
Abstract
The purpose of this study was to examine the effects of conceptual similarity between target and nontarget stimuli on the P300 component of event-related potentials using a multi-stimulus oddball paradigm. The stimuli were pictures of three objects and their corresponding Chinese and English words. The pictures and their corresponding Chinese and English words were considered to the target stimulus, respectively. The experiments elucidate that imagery and verbal information are processed by distinct but interconnected systems.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Event-related potential (ERP) has been used extensively in both clinical and experimental studies [1, 5, 12, 17, 23, 32–37]. The P300 is an endogenous component of ERP; changes in P300 amplitude, latency, and scalp topography have been correlated with a number of psychometric and cognitive variables [11, 29–31]. The most frequently used paradigm in P300 research has been the two-stimulus oddball task, in which a participant must discriminate between infrequent target and frequent nontarget stimuli. In these paradigms, the infrequent, task-relevant target stimulus is associated with large P300 components with maximal amplitude at posterior electrodes. A major difference between our paradigm and previous paradigms is that we have used multiple stimuli that all occur with the same probability. This allows us to use the ERP to index the brain’s spontaneous categorization of stimuli.
Although ERP studies of semantic priming have often focused on the N400, the P300 also appears to be sensitive to the effect. For example, Bentin et al. [4] reported that P300 latency is significantly reduced for semantically primed targets. Targets were also associated with increased P300 amplitude. More recently, it has been demonstrated that the P300 is also useful as an index of the degree of semantic relatedness between primes and targets. The authors [18] found increased P300 amplitude for targets that had a direct semantic relationship with the prime, as well as for targets that had only an indirect semantic relationship with the prime compared to targets that were unrelated to the prime and pseudo-word targets. Similarly, McPherson and Holcomb [25] demonstrated that long latency ERP negativities can differentiate the level of relationship between primes and targets using pictorial stimuli. The results of Experiment 1 of the present study extend these findings by suggesting that the degree of similarity between stimuli can be indexed physiologically using a multi-stimulus oddball paradigm.
Recently, several researchers explored the properties of nontarget P300 in three-stimulus paradigms [7–10, 20, 21, 24, 34]. It was found that target and nontarget P300s are differentially sensitive to the effects of stimulus context. Although the amplitude of the target P300 is influenced by its similarity to frequent standard stimuli, it is unaffected by the perceptual aspects of infrequent nontarget stimuli. However, P300s to infrequent nontarget stimuli are influenced by the perceptual context of both target and infrequent standard stimuli. It has also been shown that for auditory stimuli, the perceptual distinctiveness of the infrequent nontarget and target stimuli directly influences the amplitude and topography of nontarget P300s. In summary, these studies have demonstrated that the relationship between nontarget and target stimuli affects the properties of nontarget P300s.
A drawback of this previous study was that it was difficult to distinguish the effects of conceptual and perceptual/physical similarity between a target and nontarget stimuli. Similar-to-target stimuli were both conceptually and perceptually related to the target stimuli. For instance, while target and nontarget stimuli shared features that are commonly considered conceptual, such as race (black/white) and gender (male/female), they also shared common perceptual features such as color contrast, facial morphology, and hair length.
Three counterbalanced studies (“picture” as target, “English word” as target and “Chinese word” as target) were performed in the current study. In the first study, the picture “bag” was designated as the target stimulus. We hypothesized that the target stimulus (picture of bag) would elicit a prominent P300 component. We also hypothesized that the related nontarget stimulus (Chinese word and English word of bag) would elicit a significant nontarget P300. The remaining nontarget stimuli were both perceptually and conceptually distinct from the target and therefore failed to generate a prominent P300 effect. In the experiment 2, the Chinese word of bag was designated as the target stimulus. We hypothesized that the target stimulus (Chinese word of bag) would elicit a prominent P300 component, and the related nontarget stimulus (picture and English word of bag) would elicit a significant nontarget P300. The remaining nontarget stimuli failed to generate a prominent P300 effect. In the experiment 3, the English word of bag was designated as the target stimulus. We hypothesized that the target stimulus (English word of bag) would elicit a prominent P300 component, and the related nontarget stimulus (picture and Chinese word of bag) would elicit a significant nontarget P300. The remaining nontarget stimuli failed to generate a prominent P300 effect.
The purpose of the current studies was to examine the effects of conceptual similarity between target and nontarget stimuli on nontarget P300 amplitude. In this paper, we found that target stimuli elicited prominent P300 components. Nontarget stimuli that were similar to the target exhibited P300s that were similar to but smaller in amplitude than those elicited by target stimuli. P300 amplitudes for both target and similar-to-target stimuli were greater than for stimuli that were dissimilar to the target. These data suggested that P300 amplitude is a useful index of similarity between a target and nontarget stimuli. As opposed to the results of studies using the three-stimulus oddball paradigm, the changes in amplitude of nontarget P300s seen in this study were independent of the effects stimulus probability.
Methods
Participants
Twelve right-handed students (four females) from Blue Sky University and ranging in age from 18 to 23 years (mean = 20) participated in the study. All participants were known and approved the experimental schemes, and the experimental schemes were authorized by Blue Sky University Academic Committee. All participants were native Chinese speakers and had normal or corrected to normal vision. Two participant’s data were excluded because he failed to follow the experimental instructions.
Stimuli
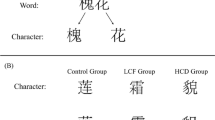
Stimuli consisted of three object pictures and their conceptual-related Chinese words and English words (Fig. 1). The picture of bag, Chinese word of bag and English word of bag were designated as the target stimulus in each experiment, respectively. Every experiment consisted of two blocks. The stimuli were presented in a pseudorandom fashion with 180 stimuli in each block, and all stimuli were presented with equal probability (0.17).

Words and pictures stimuli used in the study
Recording
Electroencephalogram (EEG) was recorded using 32-channel NeuroScan system. Recordings were referenced to electronically linked mastoid electrodes. It was recorded using a gain of 1,000 and a bandpass of 0.05–100 Hz. The horizontal EOG was monitored from electrodes at the outer canthi of the eyes, and the vertical EOG was monitored above and below the orbital region of the left eye. Electrode impedances did not exceed 10 kΩ. EEG and EOG activity were recorded using a gain of 1,000 and a bandpass of 0.1–30 Hz. To eliminate artifacts, trials with EEG voltages exceeding ±70 μV were rejected from the average. Approximately 10–15% of trials were excluded due to EOG or other artifacts.
Procedure
This study was conducted in a sound-attenuating chamber. Once the electrode cap had been fitted, participants were seated comfortably in a chair in front of the stimulus presentation monitor. Prior to each block of stimuli, participants were instructed to keep a silent mental count of the number of times the target appeared on the screen. Following each block, we asked the participants to report this count. No instructions were given with regards to any of the nontarget stimuli. Stimuli were presented on a 17-inches CRT monitor. Participants were seated 0.5 m from the CRT monitor, and the center of the screen was at eye level. To minimize eye movements, participants were instructed to attend to a centered fixation point that was displayed for the duration of the experiment. Participants were allowed to rest between blocks.
Experiments
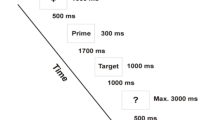
The research contained three experiments; each experiment includes two blocks, which were presented in a pseudorandom fashion with 180 stimuli in each block. Figure 2 shows schematic diagram of stimulus procedure, stimuli were presented for 250 ms with an interstimulus interval of 1,000 ms. All stimuli were presented in with equal probability (0.17).

Schematic diagram of stimulus procedure
The experiment order and the target, the related nontarget and the unrelated nontarget are listed in the Table 1. In experiment 1, the target was the picture of the bag, the Chinese word of bag and the English word “bag” were considered to be the related nontarget. This experiment includes two blocks, in the first block, we use the picture of “bag” as target, the Chinese word was considered to be the related nontarget, the other pictures and Chinese words of “car” and “cup” were considered to be the nontarget; in the second block, we use the picture of “bag” as target, the English word was considered to be the related nontarget, the other pictures and English words of “car” and “cup” were considered to be the nontarget.
Results
Behavioral Data
Participants were very accurate in counting the number of targets. For all conditions, error rate of participant reporting a count was lower than 3%. To reduce the number of statistical comparisons made in this study, while allowing for analysis of differences in the anterior/posterior and hemispheric dimensions, the data from only nine electrodes were statistically analyzed. The electrodes selected correspond to the International 10–20 system sites F3, FZ, F4, C3, CZ, C4, P3, PZ, and P4. Data were analyzed using analysis of variance (ANOVA). The variables examined were anterior vs. posterior distribution (frontal, central, parietal) and stimulus types.
Experiment 1
In experiment 1, the target was the picture of “bag”, the Chinese word of bag and the English word “bag” were considered to be the related nontarget. This experiment includes two blocks, in the first block, we use the picture of “bag” as target, the Chinese word was considered to be the related nontarget; in the second block, we use the picture of “bag” as target, the English word was considered to be the related nontarget.
Figure 3 displays grand average waveforms for the target (picture of bag), related nontarget stimuli (Chinese word of bag), and the remaining nontarget stimuli at nine electrode sites. The target stimulus elicited a large P300 response that was maximal over the vertex, but the related nontarget and remaining nontarget stimuli failed to generate a prominent P300 effect.

Grand average waveforms for the target (picture of bag), related nontarget (Chinese word of bag), and other nontarget stimuli
Figure 4 displays 32-channel ERP distribution maps for the target, related nontarget, and the other nontarget stimuli. The ERP distribution maps are taken at latency (405 ms) of peak P300 amplitude for target stimulus. We can see that there are obvious brain activities on the posterior scalp for the target stimulus, for the related nontarget stimulus and remaining nontarget stimuli, there are no obvious brain activities over the scalp.

ERP distribution maps taken at latency of peak P300 amplitude for target stimulus
In the second block of the experiment 1, the target is the picture of bag and the related nontarget is the English word of bag. Figure 5 displays grand average waveforms for the target (picture of bag), related nontarget stimuli (English word of bag), and the remaining nontarget stimuli at nine electrode sites. The target stimulus elicited a large P300 response that was maximal over the vertex, but the related nontarget and remaining nontarget stimuli failed to generate a prominent P300 effect.

Grand average waveforms for the target (picture of bag), related nontarget (English word of bag), and other nontarget stimuli
Figure 6 displays 32-channel ERP distribution maps for the target, related nontarget, and the other nontarget stimuli. The ERP distribution maps are taken at latency (400 ms) of peak P300 amplitude for target stimulus. We can see that there are obvious brain activities on the posterior scalp for the target stimulus, for the related nontarget stimulus and remaining nontarget stimuli, there are no obvious brain activities over the scalp.

ERP distribution maps taken at latency of peak P300 amplitude for target stimulus
A significant main effect was found for anterior/posterior electrode EEG amplitude distribution, F(2,20) = 20.35, P = 0.001. In the first block of experiment 1, simple comparisons revealed that peak of P300 amplitude between 375 and 600 ms was greater at central electrode sites (10.31 ± 0.45 μV) than at frontal electrode sites (6.30 ± 0.43 μV), F(1,10) = 46.13, P < 0.01, and parietal electrode sites (10.32 ± 0.65 μV), F(1,10) = 19.35, P < 0.01. A significant main effect for stimulus type was found, F(8,80) = 10.63, P < 0.01. The target (picture of bag) elicited a larger P300 amplitude (10.31 ± 0.45 μV) compared with related nontarget (Chinese word of bag: 0.65 ± 0.31 μV), as well as other nontarget.
In the second block of experiment 1, simple comparisons revealed that peak of P300 amplitude between 375 and 600 ms was greater at central electrode sites (12.71 ± 0.28 μV) than at frontal electrode sites (11.30 ± 0.65 μV), F(1,10) = 20.46, P < 0.01, and parietal electrode sites (11.72 ± 0.46 μV), F(1,10) = 14.83, P < 0.01. A significant main effect for stimulus type was also found, F(8,80) = 13.41, P < 0.01. The target (picture of bag) elicited a larger P300 amplitude (12.71 ± 0.28 μV) compared with related nontarget (English word of bag: 1.48 ± 0.25 μV), as well as other nontarget.
Experiment 2
In experiment 2, the target was the Chinese word of bag, the picture of “bag” and the English word were considered to be the related nontarget. This experiment includes two blocks, in the first block, we use the Chinese word of bag as target, the bag picture was considered to be the related nontarget; in the second block, we use the Chinese word of bag as target, the English word was considered to be the related nontarget.
Figure 7 displays grand average waveforms for the target (Chinese word of bag), related nontarget stimuli (picture of bag), and the remaining nontarget stimuli at nine electrode sites. The target stimulus and the related nontarget elicited a large P300 response that was maximal over the vertex, but the remaining nontarget stimuli failed to generate a prominent P300 effect.

Grand average waveforms for the target (Chinese word of bag), related nontarget (picture of bag), and other nontarget stimuli
Figure 8 displays 32-channel ERP distribution maps for the target, related nontarget, and the other nontarget stimuli. The ERP distribution maps are taken at latency (415 ms) of peak P300 amplitude for target stimulus. We can see that there are obvious brain activities on the posterior scalp for the target stimulus, there are some brain activities on the posterior scalp for the related nontarget stimulus, but for the remaining nontarget stimuli, there are no obvious brain activities over the scalp.

ERP distribution maps taken at latency of peak P300 amplitude for target stimulus
In the second block of the experiment 2, the target is the Chinese word of bag and the related nontarget is the English word of bag. Figure 9 displays grand average waveforms for the target (Chinese word of bag), related nontarget stimuli (English word of bag), and the remaining nontarget stimuli at nine electrode sites. The target stimulus elicited a large P300 response that was maximal over the vertex, but the related nontarget and remaining nontarget stimuli failed to generate a prominent P300 effect.

Displays grand average waveforms for the target (Chinese word of bag), related nontarget (English word of bag), and other nontarget stimuli
Figure 10 displays 32-channel ERP distribution maps for the target, related nontarget, and the other nontarget stimuli. The ERP distribution maps are taken at latency (410 ms) of peak P300 amplitude for target stimulus. We can see that there are obvious brain activities on the posterior scalp for the target stimulus, for the related nontarget stimulus and remaining nontarget stimuli, there are no obvious brain activities over the scalp.

ERP distribution maps taken at latency of peak P300 amplitude for target stimulus
A significant main effect was found for anterior/posterior electrode distribution, F(2,20) = 18.41, P = 0.001. In the first block of experiment 2, simple comparisons revealed that peak of P300 amplitude between 375 and 600 ms was greater at central electrode sites (6.69 ± 0.71 μV) than at frontal electrode sites (5.36 ± 0.32 μV), F(1,10) = 35.17, P < 0.01, and parietal electrode sites (6.43 ± 0.68 μV), F(1,10) = 9.35, P < 0.01. A significant main effect for stimulus type was found, F(8,80) = 8.42, P < 0.01. The target (Chinese word of bag) elicited a larger P300 amplitude (6.69 ± 0.71 μV) compared with related nontarget (picture of bag: 4.04 ± 0.56 μV), as well as other nontarget.
In the second block of experiment 2, simple comparisons revealed that peak of P300 amplitude between 375 and 600 ms was greater at central electrode sites (7.24 ± 0.55 μV) than at frontal electrode sites (5.30 ± 0.43 μV), F(1,10) = 17.68, P < 0.01, and parietal electrode sites (7.15 ± 0.46 μV), F(1,10) = 9.12, P < 0.01. A significant main effect for stimulus type was also found, F(8,80) = 31.53, P < 0.01. The target (Chinese word of bag) elicited a larger P300 amplitude (7.24 ± 0.55 μV) compared with related nontarget (English word of bag: 1.66 ± 0.35 μV), as well as other nontarget.
Experiment 3
In experiment 3, the target was the English word “bag”, and the picture and the Chinese word were considered to the related nontarget, others were nontarget. This experiment includes two blocks, in the first block, we use the English word “bag” as target, the bag picture was considered to be the related nontarget; in the second block, we use English word “bag” as target, the Chinese word was considered to be the related nontarget.
Figure 11 displays grand average waveforms for the target (english word of bag), related nontarget stimuli (picture of bag), and the remaining nontarget stimuli at nine electrode sites. The target stimulus and the related nontarget elicited a large P300 response that was maximal over the vertex, but the remaining nontarget stimuli failed to generate a prominent P300 effect.

Grand average waveforms for the target (English word of bag), related nontarget (picture of bag), and other nontarget stimuli
Figure 12 displays 32-channel ERP distribution maps for the target, related nontarget, and the other nontarget stimuli. The ERP distribution maps are taken at latency (439 ms) of peak P300 amplitude for target stimulus. We can see that there are obvious brain activities on the posterior scalp for the target stimulus, there are some brain activities on the posterior scalp for the related nontarget stimulus, but for the remaining nontarget stimuli, there are no obvious brain activities over the scalp.

ERP distribution maps taken at latency of peak P300 amplitude for target stimulus
In the second block of the experiment 2, the target is the English word of bag and the related nontarget is the Chinese word of bag. Figure 13 displays grand average waveforms for the target (English word of bag), related nontarget stimuli (Chinese word of bag), and the remaining nontarget stimuli at nine electrode sites. The target stimulus elicited a large P300 response that was maximal over the vertex, but the related nontarget and remaining nontarget stimuli failed to generate a prominent P300 effect.

Grand average waveforms for the target (English word of bag), related nontarget (Chinese word of bag), and other nontarget stimuli
Figure 14 displays 32-channel ERP distribution maps for the target, related nontarget, and the other nontarget stimuli. The ERP distribution maps are taken at latency (441 ms) of peak P300 amplitude for target stimulus. We can see that there are obvious brain activities on the posterior scalp for the target stimulus, for the related nontarget stimulus and remaining nontarget stimuli, there are no obvious brain activities over the scalp.

ERP distribution maps taken at latency of peak P300 amplitude for target stimulus
A significant main effect was found for anterior/posterior electrode distribution, F(2,20) = 16.76, P = 0.001. In the first block of experiment 3, simple comparisons revealed that peak of P300 amplitude between 375 and 600 ms was greater at central electrode sites (6.08 ± 0.13 μV) than at frontal electrode sites (4.60 ± 0.43 μV), F(1,10) = 38.32, P < 0.01, and parietal electrode sites (5.26 ± 0.30 μV), F(1,10) = 16.28, P < 0.01. A significant main effect for stimulus type was found, F(8,80) = 9.67, P < 0.01. The target (English word of bag) elicited a larger P300 amplitude (6.08 ± 0.13 μV) compared with related nontarget (picture of bag: 3.35 ± 0.54 μV), as well as other nontarget.
In the second block of experiment 3, simple comparisons revealed that peak of P300 amplitude between 375 and 600 ms was greater at central electrode sites (7.51 ± 0.28 μV) than at frontal electrode sites (6.70 ± 0.67 μV), F(1,10) = 18.37, P < 0.01, and parietal electrode sites (7.22 ± 0.44 μV), F(1,10) = 9.45, P < 0.01. A significant main effect for stimulus type was also found, F(8,80) = 5.68, P < 0.01. The target (English word of bag) elicited a larger P300 amplitude (7.51 ± 0.28 μV) compared with related nontarget (Chinese word of bag: 0.48 ± 0.15 μV), as well as other nontarget.
Discussion
These studies examined the ERP correlates of processing a nontarget stimulus that is conceptually related to a target. The result of experiment 1 did not conform to our hypothesis. In experiment 1, the target stimulus (picture of bag) produced a larger P300; the related nontarget stimuli (English word and Chinese word of bag) did not produce a nontarget P300 response. Distinct processing circuits for imagery (pictures) and verbal information could account for the differential effects of picture and word targets reported in this experiment 1. Previous work has found similar dissociations. Glaser and Glaser [16] suggest that word stimuli have “privileged access” to the lexicon, whereas pictorial stimuli have “privileged access” to the semantic network. The authors found that when engaged in semantic categorization of words in a modified Stroop task, a picture from another semantic category causes strong inhibition. However, when participants were required to categorize pictures, words from another semantic category caused no inhibition [15, 16]. It has also been demonstrated that ERP indices of cross-modal priming can be one directional (e.g., [13, 22]). Interestingly, Kazmerski and Friedman found that in the cross-form (word-picture or picture-word) condition of a direct recognition memory task, significant ERP repetition priming effects were found when pictorial targets were previously seen as words, but not when word targets were previously seen as pictures.
Interestingly, the result of the first block of experiment 2 supported the conclusion that nontarget P300 amplitude is correlated with the similarity between a target and nontarget stimulus. More specifically, the data indicated that nontarget P300 amplitude is affected when a nontarget stimulus activates the same mental representation as the target stimulus. This effect appears to be independent of perceptual similarities between target and nontarget stimuli. Comparing to the first block of experiment 1, the Chinese word of bag related to target picture of bag failed to elicit the P300. These data could be interpreted in light of theories such as the dual-coding model [28], and it suggested that Chinese words and pictures can be processed independently, even if they share a common meaning. The interconnections could lead to the cross-activation of corresponding units of information held in the two independent systems, although this cross-activation does not necessarily always occur. A number of previous ERP studies have supported the hypothesis that pictures and words are processed differently. In the second block of experiment 2, the target stimulus (Chinese word of bag) produced a larger P300; but the related nontarget stimulus (English word of bag) did not produce a nontarget P300 response. Friedman et al. [13] and Kazmerski and Friedman [22] found that in the cross-form (word-picture or picture-word) condition of a direct recognition memory task, significant ERP repetition priming effects were found when pictorial targets were previously seen as words, but not when word targets were previously seen as pictures.
The result of the first block of experiment 3 is similar to the result of the first block of experiment 2; the target stimulus (Chinese word and English word of bag) and the related nontarget (picture of bag) stimuli generate a prominent P300. But in the second block of experiment 3, only the target stimulus (Chinese word of bag) can generate a prominent P300, the related nontarget stimulus (English word of bag) cannot generate a prominent P300. Comparing to the second block of experiment 2, no matter whether the target stimulus is Chinese or English, the related verbal nontarget (Chinese word or English word) cannot generate a prominent P300. Therefore, we can draw a conclusion that the English word and Chinese word share a common meaning and are processed uniformly.
The results of experiment 2 and experiment 3 supported our hypothesis that the nontarget P300 amplitude is correlated with the level of similarity between a target and nontarget stimulus. More specifically, the data indicated that nontarget P300 amplitude is affected when a nontarget stimulus activates the same mental representation as the target stimulus. This effect appears to be independent of perceptual similarities between target and nontarget stimuli. Overall, the data from experiment 2 and experiment 3 indicate that nontarget P300 amplitude is a potentially useful index of how the brain defines relationships between perceptually distinct stimuli. The results of experiment 2 and experiment 3 are consistent with the hypothesis that ERPs can be used to index both voluntary and involuntary components of the human attention and memory systems. For example, numerous studies have demonstrated that semantic priming affects the amplitude and/or latency of several ERP components (e.g., [3, 4, 6, 26, 27]). Many of these studies utilized a lexical decision task, a classic experimental paradigm in which participants are required to decide if a target word is a real word or a pseudo-word. If the target word is preceded by a semantically related (as opposed to an unrelated) word, then reaction times for the lexical decision are faster. ERPs have been shown to be a useful physiological marker of this effect. Semantically primed targets have consistently been shown to elicit a significantly smaller N400 than an unrelated target. These effects have also been demonstrated using semantic priming with line drawings (e.g., [2, 19]) and, similar to the current studies, for cross-modal priming using line drawings and words (e.g., [14, 26]).
Although the results of experiment 1 seem to support cognitive models that propose separate processing streams for pictorial and verbal stimuli, it is important to note that the study was not specifically designed to test these hypotheses. Therefore, the interpretation of the data in this light should be regarded as speculative. However, taken with the results of experiment 2 and experiment 3, the data do suggest that the nontarget P300 amplitude recorded in a multi-stimulus oddball paradigm might not only be a useful index of similarity between perceptually distinct stimuli, but also as a physiological marker of how the brain might process stimuli that activate similar mental representations differently. This opens the way for more directed use of the paradigm in testing hypotheses such as those proposed by the dual-coding model.
References
Barber HA, Kutas M. Interplay between computational models and cognitive electrophysiology in visual word recognition. Brain Res Rev. 2007;53(1):98–123.
Barrett SE, Rugg MD. Event-related potentials and the semantic matching of pictures. Brain Cogn. 1990;14:201–12.
Bentin S. Event-related potentials, semantic processes, and expectancy factors in word recognition. Brain Lang. 1987;31:308–27.
Bentin S, McCarthy G, Wood CC. Event-related potentials, lexical decision, and semantic priming. Electroencephalogr Clin Neurophysiol. 1985;60:343–55.
Braun M, Jacobs AM, Hahne A, et al. Model-generated lexical activity predicts graded ERP amplitudes in lexical decision. Brain Res. 2006;1073–1074:431–9.
Chwilla DJ, Brown CM, Hagoort P. The N400 as a function of the level of processing. Psychophysiology. 1995;32:274–85.
Comerchero MD, Polich J. P3a, perceptual distinctiveness, and stimulus modality. Cogn Brain Res. 1998;7:41–8.
Comerchero MD, Polich J. P3a and P3b from typical auditory and visual stimuli. Clin Neurophysiol. 1999;110:24–30.
Courchesne E, Courchesne RY, Hillyard SA. The effect of stimulus deviation on P3 waves to easily recognized stimuli. Neuropsychologia. 1978;16:189–99.
Courchesne E, Hillyard SA, Galambos R. Stimulus novelty, task relevance and the visual evoked potential in man. Electroencephalogr Clin Neurophysiol. 1975;39:131–43.
Donchin E, Coles MGH. Precommentary: is the P300 component manifestation of context updating? Behav Brain Sci. 1988;11:357–74.
Fay S, Isingrini M, Ragot R, et al. The effect of encoding manipulation on word-stem cued recall: an event-related potential study. Cogn Brain Res. 2005;24(3):615–26.
Friedman D, Cycowicz YM, Dziobek I. Cross-form conceptual relations between sounds and words: effects on the novelty P3. Cogn Brain Res. 2003;18:58–64.
Ganis G, Kutas M, Sereno M. The search for “commonsense”: an electrophysiological study of the comprehension of words and pictures in reading. J Cogn Neurosci. 1996;8:89–106.
Glaser WR, Dungelhoff F-J. The time course of picture—word interference. J Exp Psychol Hum Percept Perform. 1984;10:640–54.
Glaser WR, Glaser MO. Context effects in stroop-like word and picture processing. J Exp Psychol Gen. 1989;118:13–42.
Hauk O, Davis MH, Ford M, et al. The time course of visual word recognition as revealed by linear regression analysis of ERP data. Neuroimage. 2006;30(4):1383–400.
Hill H, Strube M, Roesch-Ely D, Weisbrod M. Automatic vs. controlled processes in semantic priming-differentiation by event-related potentials. Int Psychophysiol. 2002;44:197–218.
Holcomb PJ, McPherson WB. Event-related potentials reflect semantic priming in an object decision task. Brain Cogn. 1994;24:257–76.
Katayama J, Polich J. P300 probability, and the three-tone paradigm. Electroencephalogr Clin Neurophysiol. 1996;100:555–62.
Katayama J, Polich J. Stimulus context determines P3a and P3b. Psychophysiology. 1998;35:23–33.
Kazmerski VA, Friedman D. Old/new differences in direct and indirect memory tests using pictures and words in within and cross-forms conditions: event-related potential and behavioral measures. Cogn Brain Res. 1997;5:255–72.
Kincses TZ, Chadaide Z, Varga ET, et al. Task-related temporal and topographical changes of cortical activity during ultra-rapid visual categorization. Brain Res. 2006;1112(1):191–200.
Knight RT. Decreased response to novel stimuli after prefrontal lesions inman. Electroencephalogr Clin Neurophysiol. 1984;59:133–9.
McPherson WB, Holcomb PJ. An electrophysiological investigation of semantic priming with pictures of real objects. Psychophysiology. 1999;36:53–65.
Nigam A, Hoffman J, Simons R. N400 to semantically anomalous pictures and words. J Cogn Neurosci. 1992;4:15–22.
Niznikiewicz M, Squires NK. Phonological processing and the role of strategy in silent reading: behavioral and electrophysiological evidence. Brain Lang. 1996;52:342–64.
Paivio A. Dual coding theory: retrospect and current status. Can J Psychol. 1991;45:255–87.
Picton TW. The P300 wave of the human event-related potential. J Clin Neurophysiol. 1992;9:456–79.
Pritchard WS. Psychophysiology of P300. Psychol Bull. 1981;89:506–40.
Ritter W, Vaughan HG, Costa LD. Orienting and habituation to auditory stimuli: a study of short term changes in average evoked responses. Electroencephalogr Clin Neurophysiol. 1968;25:550–6.
Schiller NO. Lexical stress encoding in single word production estimated by event-related brain potentials. Brain Res. 2006;1112(1):201–12.
Sitnikova T, West WC, Kuperberg GR, et al. The neural organization of semantic memory: electrophysiological activity suggests feature-based segregation. Biol Psychol. 2006;71(3):326–40.
Soltani M, Knight RT. Neural origins of the P300. Crit Rev Neurobiol. 2000;14:199–224.
Watson TD, Azizian A, Berry S, et al. Event-related potentials as an index of similarity between words and pictures. Psychophysiology. 2005;42(4):361–8.
Xia ZJ, Zhao XD, et al. Comparison of P300 evoked by image, photo and Chinese character stimulation in normal young people. Chin J Clin Neurol. 2004;17(3):167–8.
Zhang Q, Guo CY, Ding JH, et al. Concreteness effects in the processing of Chinese words. Brain Lang. 2006;96(1):59–68.
Acknowledgments
Supported by Foundation of Jiangxi Educational Committee (No. GJJ09622) and Natural Science Foundation of Jiangxi Bluesky University of Jiangxi Province of China (No. XYKJ08YB06).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hu, J., He, K. & Xiong, J. Comparison of Event-Related Potentials Between Conceptually Similar Chinese Words, English Words, and Pictures. Cogn Comput 2, 50–61 (2010). https://doi.org/10.1007/s12559-009-9025-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12559-009-9025-8