood (1978), Maia et al. (2000) and Nagarkatti and Nagaraja (1978).
Data analysis
Female longevity, cumulative fertility, realized fertility, and development time data were transformed to log10(x + 1), whereas the data of emergence rate and sex ratio were arcsine-transformed. The transformed data were then analyzed by ANOVA test using the General Linear Models (PROC GLM) procedure (SAS Institute 1996). The Student-Newman-Keuls (SNK) procedure was used to separate the means.
Results
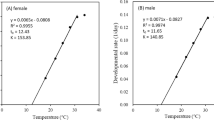
Significant differences in cumulative fertility among the various strains were detected at 15°C. Cumulative fertility was highest for Ta4 and Ta20, whereas Ta10 had the lowest values among all strains at 15°C (Table 2). The highest cumulative fertility was measured in strain Ta19 at 20° and 25°C. At the extreme temperature (30°C), cumulative fertility of Ta13 was significantly higher than that of the rest of the strains (Table 2), which did not differ significantly. Female longevity did not differ significantly at 25° or 15°C in all T. aurosum strains. At the highest temperature (30°C), females of all strains died after a maximum of 3 days. Females of Ta19 were able to live longest at all temperatures (Table 2). A significant difference was recorded at 30° and 20°C for Ta19 female longevity. Realized fertility for T. aurosum was calculated in response to temperature impact. Temperature had a significant impact on the female realized fertility for all strains studied. Ta20 had the highest realized fertility at 15°C, and Ta4 had the highest significant realized fertility at 20°C. At intermediate and high temperatures (25° and 30°C), both Ta19 and Ta13 had the highest significant differences, respectively (Table 2). Parasitism reached the highest values in the first 3 days at high temperature, whereas at the low temperature, parasitism ranged between 20% and 40% for the above mentioned strains. Mean development time was similar among all populations of Trichogramma sp. at 30°C, ranging from 8.6 to 9.2 days (Table 2). Significant differences were found among the strains at the remaining temperatures studied. No development was recorded for any strain at 35°C (preliminary experiments). The development rate from egg to adult of both female and male parasitoids increased with increasing temperature from 15° to 30°C (Fig. 1). The low temperature development threshold was 9.25°, 9.33°, 9.5°, 10.55° and 9.07°C, for Ta4, Ta10, Ta13, Ta19 and Ta20, respectively. The sum of degree-days (D-D, thermal constant) of Ta4, Ta10, Ta13, Ta19 and Ta20 was 180, 180, 185, 159 and 186 D-D, respectively. At 30°C no progeny hatched from the parasitized eggs of strain Ta19 even though black eggs were observed. This is an indication that the parasitoids reached the larval stage, but died during the prepupal stage. Therefore, the low temperature development threshold was higher and the sum of D-D was shorter than in the other strains. An inverse relationship was found between rearing temperature and development time (Fig. 1) and a direct relationship between rearing temperature and development rate. Sex ratio was significantly affected by temperatures. Interestingly, progeny of Ta10 was extremely female-biased (Table 2) compared with that of other strains, although the remaining strains had also a high sex ratio (>67%). Emergence rate of Ta10 was affected by low temperature (15°C). Ta4 had the highest emergence rate at all temperatures studied (Table 2).

Effect of constant temperatures on the development rate per day of different strains of Trichogramma aurosum
Fertility life table parameters differed significantly in response to the different temperature treatments. The net reproduction rate (R0) of Ta4, Ta10 and Ta19 varied from 3.22 to 8.12, 0.14 to 2.67, and 1.76 to 7.40 times, respectively, according to the temperature variation. The maximum increase in capacity was reached at 20°C (Table 3). The values for Ta13 and Ta20 varied from 1.56 to 2.77 and from 0.71 to 4.59 times, respectively; the maximum was reached at 30° and 25°C, respectively. The intrinsic rates of increase (rm) for the different strains is shown in Table 3; Ta4 and Ta13 had the highest rate at 30°C, whereas Ta19 and Ta20 had the highest rate at 25°C. Cohort generation time (Tc) for the studied strains differed significantly at variant temperatures. The finite capacity for increase (λ) of the parasitoids increased as temperature increased. Doubling time (Dt) decreased with the increase of temperature, the longest time required occurring at 15°C and the shortest time at 30°C.
Discussion
Temperature is one of the most important abiotic factors affecting the development rate, cumulative fertility, longevity, sex ratio and emergence rate of Trichogramma spp. Tolerance of immature stages of Trichogramma species/strains to high or low temperature extremes has been the subject of several studies (G. A. Pak, thesis, 1988; Nagarkatti and Nagaraja 1978; Smith and Hubbes 1986). However, most of the published work was conducted only at constant temperatures, which are useful for laboratory rearing and commercial mass production. No significant differences among strains were observed in longevity or development time at some temperatures tested, whereas the duration of development and longevity decreased as temperature increased. This reduction could be due to the long rearing of the wasps under constant temperature (the wasps used in these experiments had been reared for at least 40 generations under laboratory conditions). It is not well known whether the long laboratory rearing affects the wasps’ activity and vigor. According to Nagarkatti and Nagaraja (1978), female fertility of T. confusum wasps reared for a long time under laboratory conditions was significantly lower than that of wild females. However, female longevity is affected by many factors, such as temperature (Pak and Oatman 1982), humidity (Stinner et al. 1974), host size (Stinner et al. 1974), and food (Yu et al. 1984). Almatni (2003, Ph.D. thesis, Damascus Univ., Syria) found that the high temperature could have caused sterilization of European strains of T. cacoeciae, because they stopped laying eggs but lived for a few more days without laying new eggs. Also a reduction in female longevity of T. platneri was recorded from 53 days at 10°C and 3 days at 35°C (McDougall and Mills 1997).
The highest cumulative fertility for the strains studied was at intermediate temperatures (20° and 25°C). These results agree with those of G. A. Pak (thesis, 1988), who found that the number of parasitized hosts increased with increasing temperature to a maximum at 20–25°C and declined at 30°C. Some strains examined in this study showed better tolerance to high temperatures and others were able to tolerate low temperatures. The strains Ta4, Ta19 and Ta20 were able to live up to 6–11 days longer than the rest of the strains at 15°C, and Ta13 was able to live up to 1–2 days longer than the rest of the strains at 30°C. Relative cumulative parasitized eggs per day was temperature-dependent, females reaching 100% parasitization at the high temperature regimes in a short time after hatching. On the contrary, females held under low temperatures reached 100% parasitization after a longer period of time. The short period of parasitism can be considered as a specific survival strategy, because a faster oviposition at higher temperatures will allow this pro-ovogenic parasitoid to lay most of its available eggs during a short lifetime period (Garcia and Tavares 1994). Garcia and Tavares (1994) found significant differences between all temperatures for T. cordubensis longevity, which increased with the decrease of temperature, results that are similar to ours. However, it was noticed that the decrease in temperature increased the pre-oviposition period for all the studied strains. At 15°C the average pre-oviposition period was 3, 5, 2, 1.5 and 2 days for Ta4, Ta10, Ta13, Ta19 and Ta20, respectively. There was no measurable pre-oviposition period at the higher temperatures. According to Al-Ahmed and Kheir (2003), temperature is considered an important factor affecting the duration of the pre-oviposition period.
Realized cumulative fertility was calculated according to Mills and Kuhlmann (2000), and found to be the highest at the high temperature regime. This parameter was reduced at the lower temperature regime. The total cumulative fertility values did not differ from the values of the realized cumulative fertility at high constant temperatures, but both values were found to differ when T. aurosum strains were reared at 15°C. Similar results were reported for T. minutum (Smith and Hubbes 1986) and Trichogramma spp. (Pak and van Heiningen 1985).
According to Jervis and Copland (1996), there is an optimal range of temperature for insect development, beyond which they would be unable to continue oogenesis and laying eggs or unable to function appropriately for a long period of time. This could be due to the increase of respiration rate, i.e., the insects would be unable to produce fertile eggs due to the high consumption of energy (Mills and Kuhlmann 2000). Data recorded for sex ratio from the present experiments agree with some data from the literature but disagree with other data. Pintureau and Bolland (2001) found that the percentage of males in the offspring of thelytokous females increased faster according to temperature in T. cordubensis than in T. pretiosum; the sex ratio was higher at low and intermediate temperatures. These results agree with those of Bowen and Stern (1966).
In general, sex ratio was not affected with an increase of temperature; female percentage ranged from 65% to 100%. Our data agree with Haile et al. (2002), who found that sex ratio was biased to female production at all temperatures. It seems that rearing the wasps at lower temperatures has a significant effect on the biological characteristics. Crozier (1977) suggested that lower temperatures could promote fusion of nuclei and increase the proportion of diploid offspring, which could explain the high sex ratio at 15°C in all strains studied.
Development time needed by the T. aurosum strains was shorter as temperature increased; this was in agreement with Consoli and Parra (1995), who assumed that this might be due to a more appropriate metabolic process of the immature stages. The optimal temperature for development and survival was 25°, and 35°C was assumed to be the upper vital threshold (McDougall and Mills 1997; G. A. Pak, thesis, 1988). Total development time was four times faster at 30° than at 15°C. However, the total mortality was greater at the higher temperature; these results are similar to the findings of Hawkins and Smith (1986). Mean development time at constant temperature ranged between 32.2 and 8.8 days at temperatures between 15° and 30°C. Although development occurred at 10.8°C (indicated by the parasitized eggs turning black, i.e., development takes place up to the third larval stage), no emergence was observed during 6 months of incubation (unpublished data). For all the temperatures tested, a linear regression model was calculated for egg to adult development of all Trichogramma strains. As temperature increased, the duration of development decreased. Mean development time of female T. turkestanica on E. kuehniella was 32.9, 18.2, 9.1 and 7.0 days at 15°, 20°, 25° and 30°C, respectively (Hansen 2000; Pintureau and Bolland 2001).
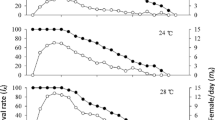
Survivorship curves according to Southwood (1978) obtained from our results at different constant temperatures differed remarkably for the strains studied. At low constant temperature (15°C) the mortality acts most heavily on the old females. All strains at 20°C had a constant mortality per unit time; whereas Ta10, Ta19 and Ta20 had a constant rate of mortality, Ta4 had the highest mortality rate of the young individuals. At 25°C, all strains had a logarithmic survivor rate where the mortality rate is constant. At 30°C, all strains had the highest mortality rate during the young stages (Fig. 2). The Ta13 strain showed a better adaptation to all temperatures studied, althoughTa4 and Ta20 also showed promising results in response to rearing temperature. The relationship between the temperature of the geographical origins of the strains and their climatic adaptability was evaluated. We found that Ta13, which was collected from southeast Germany—where the average temperature from June to September is 15–19°C—was able to tolerate the high constant temperature (30°C). Ta4, Ta19 and Ta20 were able to tolerate intermediate and low temperatures; in the above mentioned season the average temperature range in the original locations was 13–16°, 14–17° and 14–18°C, respectively.

Survivorship in five strains of Trichogramma aurosum at four constant temperatures
Emergence rate reached the highest values at intermediate temperatures in all strains studied. For T. annulata and T. pretiosum, it was found to be more than 89% (Maceda et al. 2003). The reduction in the emergence rate and the long time required for the immature stages to develop from egg to adult were recorded at the low temperature 15°C for all strains studied. This can be due to the high mortality in the immature stages. Smith and Hubbes (1986) noticed that the emergence rate was reduced when the parasitized eggs were reared for 24 days at 15°C. Fertility life table studies provide a powerful technique for evaluation of population dynamics because they provide a detailed description of age-specific mortality of individuals in the population. Pratissoli and Parra (2000) found that the net reproduction rate varied according to the temperature variation for T. pretiosum. It was the highest at 20°C for all the T. aurosum strains studied. The net reproductive rate recorded for T. mwanzai at the constant temperatures 20°, 25° and 30°C was 7.3, 35.9, and 31.9 (Lu 1992), and for T. cacoeciae at the same temperatures it was 48.69, 45.83 and 24.02, respectively (Uzun and Akten 1992). The finite increase rate was proportionally related to temperature. For T. pretiosum the relation between λ and temperature increase occurred for the range from 18° to 30°C (Pratissoli and Parra 2000), and for Trichogrammatoidea sp. it was the highest at 27°C (Baitha and Ram 1998) and in our results it was at 30°C. The mean cohort generation time (Tc) values show a decreasing trend from 18° to 30°C (Baitha and Ram 1998). This agrees with our findings, where the generation time values decreased as the temperature increased. The differences between the strains were very obvious, some strains showed good adaptability to high temperature (Ta13), whereas others showed good adaptability to intermediate temperatures (Ta4, Ta10 and Ta19), and some showed adaptability to a low temperature (Ta20). Cabello and Vargas (1988) related the reduction of the net reproductive rate at high temperatures to the production of both males and females at these temperatures. This can explain our results, where the R0 was reduced at 30°C. Accordingly, values of daily intrinsic rate of increase (rm) and finite rate of increase (exp. rm) were affected. Development of each insect stage is dependent on temperatures; insect activity such as locomotion and searching behavior is dependent on the temperature.
This study can provide us with information to select one or several strains as a suitable candidate for biological control of the codling moth according to their climatic adaptability. It is possible to select either those strains that showed high parasitization rate, higher longevity and higher tolerance to high temperatures, or those strains that showed high parasitization activity at low temperature conditions. However, further studies of host location and searching, parasitization behavior and female dispersal are also required before field release experiments are conducted.
References
Ahmad, F. R., & Abul-hab, J. K. (1977). Biological studies of the codling moth Cydia pomonella L. (Lepidoptera: Olethreutidae) in the Middle of Iraq. Iraqi Journal of Agricultural Science, 12, 210–225.
Al-Ahmed, A. M., & Kheir, S. M. (2003). Life cycle and survival of Hyalomma dromedarii (Acari: Ixodidae) under laboratory conditions. Agricultural and Marine Sciences, 8, 11–14.
Baitha, A., & Ram, A. (1998). Life table studies on Trichogrammatoidea sp. nr. armigera Nagaraja an egg parasitoid of Helicoverpa armigera. Indian Journal of Entomology, 60, 250–256.
Barnes, M. (1957). Deciduous fruit insects and their control. Journal of Economic Entomology, 50, 226–228.
Bathon, H., Singh, P., & Clare, G. K. (1991). Rearing methods. In L. P. S. van der Geest (Ed.), Tortricid pests: Their biology, natural enemies and control (pp. 283–309). Amsterdam, the Netherlands: Elsevier Science Publishers.
Bloem, S., Bloem, K. A., Carpenter, J. E., & Calkins, C. O. (1999). Inherited sterility in codling moth (Lepidoptera: Tortricidae): effect of substerilizing doses of radiation on insect fecundity, fertility, and control. Annals of the Entomological Society of America, 92, 222–229.
Blomefield, T. L., Pringle, K. L., & Sadie, A. (1997). Field observations on oviposition of codling moth, Cydia pomonella (Linnaeus) (Lepidoptera: Olethreutidae), in an unsprayed apple orchard in South Africa. African Entomology, 5, 319–336.
Bowen, W. R., & Stern, V. M. (1966). Effects of temperature on the production of males and sexual mosaics in a uniparental race of Trichogramma semifumatum (Hym.: Trichogrammatidae). Annals of the Entomological Society of America, 73, 671–673.
Cabello, T., & Vargas, P. (1988). Resistance to high temperatures in the developmental stages of Trichogramma cordubensis Vargas and Cabello and T. pintoi Voegelé (Hym.: Trichogrammatidae). Boletin de Sanidad Vegetal, Plagas, 15, 263–266.
Cerutti, F., Bigler, F., Eden, G., & Bosshart, S. (1992). Optimal larval density and quality control aspects in mass rearing of the Mediterranean flour moth Ephestia kuehniella Zell. (Lepidoptera; Phycitidae). Journal of Applied Entomology, 114, 353–361.
Consoli, F. L., & Parra, J. R. (1995). Effects of constant and alternating temperatures on Trichogramma galloi Zucchi (Hymenoptera. Trichogrammatidae) biology. I. Development and thermal requirements. Journal of Applied Entomology, 119, 415–418.
Crozier, R. H. (1977). Evolutionary genetics of the Hymenoptera. Annual Review of Entomology, 22, 263–288.
Garcia, P., & Tavares, J. (1994). Parasitic capacity, longevity and development of Trichogramma cordubensis (Hymenoptera: Trichogrammatidae) at three temperature regimes. Les Colloques de l’INRA, 73, 4–7.
Haile, A. T., Hassan, S. A., Sithanantham, S., Ogol, C. K., & Baumgärtner, J. (2002). Comparative life table analysis of Trichogramma bournieri Pintureau and Babault and Trichogramma sp. nr. mwanzai Schulten and Feijen (Hym., Trichogrammatidae) from Kenya. Journal of Applied Entomology, 12, 287–292.
Hansen, L. S. (2000). Development time and activity threshold of Trichogramma turkestanica [Trichogramma evanescens] on Ephestia kuehniella in relation to temperature. Entomologia Experimentalis et Applicata, 96, 185–188.
Hassan, S. A. (1993). The mass rearing and utilization of Trichogramma to control lepidopterous pests: achievements and outlook. Pesticide Science, 37, 387–391.
Hawkins, B. A., & Smith, J. W. (1986). Rhaconotus roslinensis (Hymenoptera: Braconidae), a candidate for biological control of stalkboring sugarcane pests (Lepidoptera: Pyralidae): development, life tables and intraspecific competition. Annals of the Entomological Society of America, 79, 905–911.
Jervis, M. A., & Copland, M. J. W. (1996). In M. A. Jervis & N. A. Kidd (Eds.), Insect natural enemies: Practical approaches to their study and evaluation (pp. 63–161). London, UK: Chapman & Hall.
Kuhlmann, U., & Mills, N. J. (1999). Comparative analysis of the reproductive attributes of three commercially produced Trichogramma species (Hym.: Trichogrammatidae). Biocontrol Science and Technology, 9, 335–346.
Livshits, I. Z., & Mitrofanov, V. I. (1986). Hymenopterans—parasites of eggs of fruit pests. Zashchita Rasteniij, 9, 46–47.
Lopatina, V. V. (1983). Ecology of Trichogramma aurosum Sug. et Sor. in the Chita Region. Bjulleten’ Vsesojuznogo Nauchno Issledovatel’skogo Instituta Zashchity Rasteniij, 55, 9–12.
Lu, Q. G. (1992). Fertility and life table study on Trichogramma (Hym.: Trichogrammatidae) sp. nr. mwanzai from Kenya. Chinese Journal of Biological Control, 8, 16–18.
Maceda, A., Hohmann, C. L., & dos Santos, H. R. (2003). Temperature effects on Trichogramma pretiosum Riley and Trichogrammatoidea annulata De Santis. Brazilian Archives of Biology and Technology, 46, 27–32.
Madsen, H. F., & Morgan, C. V. (1970). Pome fruit pests and their control. Annual Review of Entomology, 15, 295–320.
Maia, A. H., Luiz, A. J., & Campanhola, C. (2000). Statistical inference on associated fertility life table parameters using Jackknife technique: computational aspects. Journal of Economic Entomology, 93, 511–518.
McDougall, S. J., & Mills, J. (1997). The influence of hosts, temperature and food sources on the longevity of Trichogramma platneri. Entomologia Experimentalis et Applicata, 83, 195–203.
Mills, N. J., & Kuhlmann, U. (2000). The relationship between egg load and fecundity among Trichogramma parasitoids. Ecological Entomology, 25, 315–324.
Mills, N., Pickel, C., Mansfield, S., McDougall, S., Buchner, R., Caprile, J., et al. (2000). Mass releases of Trichogramma wasps can reduce damage from codling moth. California Agriculture, 54, 22–25.
Nagarkatti, S., & Nagaraja, H. L. (1978). Experimental comparison of laboratory reared vs. wild type Trichogramma confusum (Hym., Trichogrammatidae). 1. Fertility, cumulative fertility and longevity. Entomophaga, 23, 129–136.
Pak, G., & Oatman, E. R. (1982). Comparative life table behaviour and competition studies of Trichogramma brevicapillum and Trichogramma pretiosum. Entomologia Experimentalis et Applicata, 32, 68–79.
Pak, G., & van Heiningen, T. (1985). Behavioural variations among strains of Trichogramma spp.; Adaptability to field temperature conditions. Entomologia Experimentalis et Applicata, 38, 3–13.
Pinto, J. D., Koopmanschap, A. B., Platner, G. R., & Stouthamer, R. (2002). The North American Trichogramma (Hymenoptera: Trichogrammatidae) parasitizing certain Tortricidae (Lepidoptera) on apple and pear, with ITS2 DNA characterizations and description of a new species. Biological Control, 23, 134–142.
Pintureau, B., & Bolland, P. (2001). Response to selection for a high or a low sex ratio in Wolbachia-infected lines of Trichogramma cordubensis subjected to high temperature. Vestnik Zoologii, 35, 83–86.
Pratissoli, D., & Parra, J. R. (2000). Fertility life table of Trichogramma pretiosum (Hym., Trichogrammatidae) in eggs of Tuta absoluta and Phthorimaea operculella (Lep., Gelechiidae) at different temperatures. Journal of Applied Entomology, 124, 339–342.
Sakr, H. E. (2003). Use of egg parasitoids of the genus Trichogramma to control the codling moth Cydia pomonella (Lepidoptera: Tortricidae); selection of effective parasitoid species. Ph.D. thesis, Univ. of Hohenheim. Stuttgart, Germany: Verlag Ulrich E. Grauer.
Samara, R. (2005). Biology, behaviour and genetic diversity of Trichogramma aurosum Sugonjaev and Sorokina (Hymenoptera: Trichogrammatidae). Ph.D. thesis, Univ. of Hohenheim. Stuttgart, Germany: Verlag Ulrich E. Grauer.
Samara, R., Monje, J. C., Reineke, A., & Zebitz, C. P. W. (2008). Genetic divergence of Trichogramma aurosum Sugonjaev and Sorokina (Hymenoptera: Trichogrammatidae) individuals based on ITS2 and AFLP analysis. Journal of Applied Entomology, 132, 230–238.
Samara, R. Y., Monje, J. C., & Zebitz, C. P. W. (2008). Comparison of different European strains of Trichogramma aurosum (Hymenoptera: Trichogrammatidae) using fertility life tables. Biocontrol Science and Technology, 18, 75–86.
SAS Institute (1996). SAS users guide: Statistics. Cary, NC, USA: SAS Institute.
Schöller, M., & Hassan, S. A. (2001). Comparative biology and life tables of Trichogramma evanescens and T. cacoeciae with Ephestia elutella as host at four constant temperatures. Entomologia Experimentalis et Applicata, 98, 35–40.
Smith, S. M. (1996). Biological control with Trichogramma: advances, successes and potential of their use. Annual Review of Entomology, 41, 375–406.
Smith, S. M., & Hubbes, M. (1986). Isoenzyme patterns and biology of Trichogramma minutum as influenced by rearing temperature and host. Entomologia Experimentalis et Applicata, 42, 249–258.
Southwood, T. R. (1978). Ecological methods with particular reference to the study of insect population (2nd ed., pp. 1–6; 356–387). London, UK: Chapman and Hall.
Stinner, R. E., Ridgway, R. L., & Morrison, R. K. (1974). Longevity, fecundity, and searching ability of Trichogramma pretiosum reared by three methods. Environmental Entomology, 3, 558–560.
Suverkropp, B. P., Bigler, F., & van Lenteren, J. C. (2001). Temperature influences walking speed and walking activity of Trichogramma brassicae (Hym., Trichogrammatidae). Journal of Applied Entomology, 125, 303–307.
Unruh, T. R., & Lacey, L. A. (2001). Control of codling moth, Cydia pomonella (Lepidoptera: Tortricidae), with Steinernema carpocapsae: effects of supplemental wetting and pupation site on infection rate. Biological Control, 20, 48–56.
Uzun, S., & Akten, T. (1992). The effect of different temperatures on biology of Trichogramma cacoeciae March. Proceedings of the Second Turkish National Congress of Entomology (Adana, Turkey, pp. 403–410).
Wang, S., Ikedialaa, J. N., Tang, J., Hansen, J. D., Mitcham, E., Mao, R., et al. (2001). Radio frequency treatments to control codling moth in in-shell walnuts. Postharvest Biology and Technology, 22, 29–38.
Yu, D. S., Hagley, E. A., & Laing, J. E. (1984). Biology of Trichogramma minutum Riley collected from apples in southern Ontario. Environmental Entomology, 13, 1324–1329.
Acknowledgments
The present research was supported by the German Academic Exchange Service (DAAD, A/00/19004). Our thanks are extended to all who helped in the field collection and to R. Siegel for maintaining the lab culture of E. kuehniella.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Samara, R., Monje, J.C., Zebitz, C.P.W. et al. Comparative biology and life tables of Trichogramma aurosum on Cydia pomonella at constant temperatures. Phytoparasitica 39, 109–119 (2011). https://doi.org/10.1007/s12600-010-0142-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12600-010-0142-4