
Abstract
Red snappers were examined for reproductive biology and age-0 habitat preferences. Spawning in red snappers occurred throughout the year in northern Australia and eastern Indonesia; at least 10–30% of females and 40–80% of males were in ripe or spawning condition in most months. Northern Australian populations showed a spawning peak from July to December (L. erythropterus) and September to March (L. malabaricus). Eastern Indonesian L. malabaricus had a less defined pattern with two peaks: January–March and October. Size at first maturity was 240 mm for males and 250–300 mm for females. L 50 estimates were similar between species in northern Australia: 270–280 mm (males) and 350–370 mm (females). Maximum batch fecundity was 676,100 oocytes for L. erythropterus and 997,000 oocytes for L. malabaricus. Higher mean abundances of age-0 L. erythropterus were found in silty and coarse sand/rubble estuarine habitats of northern Australia (456 ± 119 fish/km2) compared with sandy coastal habitats (5 ± 3 fish/km2). Most age-0 snapper caught at Sape (eastern Indonesia) were L. malabaricus (91%) with mean abundances of 312 ± 14 fish/km2. The similarities in the reproductive characteristics of red snappers suggest that successful management approaches adopted in northern Australia should be considered in eastern Indonesia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Red snappers (genus Lutjanus) support important coastal fisheries in many tropical oceans. In the Indo-Pacific region, increased fishing pressure on populations of the red snappers Lutjanus erythropterus and L. malabaricus has meant a need for improvements in management. These two species make up a significant part of the commercial finfish and recreational catches in northern Australia [1, 2]. Currently, red snapper resources are commercially exploited by drop-lines, traps and finfish trawling in the Northern Territory and Western Australian waters, and finfish trawling in Queensland waters. In 2002, the combined catch of red snappers from the Northern Territory and Queensland finfish trawl fisheries was 800 tonnes, with another 400 tonnes taken from the Northern Territory trap fishery [2]. Red snappers are also extensively targeted throughout eastern Indonesia from small-scale artisanal fisheries to semi-industrial-scale fishing vessels using bottom long-lines, drop-lines and fish traps, to large industrial fish trawlers [2]. As many of these red snappers are either landed in local fish market sites across eastern Indonesia or transferred at sea to export carrier vessels, it is difficult to obtain accurate estimates of catches of these species. The annual catch of snappers in 1997 from Indonesian waters was estimated at 52,800 tonnes, with about 20% of this coming from the Timor and Arafura Seas, close to the international border between Australia and Indonesia [2]. While the Australian fisheries are strictly managed with both input and output controls, the Indonesian fisheries are largely unregulated with some commercial fishing licensing controls [2].
Red snappers have an extensive distribution across northern Australia to eastern Indonesia. Recent genetic studies using allozyme electrophoresis and mitochondrial DNA sequencing suggested that there are two broad fishery stocks of L. erythropterus with a genetic boundary between Kupang and the Australian Timor Sea and seven distinct stocks for L. malabaricus throughout the regions [3]. Further, their data did not distinguish separate genetic stocks among the four sampling locations across northern Australia nor among the central Indonesian sites—Bali to Sape—for both L. erythropterus and L. malabaricus [3]. While the red snapper stocks are genetically separate between northern Australia and eastern Indonesia, information on the life histories of each population, such as reproduction and recruitment, are important for future assessment and management of these fished stocks given their close proximity to each other.
There is some published data available on the reproductive characteristics of tropical lutjanids. Most species are highly fecund, serial spawners with a protracted spawning season [4–8]. Sexual maturity generally occurs at between 40% and 50% of maximum fish length, males mature at a slightly smaller size than females and larger fish have greater relative reproductive output [5]. There have also been a number of studies on seasonality and habitat preferences for age-0 red snapper (Lutjanus campechanus) in the Gulf of Mexico [9–13]. These studies reported that young snapper frequently occurred corresponding to the peak spawning season of adults and showed habitat preferences for low-relief fine sand and mud sediments. However, there has been little information published on the reproductive or recruitment patterns of the two commercially important red snappers; L. erythropterus and L. malabaricus, in either northern Australia or eastern Indonesia [14].
The aims of this study were to (1) examine the reproductive characteristics of the two Indo-Pacific red snapper species L. erythropterus and L. malabaricus (2) determine the extent of variation between populations from northern Australia and eastern Indonesia and (3) identify habitats of age-0 red snappers within the two regions.
Materials and methods
Sample collection
Biological data for this study were collected during a collaborative research project between Australia and Indonesia on shared snapper stocks [2]. The sampling methods used to collect fish were therefore constrained to the current fishing gears used at each sampling site in each country. In northern Australia, red snappers were obtained from commercial and research vessels using fish or prawn trawl nets between July 1998 and June 2002. In eastern Indonesia, red snapper specimens were collected by fisheries research staff and local fishers at fish landing sites across eastern Indonesia between February 1999 and December 2001. These fish were caught using commercial fish trawls, bottom drop-lines or fish traps.
Three research surveys using four-fathom otter trawls were also undertaken in the shallow waters (<10 m) of Darwin Harbour, Weipa and Gove (northern Australia) in May 2000, May 2001 and June 2002, respectively, to collect age-0 red snappers. These fish were <121 mm in standard length and were considered to be less than 1 year old (Fry G, unpublished data, 2005). Catch data from commercial and research prawn trawling within the Gulf of Carpentaria were also included to increase the monthly coverage of size structure data in northern Australia. Age-0 red snappers were also collected from monthly surveys using purse-seine nets at Sape (eastern Indonesia) from March 2000 to March 2002. For the abundance and size structure comparisons between these habitats, the otter trawl and seine net catches were standardised to mean numbers (±1 SE) of fish/km2. As fish were collected from a number of sites within northern Australia and eastern Indonesia and using a range of gears, mean fish lengths were compared using a general linear model in SAS (V9.1) to identify potential gear selectivity that could bias regional comparisons of their life-history traits.
Fish were either dissected for biological data in the field or frozen and returned to CSIRO Marine Laboratories (Australia) or Gondol Fish Laboratory (eastern Indonesia) for processing. Each fish was identified, measured for standard length (±1 mm) and weighed (±1 g). All age-0 red snappers collected from northern Australia and eastern Indonesia were frozen and returned to CSIRO Marine Laboratories (Australia) for processing. Snappers that were too small to be accurately identified to species were identified using starch gel electrophoresis [3].
Reproductive characteristics
Gonads were removed from the visceral cavity, weighed (±0.001 g) and placed in a fixing solution of 5% formaldehyde. The reproductive patterns of the red snappers were assessed by two methods: gonadosomatic index (GSI) and histology. GSI was calculated for each fish for both sexes to determine the spawning seasons using the equation:
For histological examination of gonads, a subsample of gonadal tissue was removed from each fish. These gonad samples were weighed (±0.001 g), placed in tissue cassettes, dehydrated and impregnated with wax. Histological sections were cut at 8 μm from each block using a tissue microtome, mounted on glass microscope slides and stained with Harris’s haematoxylin and eosin counterstain. Each histological section was scored by estimating the percentage that each of the gonad maturity stages occupied within the total area of the section. Female gonads were classified into maturity stages: I, oogonia; II, previtellogenic oocytes; III, yolk precursor; IV, nonstaining (primary) yolk; V, red-staining (secondary) yolk; VI, spent; and for males: stages I, stem spermatogonia (primary germ cells); II, spermatogonia; III, primary spermatocytes; VI, secondary spermatocytes; V, spermatids; VI, spermatozoa; VII, spent [15, 16]. As no observable difference in scoring was detected between replicate blocks from the same fish, only one subsample was taken from the mid-position on a randomly selected gonad lobe for the remaining red snappers samples.
Size at maturity
Size at first sexual maturity (L MAT) was determined from the minimum standard length of fish with developed vitellogenic eggs (maturity stages IV or V) for females and spermatids (maturity stages V or VI) for males. Gonads were classed as ripe when the majority of the gonad was in maturity stages IV and V for females and stages V and VI for males. Fish were in spawning condition when the greatest proportion of their gonad was in stage V (females) and stage VI (males).
To estimate the size of fish in the population where 50% of fish in a length class were mature (L 50), logistic curves with 95% confidence intervals were applied to the proportion mature-at-length data. Fish were grouped into 20-mm standard length classes to increase sample sizes. The logistic function [17] was defined as;
where P L is the proportion of mature fish in standard length class L, and K is the curvature of parameters. Models for each population were fitted by nonlinear regression (Proc NLIN) in SAS (V9.1). Parameters and their standard errors were estimated by least squares. We compared the slopes of the curves (K) and the estimated length at 50% maturity (L 50) with t-tests.
Batch fecundity
Batch fecundity (F B) of mature, ripe females was estimated from ovary subsamples containing hydrated oocytes (stage V). A subsample of gonad tissue was weighed, oocytes were teased apart from connective tissue and the number of hydrated ripe oocytes counted under a stereomicroscope. Batch fecundity was calculated by;
T-tests were applied to the mean batch estimates to compare between the two red snapper species and among regions. Relative fecundity: the mean number of eggs per gram of body weight of fish was also calculated and compared among species and regions using a t-test. The mean maximum oocyte size was determined by measuring the diameters from 20 randomly selected maturity stage V oocytes from each mature, spawning female fish.
Results
Sample collection
A total of 1,826 L. erythropterus (33–468 mm) and 1,736 L. malabaricus (83–645 mm) from four sites within northern Australia (Timor Sea, Arafura Sea, west Gulf and east Gulf) and 344 L. erythropterus (27–580 mm) and 2,198 L. malabaricus (22–740 mm) from nine sites within eastern Indonesia (Bali, Tanjung Luar, Sape, Kupang, Timor Sea, Ambon, Tual, Arafura Sea and Merauke) were collected for reproductive and recruitment analysis (Table 1; Fig. 1). There were differences in mean fish length among sites within northern Australia and eastern Indonesia for both L. erythropterus (F 9,1916 = 16.5; P < 0.0001) and L. malabaricus (F 10,2955 = 34.7; P < 0.0001). This was at least partly an effect of the types of sampling gear used at each site: L. erythropterus (F 1,1916 = 78.3; P < 0.0001) and L. malabaricus (F 1,2955 = 4.1; P < 0.05). However, the mean sizes of both L. erythropterus or L. malabaricus between Australia and Indonesia were similar (F 1,1916 = 3.8; P > 0.05 and F 1,2955 = 1.5; P > 0.21, respectively). Due to these biases in fishing gears across sites, low sample numbers collected from some sites and the genetic stocks for each species reported to be broadly separated along these boundaries [3], comparisons on their reproductive traits were made across the two regions and not between sites within each region. For the age-0 snapper surveys, the otter trawl and seine nets used also caught similar-sized red snappers (L. erythropterus) from northern Australia and eastern Indonesia (F 2,237 = 2.5; P > 0.08).

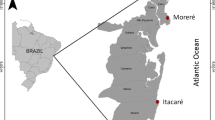
Map showing the sites across northern Australia and eastern Indonesia sampled for the red snappers, L. erythropterus and L. malabaricus
Reproductive characteristics
Gonads from 1,483 L. erythropterus and 2,395 L. malabaricus were sectioned for histological examination to determine gonad maturity stage and size at maturity. The sex ratios of both red snapper species differed from 1:1, being skewed towards females at most sites in northern Australia and eastern Indonesia (χ 226 = 59.6, P < 0.001) (Table 1). The maximum gonadosomatic index (GSI) for female red snappers was approximately 4–6% of body weight and was similar between the two species and populations (Fig. 2). However, male L. erythropterus from both northern Australia and eastern Indonesia showed higher GSI (up to 4%) than L. malabaricus males (0.5–2% of body weight). The mean monthly GSI for the red snappers suggested that most spawning takes place between the months of July to December for L. erythropterus and September to March for L. malabaricus in northern Australia (Fig. 3). The pattern for the eastern Indonesian red snappers was less defined. There were multiple peaks in GSI through the year for male and female L. erythropterus, however this may be caused by low sample numbers in these months. There were two broad peaks in spawning activity for L. malabaricus females; in January to March and in October, and males in February to June and August to December (Fig. 3).

Plot of gonadosomatic index and standard length for the red snappers, L. erythropterus and L. malabaricus. Solid and dashed lines indicate estimated length at first maturity (L MAT) for each sex

Plot of mean gonadosomatic index and month for mature red snappers, L. erythropterus and L. malabaricus for each sex. Sample numbers are shown for each month and standard error bars indicate ±1 SE
The histological examination of gonads supported the reproductive pattern shown from the GSI data; months with highest GSI values recorded corresponded to months where most gonads were staged as ripe and spawning condition. In northern Australia, the majority of oocytes in female red snapper gonads were classed as stage IV and V in the months of July to December for L. erythropterus and September to April for L. malabaricus (Fig. 4). Furthermore, it was in these months that female gonads had the highest proportion of spent gonad tissue (stage VI). The male populations in northern Australia had gonads with mature spermatozoa (stages V and VI) throughout most of the year. In eastern Indonesia, female L. malabaricus had the highest proportions of stage IV and V oocytes in their gonads between January to March and October (Fig. 4). There was little seasonal spawning pattern evident from gonad histology in the eastern Indonesian L. erythropterus populations, which may be due to lower sample numbers. Histological examination of female red snapper gonads from both northern Australia and eastern Indonesia did show there was a proportion of fish that were in ripe or spawning condition: at least 10–30% of females and 40–80% of males, in most months of the year (Fig. 4). There were also female red snappers in northern Australia and eastern Indonesia that had some stage VI (spent) tissue in their gonads in nearly all months sampled, indicating that spawning had just occurred.

Histograms of percentage of gonad maturity stages for female and male red snappers, L. erythropterus and L. malabaricus from northern Australia and eastern Indonesia. Sample numbers are shown above bars
Size at maturity
Sexual maturity occurs at about 32–50% of maximum body length for L. malabaricus and 51–62% for L. erythropterus. There were distinct differences in length at first maturity between the sexes, with male red snappers of both species first reaching sexual maturity at around 240 mm SL whereas females began maturing between 250 and 300 mm SL (Table 2). Although the smallest male L. erythropterus with developing gonads in eastern Indonesia was 309 mm SL, this is likely to be an effect of low sample numbers in the smaller size classes. The actual length at first maturity is probably comparable with males of this species from northern Australia.
There were no differences in the slope parameter (K) of the logistic functions for the L 50 estimates between either the sexes or species of L. erythropterus and L. malabaricus in northern Australia (t-test; all P > 0.05) (Fig. 5). There were significant differences in the L 50 estimates between the sexes of red snappers (t-test; all P < 0.05); at least half of the population of males were mature at 270–280 mm compared with 350–370 mm for females (Table 2, Fig. 5). However the length at which half of the population had reached sexual maturity was similar for both species in northern Australia (t-test; all P > 0.05). The L 50 estimates for the eastern Indonesian populations were less well defined. The logistic functions for L. erythropterus females and male L. malabaricus from eastern Indonesia showed a poor fit to these data (r 2 = 0.49 and 0.73) (Fig. 5). This is likely due to smaller sample numbers available for these eastern Indonesian populations and therefore L 50 was not able to be estimated for these populations. The slope parameter (K) for female L. malabaricus in eastern Indonesia was similar to that of the northern Australian L. malabaricus females (t-test; P > 0.05) but differed between the L. erythropterus male populations (t-test; P < 0.05) (Fig. 5). However, the L 50 estimates were significantly higher for both of these population in eastern Indonesia compared with the northern Australian populations (t-test; P < 0.05).

Plot of proportion of mature fish in each 20-mm standard length class for the red snappers, L. erythropterus and L. malabaricus. Logistic curves fitted to data are shown with 95% confidence limits and regression coefficients. Dashed lines indicate the estimated L 50
Batch fecundity
There was no direct positive relationship between batch fecundity and fish length over the size range sampled for either species of red snapper from northern Australia or eastern Indonesia (Fig. 6). Maximum batch fecundity of female L. erythropterus was 640,300 and 676,100 oocytes and for female L. malabaricus was 875,300 and 997,000 oocytes from northern Australia and eastern Indonesia, respectively. However, the mean batch fecundity estimates were not significantly different between the species of red snapper within regions or for a species across regions (t-test; all P > 0.05) (Table 2). While the relative fecundities were not statistically different between the two species or between L. erythropterus populations between regions (t-test; all P > 0.05), female L. malabaricus from northern Australia showed significantly higher egg counts compared with L. malabaricus females in eastern Indonesia (t-test; t = 3.23, P < 0.01) (Table 2). There were also slight differences between red snapper populations in the maximum size of hydrated oocytes in mature, with spawning (stage V) fish with females of both species from eastern Indonesia having larger oocytes compared with northern Australian fish (Table 3).

Plot of batch fecundity and standard length (mm) for mature, spawning females (stage V) of the red snappers, L. erythropterus and L. malabaricus collected from northern Australia and eastern Indonesia
Age-0 snapper surveys
The abundance and size structure of age-0 snapper collected from Sape (eastern Indonesia) were consistent with the female spawning cycle of the red snapper populations within eastern Indonesia. The monthly sampling at Sape showed two distinct peaks in abundance of age-0 red snappers, in March to June and November (Fig. 7). Mean standard lengths of snapper caught in the surveys were also at a minimum in the months of March and September for both species (Fig. 7). These peaks followed the bimodal spawning cycle for female L. malabaricus from eastern Indonesia by approximately 1 or 2 months (Fig. 4). While abundance estimates were not available for age-0 snapper from northern Australia, their size structure showed a single-modal cycle with the smallest snappers recorded (mean length; 70 mm SL) in May to June (Fig. 7). This was about 2–3 months after the highest spawning activity seen in red snapper populations in northern Australia.

Line plot of mean size (SL, mm) and bar chart of mean abundances (fish/km2) for age-0 red snappers, L. erythropterus and L. malabaricus, caught in the Gulf of Carpentaria (northern Australia) and Sape (eastern Indonesia). Standard error bars indicate ±1 SE
Observations were made on habitat types during the shallow water trawl surveys around Darwin, Gove and Weipa (northern Australia). Surveys were undertaken in consecutive years: 2000–2002 in May or June across either sandy coastal habitats (>80% sand substrate; Gove, Weipa) or silty and coarse sand/rubble estuarine habitats (≤20% sand substrate; Darwin, Weipa). Nearly all of the age-0 red snappers caught in the three northern Australian surveys were L. erythropterus (99%). Most of these fish (98%) were caught in Darwin Harbour and Albatross Bay (Weipa), with mean abundances (±1 SE) of 456 ± 119 fish/km2. These two shallow bays are characterised by mostly silt substrate with some coarse sand or rubble. The mean abundance for age-0 L. erythropterus in the sandy coastal habitats surveyed in Gove and Weipa was significantly lower (5 ± 3 fish/km2). Only one L. malabaricus fish was caught during these three otter trawl surveys. Commercial and research trawl data from offshore Northern Prawn Fishery (NPF) grounds in the Gulf of Carpentaria also recorded low abundances of age-0 red snappers (26 ± 4 fish/km2). Although, these offshore trawl grounds consist predominantly of silt and mud substrate with patches of coarse rubble and low reef structure, they were in deeper waters (15–30 m). In contrast, most of the age-0 snapper collected from Sape were L. malabaricus (91%) with a mean abundance of 312 ± 14 fish/km2 for both species combined. These fish were also caught within a shallow bay characterised by predominantly silty substrate and some coarse sand and rubble.
Discussion
This is the first comparative study on the reproductive biology of the two commercially important red snapper species, L. erythropterus and L. malabaricus, from northern Australia and eastern Indonesia. Our results showed that the northern Australian populations of red snappers demonstrated many similar reproductive characteristics to eastern Indonesian populations, however there were some important differences with these stocks on a species and regional level. The most notable difference was in the timing and intensity of their spawning activity. The northern Australian populations of L. erythropterus and L. malabaricus showed a single-modal cycle in their reproductive activity with spawning occurring over several months of the year. Further, the population of L. erythropterus commenced spawning about 2 months earlier than L. malabaricus, suggesting that there are also interspecies variations in the cues for the onset of spawning. The increase in spawning activity for the red snappers in northern Australia were in the summer months and may have been related to factors such as an increase in water temperature and photoperiod, as suggested for a number of other lutjanid species [10, 18, 19].
The eastern Indonesian red snapper populations displayed a less defined spawning cycle through the year to that of the northern Australian populations. This is possibly an effect of these populations living in a more tropical environment or these populations showing greater variability in their reproductive cycles across the sampling sites of eastern Indonesia. Spatial variation in reproductive seasonality has also been observed for several other tropical lutjanid species. Luckhurst et al. [20] reported that the lane snapper (L. synagris) in Bermuda displayed a summer spawning pattern between May and September, whereas in the more tropical waters around Cuba (Caribbean) this species showed a more extended spawning season between March and September. Also, gray snapper (L. griseus) spawning occurred only in summer in the northern sampling regions along the West Florida shelf but in both summer and winter in more southerly, lower-latitude sampling regions [21].
The Indo-Pacific species also showed similar reproductive patterns to two red snapper species from the northern hemisphere: the Atlantic (L. campechanus) and Pacific red snapper (L. peru) (Table 3). These congeneric species are a significant component of commercial and recreational fishing in the Western Atlantic: the Gulf of Mexico and the eastern coast of the USA [22–24], and the Eastern Pacific: Mexico to Peru [25]. Lutjanus campechanus has been reported to display seasonal spawning within the Gulf of Mexico, from May through September or early October [5, 9–11]. Similarly, along the tropical coast of Guerrero, Mexico (17° latitude) the spawning season for L. peru extends from March to September with a peak in August [26]. These spawning periods were consistent with the spring–summer spawning peaks seen for the red snappers in our study.
There was little spatial variability in the sizes at sexual maturity, with the Indo-Pacific species developing active gonads at a relatively small size (about 250–300 mm), except that males matured slightly earlier than females. Length at maturity was also comparable between the two red snapper species within northern Australia. This has significant fishery management implications for the demersal finfish trawl and line fisheries across northern Australia, where these two targeted species usually occur in mixed schools of similar-sized individuals. The Australian red snapper fisheries are managed using sustainable yield estimates and reference points where an increase in total annual catch above sustainable yield estimates or a continued decline in catch triggers a review in management actions. These sustainable yield estimates were arrived at without some of the biological data on these species and without accurate data from adjacent Indonesian waters [2]. With the likely increase in fishing pressure on these Australian stocks and the high level of fishing from Indonesian fisheries, obtaining additional life-history information such as size at maturity and spawning potential will lead to more accurate stock assessments and management.
Presently, the recreational and commercial minimum size limit for both of these species in Australia is 400 mm TL (or 320 mm SL). Our data indicates that this minimum size limit allows a comparable proportion of the population of both species to spawn before being recruited into the fishery. However, the minimum length should be increased to 450 mm TL (or 360 mm SL) to allow at least half of the population of female red snappers to contribute to future recruitment. Further, as these two species of red snappers are recreationally and commercially fished in a mixed-species fishery, management strategies can presently be applied to both species and regulated as a multispecies stock within regions of northern Australia. The accuracy of some of the L 50 estimates for eastern Indonesian fish were doubtful as the logistic curves were a poor fit to the data for female L. erythropterus and male L. malabaricus, with large variations in the proportion of mature red snapper over most size classes sampled for L. erythropterus females and male L. malabaricus from eastern Indonesia. This may be attributed to small sample numbers or a more complex reproductive pattern for these red snapper populations such as differences in their lengths at maturity among the sampling sites in eastern Indonesia. Further sampling is needed at these sites to identify possible small-scale variations.
There is much published evidence to show that tropical lutjanids are generally highly fecund species [5]. Most tropical lutjanids are also serial spawners, with extended spawning seasons throughout most of the year [5, 8]. This is also true for the Indo-Pacific red snappers where we observed large numbers of oocytes in their ovaries and a large proportion of ripe or spawning females in most months of the year. While their batch fecundity estimates were relatively high, they were also quite variable and not linearly related to body size. Variability in fecundity estimates, even within species, is a common characteristic of many lutjanids [5]; for example, we estimated a 520-mm L. malabaricus to have 29,100 hydrated oocytes, whereas a considerably smaller individual, 460 mm, was estimated to have more than 997,000 oocytes in its ovary. This is most likely due to these species being serial spawners with rapid progression of yolked primary oocytes to fully hydrated oocytes, being spawned within a short period of time. This makes fecundity comparisons with previous studies difficult. In our study, L. malabaricus females had higher maximum batch fecundity estimates than did L. erythropterus females. The range in fecundities estimated for these two red snappers were generally consistent with reported fecundities for similar-sized Lutjanus species. The lane snapper, L. synagris from Trinidad, West Indies, showed counts of between 28,750 and 595,700 oocytes for individuals ranging from 294 to 419 mm (TL) [27]. Also, Rojas [28] reported fecundity estimates for the spotted rose snapper, L. guttatus, from Costa Rica, to be 60,454 and 1,363,000 oocytes for a 319- and 596-mm (TL) female, respectively. The number and size of eggs produced by the Atlantic red snapper, L. campechanus, from the South Atlantic Bight and Gulf of Mexico, was reported to be much higher than estimates for the two Indo-Pacific species (Table 3). Although this species attains a large size (>870 mm TL), the number of eggs per gram of body weight (56 ± 10 oocytes) was considerably less than the smaller-sized red snapper species in our study [9].
The Atlantic and Pacific red snappers also showed many similarities to the two species from our study in their age-0 abundance and size structure patterns (Table 3). Highest catch per unit effort of age-0 L. campechanus was recorded in July to September in the northeast Gulf of Mexico, corresponding to the peak spawning period in May through September or early October observed for this species [11]. They also found that abundances of these age-0 L. campechanus in the northeast Gulf of Mexico were markedly higher (80%) around shell beds with fine sand and mud sediments than clean sand substrates. Other studies reported that the young of both L. campechanus and L. peru were commonly associated with near-shore, soft substrate habitats typical of shrimp trawl grounds [12, 25]. However in the Gulf of Mexico and southeastern Atlantic shrimp trawl fishery, age-0 red snapper (L. campechanus) were reported to account for less than 1% in biomass of bycatch [29].
This is the first published study on abundances and habitats of age-0 red snappers in northern Australia and eastern Indonesia. From the surveys we undertook in Darwin, Gove and Weipa in northern Australia and Sape in eastern Indonesia, young L. erythropterus and L. malabaricus were found in nearshore, turbid waters with silty, muddy or coarse sand substrate and low structures such as coral rubble. Unlike the seine net surveys carried out in Sape, we caught very few L. malabaricus in the otter trawl surveys across northern Australia. This may be due to this species having different nursery habitats to L. erythropterus. However, given the coexistence of young of these two species in eastern Indonesia, it is more probable that it is a result of the differences in the spawning patterns observed for the adults in northern Australia and the timing of the surveys. Catch records from previous research prawn trawling studies in the Gulf of Carpentaria between 1996 and 1998 also indicated that few age-0 red snappers were caught in the deeper water commercial trawl grounds, habitats typically with muddy or silty substrate. Therefore, the economically important prawn fishery within northern Australia would have little impact on the early life-history stages of these red snapper species. Further work is required in determining the importance of these inshore estuarine habitats for age-0 red snapper and in identifying the habitats of age-0 L. malabaricus within northern Australia.
Our study showed that the red snappers of northern Australia and eastern Indonesia showed many reproductive characteristics typical of other similar-sized tropical lutjanids. While there were some distinct differences in the reproductive characteristics for the red snappers between northern Australia and eastern Indonesia, there were few differences between species. Therefore, as these two species of red snappers are recreationally and commercially fished in a mixed-species fishery, management strategies can presently be applied to both species and regulated as a multispecies stock within regions of northern Australia. In addition, these similarities to the eastern Indonesian red snappers suggest that their fisheries face similar management issues and that successful approaches adopted in northern Australia should be considered in eastern Indonesia.
References
Williams DMcB, Russ GR (1994) Review of data on fishes of commercial and recreational fishing interest on the Great Barrier Reef, vol 1. Great Barrier Reef Marine Park Authority Research Publication No. 33
Blaber SJM, Dichmont CM, Buckworth RC, Badrudin, Sumiono B, Nurhakim S, Iskandar B, Fegan B, Ramm DC, Salini JP (2005) Shared stocks of snappers (Lutjanidae) in Australia and Indonesia: integrating biology, population dynamics and socio-economics to examine management scenarios. Rev Fish Biol Fish 15:111–127
Salini JP, Ovenden JR, Street R, Pendrey R, Haryanti, Ngurah (2006) Genetic population structure of red snappers (Lutjanus malabaricus Bloch & Schneider, 1801 and L. erythropterus Bloch, 1790) in central and eastern Indonesia and northern Australia. J Fish Biol 68:217–234
Davis TLO, West GJ (1993) Maturation, reproductive seasonality, fecundity, and spawning frequency in Lutjanus vittus (Quoy and Gaimard) from the North West Shelf of Australia. Fish Bull US 91:224–236
Grimes CB (1987) Reproductive biology of the Lutjanidae: a review. In: Polovina JJ, Ralston S (eds) Tropical snapper and groupers: biology and fisheries management. Westview, Boulder, pp 239–294
Kritzer JP (2004) Sex-specific growth and mortality, spawning season, and female maturation of the stripey bass (Lutjanus carponotatus) on the Great Barrier Reef. Fish Bull 102:94–107
Marriott RJ, Mapstone BD, Begg GA (2007) Age-specific demographic parameters, and their implications for management of the red bass, Lutjanus bohar (Forsskal 1775): a large long-lived reef fish. Fish Res 83:204–215
Shimose T, Tachihara K (2005) Age, growth and maturation of the blackspot snapper Lutjanus fulviflammus around Okinawa Island, Japan. Fish Sci 71:48–55
Collins LA, Johnson AG, Keim CP (1996) Spawning and annual fecundity of the red snapper (Lutjanus campechanus) from the northeastern Gulf of Mexico. In: Arreguin-Sanchez F, Munro JL, Balgos MC, Pauly D (eds) Biology, fisheries and culture of tropical groupers and snappers. International Center for Living Aquatic Resources Management Conference Proceedings No. 48, pp 174–188
Rooker JR, Landry AM Jr, Geary BW, Harper JA (2004) Assessment of a shell bank and associated substrates as nursery habitat of postsettlement red snapper. Estuar Coast Shelf Sci 59:653–661
Szedlmayer ST, Conti J (1999) Nursery habitats, growth rates, and seasonality of age-0 red snapper, Lutjanus campechanus, in the northeast Gulf of Mexico. Fish Bull 97:626–635
Workman IK, Foster DG (1994) Occurrence and behavior of juvenile red snapper, Lutjanus campechanus, on commercial shrimp fishing grounds in the northeastern Gulf of Mexico. Mar Fish Rev 56:9–11
Workman I, Shah A, Foster D, Hataway B (2002) Habitat preferences and site fidelity of juvenile red snapper (Lutjanus campechanus). ICES J Mar Sci 59:S43–S50
Andamari R, Milton DA, Van der Velde T, Sumiono B (2004) Reproductive biology of red snapper (Lutjanus malabaricus) from Sape and Kupang waters. Indones Fish Res J 10:65–75
Blaber SJM, Brewer DT, Milton DA, Merta GS, Efizon D, Fry G, van der Velde T (1999) The life history of the protandrous tropical shad Tenualosa macrura (Alosinae: Clupeidae): fishery implications. Estuar Coast Shelf Sci 49:689–701
Cyrus DP, Blaber SJM (1984) The reproductive biology of Gerres in Natal estuaries. J Fish Biol 24:491–504
King M (1995) Fisheries biology. Assessment and management. Fishing News Books, Oxford
Arnold CR, Wakeman JR, Williams TD, Teece GD (1978) Spawning of red snapper (Lutjanus campechanus) in captivity. Aquaculture 15:301–302
Grimes CB, Huntsman GR (1980) Reproductive biology of the vermilion snapper, Rhomboplites aurorubens, from North Carolina and South Carolina. Fish Bull 78:137–146
Luckhurst BE, Dean JM, Reichert M (2000) Age, growth and reproduction of the lane snapper Lutjanus synagris (Pisces: Lutjanidae) at Bermuda. Mar Ecol Prog Ser 203:255–261
Allman RJ, Grimes CB (2002) Temporal and spatial dynamics of spawning, settlement, and growth of gray snapper (Lutjanus griseus) from the West Florida shelf as determined from otolith microstructures. Fish Bull 100:391–403
Manooch CSIII, Potts JC, Vaughan DS, Burton ML (1998) Population assessment of the red snapper from the southeastern United States. Fish Res 38:19–32
Szedlmayer ST, Shipp RL (1994) Movement and growth of red snapper, Lutjanus campechanus, from an artificial reef area in the northeastern Gulf of Mexico. Bull Mar Sci 55:887–896
Wilson CA, Nieland DL (2001) Age and growth of red snapper, Lutjanus campechanus, from the northern Gulf of Mexico off Louisiana. Fish Bull 99:653–664
Rocha-Olivares A (1998) Age, growth mortality, and population characteristics of the Pacific red snapper, Lutjanus peru, off the southeast coast of Baja California, Mexico. Fish Bull 96:562–574
Santamaria-Miranda A, Elorduy-Garay JF, Villalejo-Fuerte M, Rojas-Herrera AA (2003) Gonadal development and reproductive cycle of Lutjanus peru (Pisces: Lutjanidae) in Guerrero, Mexico. Rev Biol Trop 51:489–501
Manickchand-Dass S (1987) Reproduction, age and growth of the lane snapper, Lutjanus synagris (Linnaeus), in Trinidad, West Indies. Bull Mar Sci 40:22–28
Rojas JR (1997) Fecundity and reproductive seasons of Lutjanus guttatus (Pisces: Lutjanidae) in Gulf of Nicoya, Costa Rica. Rev Biol Trop 45:477–487
Scott-Denton E, Nance J (1996) Shrimp trawl bycatch research in the US Gulf of Mexico and southeastern Atlantic. In: Arreguin-Sanchez F, Munro JL, Balgos MC, Pauly D (eds) Biology, fisheries and culture of tropical groupers and snappers. International Center for Living Aquatic Resources Management Conference Proceedings No. 48, pp 360–362
Acknowledgments
We thank the eastern Indonesian fisheries staff; Mujimin and Noto for coordinating and collecting red snapper samples in eastern Indonesia. Stephanie Boubaris, Mike Elmsley, Chris Errity and Chris Tarca from Northern Territory Department of Business, Industry and Resource Development helped collect the northern Australian samples. We would also like to thank Don Heales and Shane Griffiths for coordinating and participating in field surveys in northern Australia. We are also grateful to Bill Passey of FV Ocean Harvest and Clive Perry of FV Reef Venture for supplying red snapper samples from northern Australia. Steve Blaber and John Salini made constructive comments on the draft manuscript. This project was partly funded by the Australian Centre for International Agricultural Research (ACIAR FIS 97/165).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fry, G., Milton, D.A., Van Der Velde, T. et al. Reproductive dynamics and nursery habitat preferences of two commercially important Indo-Pacific red snappers Lutjanus erythropterus and L. malabaricus . Fish Sci 75, 145–158 (2009). https://doi.org/10.1007/s12562-008-0034-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12562-008-0034-4