
Abstract
Semantic priming in Turkish was examined in 36 right-handed healthy participants in a delayed lexical decision task via taxonomic relations using EEG. Prime–target relations included related- unrelated- and pseudo-words. Taxonomically related words at long stimulus onset asynchrony (SOA) were shown to modulate N400 and late positive component (LPC) amplitudes. N400 semantic priming effect in the time window of 300–500 ms was the largest for pseudo-words, intermediate for semantically-unrelated targets, and smallest for semantically-related targets as a reflection of lexical-semantic retrieval. This finding contributes to the ERP literature showing how remarkably universal the N400 brain potential is, with similar effects across languages and orthography. The ERP data also revealed different influences of related, unrelated, and pseudo-word conditions on the amplitude of the LPC. Attention scores and mean LPC amplitudes of related words in parietal region showed a moderate correlation, indicating LPC may be related to “relationship-detection process”.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The language system is one of the most competent products of the human brain and the functioning of its components lead the most intriguing questions in neuroscience. A fundamental question is how word knowledge is organized, stored, and recalled in the brain. Whether written or spoken, in the process of language production and language comprehension, the brain needs to have and utilize mental representations of words. The phonological, semantic, morphological, and orthographic information of words are represented in our mental lexicon which is defined as a mental dictionary (Bonin, 2004).
Semantic priming in the lexical decision task is a prevalent way to understand the structure and organization of mental lexicon. This method also provides an investigation of cognitive abilities such as perception, working and semantic memory, and attention (Jones & Estes, 2012; Ortells et al., 2006). In the paradigm, a word stimulus (i.e., prime) is presented shortly before a target stimulus and participants have to determine if the target stimulus is a word or a non-word. The prime facilitates processing of the target word when a prime is followed by a semantically-related target word. This facilitation yields a semantic priming effect. Behaviorally, this effect manifests itself as an increase in response accuracy and a reduction in reaction time in semantically-related pairs. In addition to behavioral semantic priming, the N400 is generally regarded as an electrophysiological measurement of semantic priming (Osterhout & Holcomb, 1995). Since the pioneering report of Bentin et al., (1985), numerous studies tended to focus on N400 in a paired stimulus paradigm or list format (e.g., Bentin 1987; Brown et al., 2000; Holcomb, 1988; Holcomb & Neville 1990; Hill et al., 2002; Hill et al., 2005; Landi & Perfetti, 2007; Noldy et al., 1990; Rossell et al., 2003; Rugg, 1984; Wang et al., 2017).
The main theories that elucidated the semantic priming effect include automatic spreading activation (Posner & Snyder, 1975) and strategic (controlled) processes (Neely, 1991). The automatic spreading activation mechanism acts quickly and occurs unconsciously or unintentionally (Posner & Snyder, 1975). The strategic (controlled) processes are either expectancy-based or related to semantic matching which are voluntary, capacity-limited, and require attention (Neely, 1977). Researchers manipulate the stimulus onset asynchrony (SOA) with the purpose of investigating these processes. The SOA has been identified as a time interval between the appearance of the first stimulus (i.e., prime) and the second one (i.e., target). It is considered that a long SOA allows time for strategic processing, whereas a short SOA (i.e., shorter than 400 ms) allows automatic processing to dominate the priming effects (Hill et al., 2005; Steinhauer et al., 2017). Long SOA allows participants to process the prime in a controlled way, related to attention and strategy. For example, participants may consciously use the prime to generate a list of candidate targets related to the prime, since they have enough time. If the actual target is a member of candidate targets, word identification is facilitated. There is general agreement on the fact that both controlled processes and automatic activation can elicit the N400 priming effect in adults (Anderson & Holcomb, 1995; Hill et al., 2002, 2005; Rossell et al., 2003).
Lexical-semantic system can be based on taxonomic relations or thematic relations. The taxonomic relationship is a classification system that hierarchically contains the category and the members belonging to that category. The organization of concepts in taxonomic relations depends on shared features, functional properties, and similarity distance (Mirman et al., 2017). Taxonomic knowledge is organized among the two dimensions: vertical and horizontal (Markman & Wisniewski, 1997; Rosch, 1978). Subordinate categories (e.g., warship), basic categories (e.g., ship), and superordinate categories (e.g., vehicle), represent the relations of the words in the vertical direction. However, we must mention the horizontal axis where the most typical member of any category is identified. Words such as ‘bus’, ‘truck’, ‘ship’, ‘airplane’ are arranged in horizontal axis below the superordinate category of ‘vehicle’. From this perspective, in terms of the formation of lexical categories and language, taxonomic relations are considered as an important concept (Bermeitinger et al., 2010; Maguire et al., 2010). However, in electrophysiological studies based on paired stimulus paradigms, priming effect on N400 component has been studied by using different subtypes of relations (e.g., restaurant-wine, Chwilla & Kolk 2005; dog-bone-muscle, Deacon et al., 2004; mouse-cheese, Franklin et al., 2007; fruit-apple, Heinze et al., 1998; offspring-grandson, Chen et al., 2014; taxonomically (cow-chicken) and thematically related (egg-chicken), Maguire et al., 2010; donkey-cow, Savic et al., 2017; lamp-mirror, Wamain et al., 2015; Part-whole: finger-hand, Member-category: hammer-tool, Synonyms: far-distant and Antonyms: fast-slow, Steinhauer et al., 2017).
In priming experiments, one important issue to be considered is the processing and activation of thematic and taxonomic representations may differ from each other in the brain. Previous functional neuroimaging studies suggested that thematic and taxonomic relations are stored by distinct neural representations (Kotz et al., 2002; Sachs et al., 2008; Sass et al., 2009; Kumar, 2017). Using event-related potentials (ERP), it was shown that taxonomic mismatch pairs evoked more negativity than thematic ones in the N400 time window (Savic et al., 2017), suggesting more effortless processing of taxonomically-related items. Wamain et al., (2015) reported that thematic priming only affected early ERP components (N1 and P3), indicating earlier activation of thematic relations than taxonomic ones. In an eye-tracking experiment, Kalénine et al., (2012) have reported that the activation time differed across types of relations. This evidence highlights that thematic and taxonomic information vary in terms of temporal dynamics.
Finally, it is crucial to emphasize that the mentioned studies above examined the processing of semantic relations, with various paradigms. For example, Heinze et al., (1998) asked participants to determine if the target word was a member of the superordinate category indicated by the prime or not. Deacon et al., (2004) required participants to decide whether the target word was semantically-related to S1 and /or S2 primes. Maguire et al., (2010) assessed the processing of two types of relations which are taxonomic and thematic by using a passive listening task. In a study by Chen et al., (2014) taxonomic and different subtypes of thematic relations processing was examined in a lexical decision task. Steinhauer et al., (2017) used a semantic judgment task. Savic et al., (2017) focused on a picture-word priming task in which the participants decide whether the image and word matched.
In the current study, semantic priming in the lexical decision task, which may be regarded as a controlled process, was used. As lexical decision task involves different stages of linguistic and cognitive processing, such as perception of visual stimuli, orthographic and phonological analysis of the target word string, detection of this sequence in the mental dictionary, determining whether the target word is in the mental dictionary, accessing the meaning of the target word, this task is believed to evoke lexical search and retrieval processing. However, besides the relationship between prime and target, one potential problem with semantic priming in the lexical decision task is the overlapping of N400, P300, and motor-related potentials (MRP) when participants are instructed to respond quickly to a target stimulus using a button-press. Thus, a delayed lexical decision task is recommended by several studies to prevent overlapping of N400 with MRPs (Hill et al., 2002, 2005; Van Vliet et al., 2014). Therefore, we presented our participants with a delayed lexical decision task in which they had to delay their button-press response until a question mark was presented.
The Late Positive Component (LPC) is another language component, and also termed P600 investigating sentence processing or the P3b component, reflects context-updating. Here, we use the term “LPC” in reference to similar positive components (i.e., P600, P3b). LPC is a positive deflection with a centro-parietal peak at approximately 600 ms. In language studies, the P600/LPC, also known as the ‘Syntactic Positive Shift’, has been linked to a wide range of disagreements in syntactic rules. An increasing number of studies have reported that violations of case, number or gender agreement, phrase structure, or garden path sentences elicited this component (e.g., Coulson et al., 1998; Friederici et al., 1993; Hagoort et al., 1993; Kaan & Swaab 2003; Münte et al., 1997; Neville et al., 1991; Osterhout et al., 1994). Consequently, it was identified as a marker of syntactic repair or reanalysis in previous electrophysiological studies (Friederici, 1995; Hagoort et al., 1999; Kaan & Swaab, 2003). Recent findings have demonstrated that LPC effects, which are termed as “semantic P600”, also appear in sentences with semantic conflict (Hoeks et al., 2004; Kim & Osterhout, 2005; Kuperberg et al., 2003), semantic-thematic violations (Bornkessel-Schlesewsky & Schlesewsky, 2008; Kuperberg, 2007) and semantic reversal anomalies (Kolk et al., 2003; van Herten et al., 2005). In addition, Brouwer et al., (2012) postulated distinct functional roles for LPC and N400 in sentences. LPC has linked to effortful semantic integration into the preceding context, while the N400 is suggested to reflect retrieval of lexical information from memory. But it is important to note that these findings indicate different stages of sentence processing.
Some ERP semantic priming results also reported LPC after the N400 (Bouaffre and Faita-Ainseba 2007; Hill et al., 2002, 2005; Rossell et al., 2003). By using immediate semantic priming task with short and long SOAs, Hill et al., (2002) reported LPC amplitudes decreased with semantic distance in short SOA. Similarly, in Rossell et al.’s (2003) study, increased LPC amplitudes for taxonomically-related target words were detected over the parietal areas only at the short SOA condition. Using a delayed response task, Hill et al., (2005) reported an early (between 430 and 530 ms) LPC evoked by related condition in short SOA and a late LPC (between 700 and 800 ms) evoked by the pseudo-word targets in both SOAs. The authors suggested that the early LPC reflected facilitating the categorization of related-target words consequent to detection of semantic relationships implicitly, while the late one was due to lexical decision. In the study of Bouaffre and Faita-Ainseba (2007), larger LPC amplitudes were recorded in response to thematically-related word pairs at short SOA.
Several studies on different languages have been carried out to investigate language-brain associations in order to reveal whether there is a universal feature accepted for each language. In fact, the N400 priming effect has been demonstrated in several different languages including English (e.g., Anderson & Holcomb 1995; Holcomb & Neville, 1990; Rossell et al., 2003), Spanish (e.g., Silva-Pereyra et al., 1999), German (e.g., Hill et al., 2002; Hill et al., 2005; Weisbrod et al., 1999), Chinese (e.g., Chen et al., 2014; Luo et al., 1999) and Japanese (e.g., Koyama et al., 1992; Matsumoto et al., 2005). To our knowledge, in the current literature there is no data which investigated whether the N400 priming effect also prevails for Turkish language. In order to fill the gap in the literature, the present study investigated semantic priming in Turkish in healthy participants with a delayed lexical decision task via high-temporal resolution recordings of ERPs. Turkish language includes a culture in which oral aspect predominates. However, in cultures where the verbal aspect is dominant, conceptual associations are more concrete and determined by life practices, not by an analytic way. One of our goals was to reveal whether taxonomic relations demonstrate the N400 priming effect in Turkish language too. Secondly, even though LPC may have a potential role in lexico-semantic processing mostly in short SOA, we enquired about the LPC component in the processing of word pairs in Turkish in long SOA.
Materials and Methods
Participants
The participants included in the present study were healthy, native Turkish speakers, aged between 18 and 30 years. The exclusion criteria were as follows: (1) history of psychiatric disorders, (2) Beck Depression Inventory scores ≥ 16 (Beck et al., 1961), (3) Beck Anxiety Inventory scores ≥ 16 (Beck et al., 1988), (4) medications that affect cognition (e.g., anti-psychotics, antidepressants, anxiolytics) (5) history of head trauma, drug or alcohol abuse. A summary of participant general demographic and clinic features is provided in Table 1.
A total of 53 participants were included in cognitive assessment at baseline. 13 participants were excluded at the beginning of the study due to not meeting the inclusion criteria. Electroencephalogram (EEG) was recorded from 40 participants, four of them were excluded by a reason of inadequate epoch numbers (number of artifact-free epochs < 25). The remaining 36 participants were evaluable in the statistical analysis. All participants were confirmed to be right-handed by a Turkish version of Chapman’s Handedness Inventory (Nalçacı et al., 2002) and had an intact vision. They signed informed consent form. Ethical approval for this study was obtained by the Clinical Research Ethics Board of Ankara University, Turkey.
Cognitive Assessment
Neuropsychological measures of the participants were assessed by neuropsychologists. Attention/working memory were evaluated with WMS-R digit span test (Wechsler, 1987); visuospatial abilities using Benton Line Orientation Judgment Test (Benton et al., 1978); language using Boston Naming Test with 31-item (Kaplan et al., 2001); episodic and visual memory with Öktem Verbal Memory Processes Test (OVMPT; Öktem 1992), and WMS-R subtest of visual reproduction (Wecshler, 1987); and executive functions with Wisconsin Card Sorting Test (Heaton et al., 1993), verbal fluency tests (categorical and phonemic), Çapa Version of Stroop Test (Emek-Savaş et al., 2019), WAIS-fourth edition similarities test, Trail Making Test (TMT; Reitan, 1985). All participants performed within local norms.
Stimulus Material
The stimulus consisted of 360 concrete nouns and 120 pseudo-words. All words were nouns and selected from the Word Frequency Dictionary of Written Turkish (Göz, 2003). The stimulus set included 240 prime-target pairs; which were classified as 60 pairs of semantically-related words, 60 pairs of semantically-unrelated and 120 pairs followed by pseudo-words. Words belonging to different superordinate categories such as body parts, animals, clothes, furniture, food, and vehicles were selected. The semantically-related pairs were selected based on semantic fluency and typicality research (Maviş & Tuncer, 2013) in Turkish and they were at the same hierarchical level as category co-members (e.g., apple-pear). The semantic-relatedness was rated by seven speech-language pathologists who were not included in the ERP experiments, by a 5-point Likert scale (A point of 5 grades very strong relatedness). The mean relatedness between the word pairs was 3.8 (SD = 0.41).
The software program used to generate the pseudo-words was “Wuggy” (Keuleers and Brstbaert, 2010). Pseudo-words were phonologically similar to a word, pronounceable, meaningless, legally spelled letter strings. Erten et al., (2014) developed the Turkish version of the Wuggy software. Table 2 demonstrates examples of types of prime-target conditions.
No significant difference was found among mean word frequency of primes (68.66 ± 84.48 for the related prime word, 68.58 ± 84.6 for the unrelated prime words, 68.63 ± 101.09 for the pseudo-word prime words) in all conditions and targets in related and unrelated conditions. Also, the mean word frequency of primes and targets in related and unrelated conditions were not significantly different from each other. All items were three to nine letters in length and contained no more than four syllables and matched for mean word length in both conditions (see Table 3). The experimental stimuli were composed of three blocks in which have 80 trials; 20 related-, 20 unrelated-word pairs, and 40 word pseudo-word pairs. A short break (2–3 min.) was given between blocks.
Design and Procedure
During the EEG recordings, each participant fulfilled a lexical decision task, which was comprised of 240 trials. The first word was defined as “prime” and the second one was defined as the “target”, in all word pairs which were presented successively to measure semantic priming. Participants were asked to choose if the target was a real word or not. Stimuli were presented visually: bold black letters from the Latin alphabet appeared in the center of a gray background and occupied a vertical visual angle of 1ͦ. Both word and pseudo-word (prime and target) were shown only once.
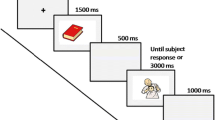
Each trial defined with the following sequence of events. A plus sign as a fixation point was displayed in the center of the monitor for 1000 ms, and then a gray screen was displayed for 500 ms. A real word as a prime was displayed in the center of the monitor for 300 ms. An interstimulus interval for 1700 ms was displayed as a gray screen. A real word or pseudo-word as a target was displayed in the center of the monitor during 1000 ms. A gray screen was displayed for 1000 ms with the purpose of delayed response. The participants are required to give an answer during the question mark displayed in the center of the monitor for a maximum of 3 s. A fixed inter-trial interval was set up to 500 ms. Total trial time was ≤ 9 sec. [5–9 s range]. Figure 1 demonstrates the experimental procedure.

Illustration of the experimental design
Participants were required to delay their button-press response until the question mark was presented. Furthermore, participants were instructed to press the left or right mouse button respectively to classify the target word as a real word or a pseudo word as accurate as possible within a 3000 ms response window. Participants were instructed to favor accuracy over speed. Stimuli presentation and accuracy of responses were controlled by MATLAB (MATLAB 2010a, The MathWorks, Natick) program.
Electrophysiological Recording
EEGs were recorded in a shielded room isolated from electrical and magnetic fields. A standard 22-inch Philips LCD monitor with a response time of 5 ms was used for the stimuli presentation. The distance between the participant’s location and the monitor was 120 cm. EEG signals were recorded with 30 Ag/AgCl surface electrodes placed on an elastic cap (EasyCap; Brain Products GmbH; Gliching, Germany) according to international 10–20 system. The BrainAmp 32-channel DC system amplified the signals with a high-frequency cut-off of 250 Hz, and a sampling rate of 500 Hz. As references linked earlobe electrodes (A1 + A2) were used and the ground electrode was placed at the right earlobe. Horizontal and vertical electrooculograms (EOG) were placed on the outer canthus of the right eye and the supraorbital region, respectively. For all recordings, electrode impedances were kept 10 kΩ.
ERP Processing
EEG signals were analyzed through the use of the Brain Vision Analyzer 2.1 (Brain Products GmbH; Gilching, Germany) software. For each participant, raw data were band-pass filtered at 0.5–30 Hz (slope: 24 dB/octave) and extended-infomax Independent Component Analysis (ICA) decompositions were performed on 28 channels (except reference and EOG channels) in semiautomatic mode to identify and remove ocular artifacts. Data were then epoched from − 200 ms to 1000 ms relative to the onset of the target words and baseline corrected from − 200 ms to 0 ms. The remaining artifacts (muscle, electrical) defined with the following criteria: (1) amplitude changes exceeding ± 50 µV/ms in an epoch; (2) voltage step was more than 50 µV /ms; (3) minimum allowed activity was 0.5 µV in a 100 ms interval and (4) in a 200 ms interval maximum value differences of more than 100 µV. The epoch numbers were equalized randomly between related, unrelated, and pseudo-word stimulation conditions. Only trials from participants who had a minimum of 25 correct artifact-free responses were averaged for each condition and data were analyzed across conditions. After the artifact rejection, the mean number of trials was 41.56 ± 5.98 for related, 41.36 ± 6.03 for unrelated, 41.5 ± 5.96 for pseudo-word conditions [F(2,105) = 0.01, p = 0.99].
For each experimental condition, the mean amplitude values were computed separately to identify the averaged ERPs elicited by target words. Based on observation of time-locked ERPs in data set as well as on previous findings (e.g., Bermeitinger et al., 2010; Hoshino & Thierry, 2012; Rossell et al., 2003), we analyzed the N400 between 300 ms and 500 ms, and the later positive ERP component-LPC (P600) in a time window of 500 ms and 650 ms. The N400 and LPC analysis included nine centro-parietal (C3, Cz, C4, CP3, CPz, CP4, P3, Pz, P4) electrode sites, since these components have a centro-parietal distribution as reported by the literature (for a review, see; Kuperberg 2007; Kutas & Federmeier, 2011 and references therein).
Statistical analysis: Statistical significance was analyzed through the use of the SPSS statistical package (IBM SPSS Statistics, version 22). The percentage of correct responses were compared using repeated-measures ANOVA with condition (3 levels: related, unrelated, pseudo) as a within-subjects factor. The EEG data for N400 and LPC components were analyzed separately with repeated measures ANOVA, which included the within-subject factors condition (3 levels: related, unrelated, pseudo-word), laterality (3 levels: right, midline, left) and anterior-posterior location (3 levels: central, centro-parietal, parietal). Greenhouse-Geisser’s corrections were used for nonsphericity when appropriate. Relevant post-hoc comparisons were computed with paired sample t-test. Bonferroni correction was employed for all multiple comparisons. Results were considered statistically significant at the ≤ 0.05 level (two-tailed).
Results
Behavioral Results
Reaction time (RT) was calculated from the presentation of the question mark until the participants made a response. Participant’s mean reaction times with a delay of 2 s from stimulus onset and the percentage of correct responses were calculated in each condition. As participants had to respond to the question mark probe (which was at a substantial delay of 2 s after target word onset), these data were reported only briefly. Mean RTs were 759 ms. Regarding the correct-response percentages, a repeated-measures ANOVA revealed no significant main effect of correct-response percentages [F(2,70) = 2.736, p = 0.08, ηp2 = 0.073] (see Table 4).
Event-related Potentials
Observation of the grand averaged ERPs time-locked to target word-onset showed characteristic waveforms including earlier visual responses (P1, N1), the N400 potential, and a late positive component (i.e., LPC) with scalp topographies typical for these components. Figure 2 presents elicited ERP waveforms by related, unrelated, and pseudo-word targets in the central and parietal channels.

Grand-average ERPs (N = 36 participants) evoked by semantically-related, unrelated, and pseudo-word targets of the semantic priming task at left, midline, and right central, centro-parietal, and parietal sites. The light gray hue and dark gray hue regions correspond to time windows of N400 and LPC, respectively
Mean N400 Amplitudes
A main effect for CONDITION [F(2,70) = 18.93, p < 0.001, ηp2 = 0.35], presenting that mean N400 amplitudes were higher for pseudo-words and semantically-unrelated targets compared to semantically-related ones (p < 0.001, p = 0.007, respectively). Moreover, N400 amplitudes were higher for pseudo-words than semantically-unrelated targets (p = 0.009) (as displayed in Fig. 3a).

Illustration of the mean N400 (300–500 ms) and LPC (500–650 ms) amplitudes for all experimental conditions. As described in Fig. 3a, the pseudo-words and unrelated-words elicited higher N400 amplitudes in comparison to related-words. N400 amplitudes of pseudo-words were higher than unrelated-words. As highlighted in Fig. 3b the amplitude of LPC was significantly increased for related word pairs. Error bars show standard errors. *p < 0.05, **p < 0.001
A main effect for LOCATION was detected [F(2,70) = 26, p < 0.001, ηp2 = 0.43], indicating that mean N400 amplitudes at central (-0.96 µV, SE = 0.36) and centro-parietal (-0.46 µV, SE = 0.34) sites were more negative than parietal (0.2 µV, SE = 0.36) sites (for all, p < 0.001). Moreover, mean N400 amplitudes at central sites were more negative than centro-parietal (p < 0.001).
A main effect for LATERALITY was found [F(2, 70) = 26.211, p < 0.001, ηp2 = 0.428], indicating that the N400 amplitudes at middle sites (-1.01 µV, SE = 0.4) were more negative than at the left (-0.21 µV, SE = 0.33) and right sites (-0.005 µV, SE = 0.31) (for all, p < 0.001), but there were no significant differences between left and right sites (p = 0.55).
There were no significant interactions between CONDITION x LATERALITY [F(4,140) = 2.031, p = 0.11, ηp2 = 0.055], CONDITION x LOCATION [F(4, 140) = 1.155, p = 0.32, ηp2 = 0.032], or CONDITION x LOCATION x LATERALITY [F(8, 280) = 1.99, p = 0.85, ηp2 = 0.054].
LPC Mean Amplitudes
A main effect for CONDITION [F(2,70) = 9.015, p = 0.001, ηp2 = 0.205] presenting that mean LPC amplitudes were higher for semantically-related words than semantically-unrelated and pseudo-words (p = 0.025, p = 0.002, respectively). No significant difference was obtained for LPC amplitudes between unrelated-words and pseudo-words (p = 0.13) (see Fig. 3b).
A main effect for LOCATION was found [F(2,70) = 22.72, p < 0.001, ηp2 = 0.394], indicating that mean LPC amplitudes at parietal sites (2.4 µV, SE = 0.36) were higher than central (1.53 µV, SE = 0.33) and centro-parietal (2.05 µV, SE = 0.35) sites (p < 0.001, p = 0.002, respectively). Furthermore, higher LPC amplitudes were found at centro-parietal electrode locations compared to central ones (p < 0.001).
The interaction effect of CONDITION x LOCATION x LATERALITY was also significant [F(8,280) = 3.14, p = 0.011, ηp2 = 0.082]. Paired sample t-tests indicated that mean LPC amplitudes elicited by related-words were higher at central, centro-parietal, and parietal locations than pseudo-words (for all; p < 0.006). Increased LPC amplitudes for related-words were only detected at the left centro-parietal (CP3) electrode location compared to semantically-unrelated words (p = 0.004).
The interaction effect of CONDITION x LOCATION [F(4,140) = 2.99, p = 0.06, ηp2 = 0.079] did not reach statistical significance. Neither main effect for LATERALITY [F(2,70) = 2.59, p = 0.093, ηp2 = 0.069], nor the interaction effect of CONDITION x LATERALITY [F(4,140) = 2.05, p = 0.11, ηp2 = 0.055] were significant.
Correlations of LPC with Memory and Attention
The relationship of LPC measures with memory and attention domains were investigated exploratively using Pearson’s correlation. To avoid multiple comparisons, LPC mean amplitudes were defined as the average value of three electrodes at parietal region (P3, Pz, P4), between 500 and 650 ms for related conditions. We focused on parietal electrode sites and related condition since participants show the highest amplitudes in those areas and condition.
Composite scores for memory and attention domains were formed by computing average z-scores of tests in same domain. The scores of OVMPT total learning score, immediate recall, free recall were included for episodic memory domain. The scores of, WMS-R digit span forward and TMT part A were included for attention domain. There were 3 missing values for TMT part A. Only significant correlations that were controlled by Bonferroni correction were presented.
Attention scores and mean LPC amplitudes of related words in parietal region showed moderate correlation (r = 0.407, p = 0.019) (see Fig. 4).

Correlation plot between attention domain and LPC amplitudes at related condition
Discussion
This study used ERPs as a brain-based approach to examine semantic priming in Turkish. In order to investigate the effects of lexical retrieval, participants completed visual semantic priming in a delayed lexical decision task. Semantically-related, unrelated, and pseudo-word conditions were included in the study. The accuracy rate for each target type was very high indicating that participants performed the experiment successfully. Besides, the high correct-response percentages were indicative of greater attentiveness to the experiment. Regarding the ERP data, there were two main findings in the present study. First, N400 peaking around 400 ms was obtained for all three conditions and was the largest for pseudo-words, intermediate for unrelated targets, and the smallest for related targets in the 300–500 ms time window, not surprisingly. Secondly, LPC amplitudes also differed significantly among conditions. The LPC amplitudes for semantically-related words were higher than semantically-unrelated and pseudo-words. Unrelated-word pairs did not differ in amplitude from pseudo-words.
The amplitude of N400 plays a role in retrieving semantic word information from long-term memory (Kutas & Federmeier, 2000); in other words, N400 is an indicator of an activation degree of the words’ in semantic long term memory (Van Petten & Kutas, 1991). The larger and more negative N400 amplitudes are associated with increased efforts of semantic processing (Kutas & Federmeier, 2011). According to Holcomb & Neville (1990), more resources are needed to identify the target when the target is preceded by an unrelated prime. The authors, therefore, presumed the larger N400 for unrelated-word targets is due to the recruitment of these extra resources. The evidence from neuroimaging studies confirmed electrophysiological results by reporting broader activations in the unrelated condition in contrast to the related condition (for a review, see; Holderbaum et al., 2019). However, when the prime-target word pairs are related, lexical activation is facilitated. This may lead to a less ‘effortful’ search in the semantic memory, yielding smaller-amplitude N400s in the related condition. Therefore, significantly larger N400 amplitudes to words preceded by unrelated-words than to words preceded by a semantic associate in our study demonstrate that N400 is sensitive to semantic processing load.
The literature regarding examining word processing by ERPs acknowledged that the N400 waveform was larger to meaningless pseudo-words than to words (Anderson & Holcomb, 1995; Bentin et al., 1985; Chwilla et al., 1995; Holcomb & Neville, 1990; Ziegler et al., 1997; Wang et al., 2017), but notably, not by illegal strings (Coch & Holcomb, 2003; Federmeier & Laszlo, 2009). Evidence of larger N400 to pseudo-words in these studies suggests that instead of lexical access, the N400 reflects accessing of semantic memory. The pseudo-words in the present study were pronounceable non-words that obey the orthographic and phonologic rules of Turkish but carry no meaning. Our ERP findings -more negative deflection for pseudo-words in contrast to related and unrelated conditions- are line with those of Federmeier & Laszlo (2009), Lau et al., (2008), and Pulvermüller et al., (2009). In conclusion, more negative deflection in pseudo-word condition reflects increased working memory load and effort/difficulty in accessing memory representations for these meaningless words.
In the present study, N400 amplitudes were the largest (i.e., most negative) in central sites. Previous studies showed that scalp distributions of N400 vary according to the stimulus type, mainly centro-parietal for sentences (Kutas & Hillyard, 1983; Kutas et al., 1988), central and midline regions for isolated words (Bermeitinger et al., 2010; Landi & Perfetti, 2007) with a right hemisphere for visual (not auditory) words, and bi-hemispheric distribution for visual non-linguistic stimuli (e.g., images) (Khachatryan et al., 2016). As reported by Halgren et al., (2002), N400 may arise in different regions with different stimuli and tasks. Results of a series of neuroimaging studies reported increased activation for sentences including semantic anomalies in various cortical regions like left inferior frontal gyrus, insula, bilateral precentral and middle frontal gyri; temporal pole, fusiform gyrus, superior temporal gyri and angular gyrus, indicating difficulty in integrating anomalous words into semantic context (e.g., Friederici et al., 2003; Hagoort et al., 2004; Kiehl et al., 2002; Luke et al., 2002; Newman et al., 2003; Ni et al., 2000). Moreover, event-related fMRI studies showed that the lexical decision task was associated with larger hemodynamic signals in the unrelated condition in superior/middle temporal gyrus, anterior cingulate, and left inferior frontal gyrus compared with semantically-related ones (e.g., Copland et al., 2003; Kircher et al., 2009; Matsumoto et al., 2005; Ruff et al., 2008; Wible et al., 2006). Considering this literature, it may seem that sentence processing could be related to broader cortical networks, whereas word processing may be associated with more limited brain areas and circuits. The anatomical regions which have been mentioned above are also known as N400 generators based on evidence from event-related optical signal, intracranial and magnetoencephalogram recordings (e.g., Guillem et al., 1995; Halgren et al., 1994; Halgren et al., 2002; Simos et al., 1997; Tse et al., 2007; for a review, see, Lau et al., 2008). This concurs well with the assumption of this distributed network of brain areas involved in semantic memory storage and processes (Kutas & Federmeier, 2011).
Until now, the N400 priming effect has been demonstrated in several different languages. As far as we know, the present ERP findings represent the first published study showing the expected gradient of larger N400 to pseudo-words than unrelated words than related words using Turkish words. It also extends the published literature by replicating the N400 semantic priming effect during a lexical decision task with very long SOAs and a delayed response. The present study adds to the ERP literature by showing how remarkably universal the N400 brain potential is, with similar effects across languages and orthography, whether using letters or pictorial characters (Koyama et al., 1992; Luo et al., 1999; Matsumoto et al., 2005; Chen et al., 2014) and also elicited by images of objects (Ganis et al., 1996) or spoken language (Holcomb & Neville, 1990). This supports a view of the distributed semantic network being remarkably similar across cultures and human populations. Our paradigm used a long SOA, which had a major effect on the behavioral /response time (RT) results, but which shows that motor response processes (commonly overlapping in time with the N400 more than the LPC) have minimal effects on these specific brain potentials.
The other main finding of current study is related to the LPC potential. The LPC has identified as a member of a family of late positivity, including the P300 (Donchin, 1981), which is larger for rare and unexpected stimuli. In the present study, LPC amplitudes were enhanced as the degree of semantic association between prime and target words increased. However, our paradigm was designed to minimize P300 effects by using equal probabilities of related and unrelated words and also by using each Turkish word only once reducing possible ‘target’ effects and high probability occurrences.
LPC has also been associated with memory and recognition processes, especially in old/new experiments. Repeated (old) words elicit greater positivity than new words (for a review see Rugg & Allan 2000) and the increased amplitude in new words is associated with episodic verbal memory capacity (Olichney et al., 2000, 2002, 2010). Each target was shown only once in the present study, thus the modulation of LPC amplitude may not have resulted from the old/new effect.
Results of our study clearly showed that LPC was modulated with semantic relatedness of words as compared to the unrelated- or pseudo-words. LPC priming effects may reflect post-lexical relational processing between prime and target. Moreover, our results indicate a moderate association between LPC amplitudes and attention scores. Some authors argue that LPC is associated with general attentional processes (Coulson & Kutas, 2001), and also may be an index of the “relationship-detection process” (Bouaffre and Faita-Ainseba, 2007). Given that long SOA is used, participants are likely to confirm a semantic relationship between prime and target words, after introducing the target. As the semantically-related words shared category membership, matching the semantic relatedness between the prime-target compounds in attention may have yielded more positive LPC amplitudes for semantically-related words in contrast to unrelated and pseudo-words.
In the current study, we have once more showed that LPC was not only triggered by syntactic processing, but also may occur in the processing of Turkish word pairs. As it was also stated in the introduction, the LPC amplitudes increase in short SOA conditions in taxonomically related words (Hill et al., 2002, 2005; Rossel et al., 2003), and in thematically related word pairs (Bouaffre and Faita-Ainseba, 2007). We believe that this is the first study demonstrating a LPC effect in response to taxonomically-related words at long SOA in a lexical decision task.
Conclusions
Findings of this study emphasize the differential processing of related-, unrelated- and pseudo-word targets as measured by N400 and LPC. Our work has led us to the conclusion that N400 was associated with a reflection of lexical-semantic retrieval, and LPC may reflect retrieval of strategic information during semantic tasks such as conscious recognition of prime-target relationships. Moreover, both N400 and LPC amplitudes were found to be modulated by semantically-related word pairs. The novelty of our work was the use of a delayed semantic priming paradigm with only taxonomic associations by means of ERPs. In addition, semantic priming and lexical decision paradigm has been designed to minimize methodological problems regarding the superposition of P300 and N400, as well as the involvement of MRP in processing. This approach can overcome the limitations of the existing literature in the context of understanding brain dynamics that play a role in the processing of mental representations of the relationship between word meanings and written forms of words. Taken together, the findings of our study add to a growing body of literature on the understanding semantic processing of taxonomic relations. Future studies should aim to compare the effects of short and long SOAs, as well as taxonomic and thematically-related word pairs at different stages of language development or in individuals with language disorders.
Data Availability
The data from this study are available from the corresponding author upon reasonable request.
References
Anderson, J. E., & Holcomb, P. J. (1995). Auditory and visual semantic priming using different stimulus onset asynchronies: an event-related brain potential study. Psychophysiology,32(2),177–190. https://doi.org/10.1111/j.1469-8986.1995.tb03310.x
Beck, A. T., Epstein, N., Brown, G., & Steer, R. A. (1988). An inventory for measuring clinical anxiety: Psychometric properties. Journal of Consulting and Clinical Psychology, 56(6), 893–897. https://doi.org/10.1037/0022-006X.56.6.893
Beck, A. T., Ward, C. H., Mendelson, M., Mock, J., & Erbaugh, J. (1961). An inventory for measuring depression. Archives of General Psychiatry, 4, 561–571. https://doi.org/10.1001/archpsyc.1961.01710120031004
Bentin, S. (1987). Event-related potentials, semantic processes, and expectancy factors in word recognition. Brain and Language, 31, 308–327. https://doi.org/10.1016/0093-934X(87)90077-0
Bentin, S., McCarthy, G., & Wood, C. C. (1985). Event-related potentials, lexical decision and semantic priming. Electroencephalography and Clinical Neurophysiology, 60(4), 343–355. https://doi.org/10.1016/0013-4694(85)90008-2
Benton, A. L., Varney, N. R., & Hamsher, K. D. (1978). Visuospatial judgment. A clinical test. Arch Neurol., 35(6), 364–367. https://doi.10.1001/archneur.1978.00500300038006
Bermeitinger, C., Frings, C., & Wentura, D. (2010). N400 effects for category exemplars primed by category labels. Scientific Report, PsyDok,1–10
Bonin, P. (2004). Mental lexicon: Some words to talk about words. New York: Nova Science Publishers
Bornkessel-Schlesewsky, I., & Schlesewsky, M. (2008). An alternative perspective on “Semantic P600” effects in language comprehension. Brain Research Reviews, 59(1), 55–73. https://doi.org/10.1016/j.brainresrev.2008.05.003
Bouaffre, S., & Faïta-Ainseba, F. (2007). Hemispheric differences in the time-course of semantic priming processes: evidence from event-related potentials (ERPs). Brain And Cognition, 63(2), 123–135. https://doi.org/10.1016/j.bandc.2006.10.006
Brouwer, H., Fitz, H., & Hoeks, J. (2012). Getting real about semantic illusions: Rethinking the functional role of the P600 in language comprehension. Brain Research, 1446, 127–143. https://doi.10.1016/j.brainres.2012.01.055
Brown, C. M., Hagoort, P., & Chwilla, D. J. (2000). An event-related brain potential analysis of visual word priming effects. Brain and Language, 72(2), 158–190. https://doi.org/10.1006/brln.1999.2284
Chen, Q., Ye, C., Liang, X., Cao, B., Lei, Y., & Li, H. (2014). Automatic processing of taxonomic and thematic relations in semantic priming - Differentiation by early N400 and late frontal negativity. Neuropsychologia, 64, 54–62. https://doi.org/10.1016/j.neuropsychologia.2014.09.013
Chwilla, D. J., & Kolk, H. H. J. (2005). Accessing world knowledge: Evidence from N400 and reaction time priming. Cognitive Brain Research, 25(3), 589–606. https://doi.10.1016/j.cogbrainres.2005.08.011
Chwilla, D. J., Brown, C. M., & Hagoort, P. (1995). The N400 as a function of the level of processing. Psychophysiology, 32(3), 274–285. https://doi.10.1111/j.1469-8986.1995.tb02956.x
Coch, D., & Holcomb, P. J. (2003). The N400 in beginning readers. Developmental Psychobiology, 43(2), 146–166. https://doi.org/10.1002/dev.10129
Copland, D. A., de Zubicaray, G. I., McMahon, K., Wilson, S. J., Eastburn, M., & Chenery, H. J. (2003). Brain activity during automatic semantic priming revealed by event-related functional magnetic resonance imaging. NeuroImage, 20(1), 302–310. https://doi.10.1016/s1053-8119(03)00279-9
Coulson, S., King, J. W., & Kutas, M. (1998). Expect the unexpected: Event-related brain response to morphosyntactic violations. Language and Cognitive Processes, 13(1), 21–58. https://doi.org/10.1080/016909698386582
Coulson, S., & Kutas, M. (2001). Getting it: human event-related brain response to jokes in good and poor comprehenders. Neuroscience Letters, 316(2), 71–74. https://doi.org/10.1016/S0304-3940(01)02387-4
Deacon, D., Grose-Fifer, J., Yang, C. M., Stanick, V., Hewitt, S., & Dynowska, A. (2004). Evidence for a New Conceptualization of Semantic Representation in the Left and Right Cerebral Hemispheres. Cortex; A Journal Devoted To The Study Of The Nervous System And Behavior, 40(3), 467–478. https://doi.org/10.1016/S0010-9452(08)70140-0
Donchin, E. (1981). Surprise?… Surprise! Psychophysiology, 18(5), 493–513. https://doi.org/10.1111/j.1469-8986.1981.tb01815.x
Emek-Savaş, D. D., Yerlikaya, D., Yener, G. G., & Öktem, Ö. (2019). Validity, reliability and norm scores of the Stroop test Çapa version. Turkish Journal of Psychiatry. In press, https://doi.org/10.5080/u23549
Erten, B., Bozsahin, C., & Zeyrek, D. (2014, May). Turkish Resources for Visual Word Recognition. In LREC (pp. 2106–2110)
Federmeier, K. D., & Laszlo, S. (2009). Time for meaning: Electrophysiology provides insights into the dynamics of representation and processing in semantic memory. Psychology of learning and motivation, 51, 1–44. https://doi.org/10.1016/S0079-7421(09)51001-8
Franklin, M. S., Dien, J., Neely, J. H., Huber, E., & Waterson, L. D. (2007). Semantic priming modulates the N400, N300, and N400RP. Clinical Neurophysiology, 118(5), 1053–1068. https://doi.10.1016/j.clinph.2007.01.012
Friederici, A. D., Pfeifer, E., & Hahne, A. (1993). Event-related brain potentials during natural speech processing: Effects of semantic, morphological and syntactic violations. Brain Research: Cognitive Brain Research, 1(3), 183–192. https://doi.org/10.1016/0926-6410(93)90026-2
Friederici, A. D., Rueschemeyer, S. A., Hahne, A., & Fiebach, C. (2003). The role of left inferior frontal and superior temporal cortex in sentence comprehension: localizing syntactic and semantic processes. Cerebral Cortex, 13(2), 170–177. https://doi.org/10.1093/cercor/13.2.170
Friederici, A. D. (1995). The time course of syntactic activation during language processing: a modelbased on neurological and neurophysiological data. Brain and Language, 50(3), 259–281. https://doi.org/10.1006/brln.1995.1048
Ganis, G., Kutas, M., & Sereno, M. I. (1996). The search for “common sense”: An electrophysiological study of the comprehension of words and pictures in reading. Journal Of Cognitive Neuroscience, 8, 89–106. https://doi.org/10.1162/jocn.1996.8.2.89
Göz, İ. (2003). Yazılı Türkçenin Kelime Sıklığı Sözlüğü. Ankara: Türk Dil Kurumu Yayınları
Guillem, F., N’kaoua, B., Rougier, A., & Claverie, B. (1995). Intracranial topography of event-related potentials (N400/P600) elicited during a continuous recognition memory task. Psychophysiology, 32(4), 382–392. https://doi.org/10.1111/j.1469-8986.1995.tb01221.x
Hagoort, P., Brown, C. M., & Groothusen, J. (1993). The syntactic positive shift as an ERP measure of syntactic processing. Language and Cognitive Processes, 8(4), 439–483. https://doi.org/10.1080/01690969308407585
Hagoort, P., Brown, C., & Osterhout, L. (1999). The neurocognition of syntactic processing. In C. Brown, & P. Hagoort (Eds.), Neurocognition of Language (pp. 273–316). Oxford, UK: Oxford University Press
Hagoort, P., Hald, L., Bastiaansen, M., & Petersson, K. M. (2004). Integration of word meaning and world knowledge in language comprehension. Science, 304, 438–441. http://doi.10.1126/science.1095455
Halgren, E., Baudena, P., Heit, G., Clarke, J. M., & Marinkovic, K. (1994). Spatio-temporal stages in face and word processing. 1. Depth-recorded potentials in the human occipital, temporal and parietal lobes. Journal of Physiology-Paris, 88(1), 1–50. https://doi.org/10.1016/0928-4257(94)90092-2
Halgren, E., Dhond, R. P., Christensen, N., Van Petten, C., Marinkovic, K., Lewine, J. D., & Dale, A. M. (2002). N400-like magnetoencephalography responses modulated by semantic context, word frequency, and lexical class in sentences. Neuroimage, 17(3), 1101–1116. https://doi.org/10.1006/nimg.2002.1268
Heaton, R. K., Chelune, G. J., Talley, J. L., Kay, G. G., & Curtiss, G. (1993). Wisconsin Card Sorting Test Manual: Revised and Expanded. Odessa, FL: Psychological Assessment Resources
Heinze, H. J., Muente, T. F., & Kutas, M. (1998). Context effects in a category verification task as assessed by event-related brain potential (ERP) measures. Biological Psychology, 47(2), 121–135. https://doi.org/10.1016/S0301-0511(97)00024-0
Hill, H., Ott, F., & Weisbrod, M. (2005). SOA-dependent N400 and P300 semantic priming effects using pseudoword primes and a delayed lexical decision. International Journal of Psychophysiology, 56(3), 209–221. https://doi.org/10.1016/j.ijpsycho.2004.12.004
Hill, H., Strube, M., Roesch-Ely, D., & Weisbrod, M. (2002). Automatic vs. controlled processes in semantic priming - Differentiation by event-related potentials. International Journal of Psychophysiology, 44(3), 197–218. https://doi.org/10.1016/S0167-8760(01)00202-1
Hoeks, J. C., Stowe, L. A., & Doedens, G. (2004). Seeing words in context: the interaction of lexical and sentence level information during reading. Cognitive Brain Research, 19(1), 59–73.https://doi.org/10.1016/j.cogbrainres.2003.10.022
Holcomb, P. J., & Neville, H. J. (1990). Auditory and visual semantic priming in lexical decision: A comparison using event-related brain potentials. Language and Cognitive Processes, 5(4), 281–312. https://doi.org/10.1080/01690969008407065
Holcomb, P. J. (1988). Automatic and attentional processing: An event-related brain potential analysis of semantic priming. Brain and Language, 35(1), 66–85. https://doi.org/10.1016/0093-934X(88)90101-0
Holderbaum, C. S., Sbicigo, J. B., Buchweitz, A., & de Salles, J. F. (2019). A systematic review of brain imaging studies of semantic priming. Interação Em Psicologia (online). https://doi.org/10.5380/psi.v23i02.58553
Hoshino, N., & Thierry, G. (2012). Do Spanish-English bilinguals have their fingers in two pies - or is it their toes? An electrophysiological investigation of semantic access in bilinguals. Frontiers in Psychology, 3, 9. https://doi.org/10.3389/fpsyg.2012.00009
Jones, L. L., & Estes, Z. (2012). Lexical priming: Associative, semantic, and thematic influences on word recognition. In J. S. Adelman (Ed.), Word Recognition (Vol. 2): Meaning and context, individuals and development (pp. 44–72). Hove, UK: Psychology Press
Kaan, E., & Swaab, T. Y. (2003). Repair, revision, and complexity in syntactic analysis: an electrophysiological differentiation. Journal Cognitive Neuroscience, 15, 98–110. https://doi.org/10.1162/089892903321107855
Kalénine, S., Mirman, D., Middleton, E. L., & Buxbaum, L. J. (2012). Temporal dynamics of activation of thematic and functional knowledge during conceptual processing of manipulable artifacts. Journal of Experimental Psychology: Learning Memory and Cognition, 38(5), 1274–1295. https://doi.org/10.1037/a0027626
Kaplan, E. F., Goodglass, H., & Weintraub, S. (2001). The Boston naming test (2nd ed.). Philadelphia: Lippincott Williams & Wilkins
Khachatryan, E., Chumerin, N., Carrette, E., Camarrone, F., De Taeye, L., Meurs, A. … Van Hulle, M. M. (2016, June). Cortical distribution of N400 potential in response to semantic priming with visual non-linguistic stimuli. In 2016 IEEE Statistical Signal Processing Workshop (SSP) (pp. 1–5). IEEE. https://doi.org/10.1109/SSP.2016.7551795
Keuleers, E., & Brysbaert, M. (2010). Wuggy: A multilingual pseudoword generator. Behavior Research Methods, 42(3), 627–633. https://doi.org/10.3758/BRM.42.3.627
Kiehl, K. A., Laurens, K., & Liddle, P. F. (2002). Reading anomalous sentences: an event-related fMRI study of semantic processing. Neuroimage, 17(2), 842–850
Kim, A., & Osterhout, L. (2005). The independence of combinatory semantic processing: evidence from event-related potentials. Journal of Memory and Language, 52(2),205 – 25. https://doi.10.1016/j.jml.2004.10.002
Kircher, T., Sass, K., Sachs, O., & Krac, S. (2009). Priming words with pictures: neural correlates of semantic associations in a cross-modal priming task using fMRI. Human Brain Mapping, 30(12), 4116–4128. https://doi.org/10.1002/hbm.20833
Kolk, H. H., Chwilla, D. J., Van Herten, M., & Oor, P. J. (2003). Structure and limited capacity in verbal working memory: A study with event-related potentials. Brain and Language, 85(1), 1–36. https://doi.org/10.1016/S0093-934X(02)00548-5
Kotz, S. A., Cappa, S. F., von Cramon, D. Y., & Friederici, A. D. (2002). Modulation of the lexical-semantic network by auditory semantic priming: An event-related functional MRI study. Neuroimage, 17(4), 1761–1772. https://doi.org/10.1006/nimg.2002.1316
Koyama, S., Nageishi, Y., & Shimokochi, M. (1992). Effects of semantic context and event-related potentials: N400 correlates with inhibition effect. Brain and Language, 43(4), 668–681. https://doi.org/10.1016/0093-934X(92)90089-W
Kumar, U. (2017). The neural realm of taxonomic and thematic relation: an fMRI study. Language Cognition and Neuroscience, 33(5), 648–658. https://doi.org/10.1080/23273798.2017.1411962
Kuperberg, G. R., Sitnikova, T., Caplan, D., & Holcomb, P. J. (2003). Electrophysiological distinctions in processing conceptual relationships within simple sentences. Cognitive Brain Research, 17(1), 117–129. https://doi.org/10.1016/S0926-6410(03)00086-7
Kuperberg, G. R. (2007). Neural mechanisms of language comprehension: Challenges to syntax. Brain Research, 1146, 23–49. https://doi.org/10.1016/j.brainres.2006.12.063
Kutas, M., & Federmeier, K. D. (2000). Electrophysiology reveals semantic memory use in language comprehension. Trends in Cognitive Sciences, 4(12), 463–470. https://doi.org/10.1016/S1364-6613(00)01560-6
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting: Finding meaning in the N400 component of the event related brain potential (ERP). Annu Rev Psychol, 62, 621–647. https://doi.org/10.1146/annurev.psych.093008.131123
Kutas, M., & Hillyard, S. A. (1983). Event-related brain potentials to grammatical errors and semantic anomalies. Memory and Cognition, 11(5), 539–550. https://doi.org/10.3758/BF03196991
Kutas, M., Van Petten, C., & Besson, M. (1988). Event-related potential asymmetries during the reading of sentences. Electroencephalography and Clinical Neurophysiology, 69(3), 218–233. https://doi.10.1016/0013-4694(88)90131-9
Landi, N., & Perfetti, C. A. (2007). An electrophysiological investigation of semantic and phonological processing in skilled and less-skilled comprehenders. Brain and Language, 102(1), 30–45. https://doi.org/10.1016/j.bandl.2006.11.001
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network for semantics: (De)constructing the N400. Nature Reviews Neuroscience, 9, 920–933. https://doi.org/10.1038/nrn2532
Luke, K. K., Liu, H. L., Wai, Y. Y., Wan, Y. L., & Tan, L. H. (2002). Functional anatomy of syntactic and semantic processing in language comprehension. Human Brain Mapping, 16(3), 133–145. https://doi.org/10.1002/hbm.10029
Luo, Y. J., Hu, S., Weng, X. C., & Wei, J. H. (1999). Effects of semantic discrimination of chinese words on n400 component of event-related potentials. Perceptual and Motor Skills, 89(1), 185–193. https://doi.org/10.2466/pms.1999.89.1.185
Maguire, M. J., Brier, M. R., & Ferree, T. C. (2010). EEG theta and alpha responses reveal qualitative differences in processing taxonomic versus thematic semantic relationships. Brain and Language, 114(1), 16–25. https://doi.org/10.1016/j.bandl.2010.03.005
Markman, A. B., & Wisniewski, E. J. (1997). Similar and different: The differentiation of basic-level categories. Journal of Experimental Psychology: Learning Memory and Cognition, 23(1), 54–70. https://doi.org/10.1037/0278-7393.23.1.54
Matsumoto, A., Iidaka, T., Haneda, K., Okada, T., & Sadato, N. (2005). Linking semantic priming effect in functional MRI and event-related potentials. Neuroimage, 24, 624–634. https://doi.org/10.1016/j.neuroimage.2004.09.008
Maviş, İ., & Tuncer, M. (2013). Semantik Akıcılık. Ankara: Detay Yayıncılık
Mirman, D., Landrigan, J. F., & Britt, A. E. (2017). Taxonomic and thematic semantic systems. Psychological Bulletin, 143(5), 499–520. https://doi.org/10.1037/bul0000092
Münte, T. F., Szentkuti, A., Wieringa, B. M., Matzke, M., & Johannes, S. (1997). Human brain potentials to reading syntactic errors in sentences of different complexity. Neuroscience Letters, 235(3), 105–108. https://doi.org/10.1016/S0304-3940(97)00719-2
Nalçaci, E., Kalaycioğlu, C., Güneş, E., & Ciçek, M. (2002). Reliability and validity of a handedness questionnaire. Turk Psikiyatri Dergisi = Turkish Journal of Psychiatry, 13(2), 99–106
Neely, J. (1991). Semantic priming effects in visual word recognition: a selective review of current findings and theories. In D. Besner, & G. W. Humphreys (Eds.), Basic processes in reading: Visual word recognition (pp. 264–336). Hillsdale: Erlbaum
Neely, J. H. (1977). Semantic priming and retrieval from lexical memory: Roles of inhibition less spreading activation and limited-capacity attention. Journal of Experimental Psychology: General, 106(3), 226–254. https://doi.org/10.1037/0096-3445.106.3.226
Neville, H., Nicol, J. L., Barss, A., Forster, K. I., & Garrett, M. F. (1991). Syntactically based sentence processing classes: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience, 3(2), 151–165. https://doi.org/10.1162/jocn.1991.3.2.151
Newman, S. D., Just, M. A., Keller, T. A., Roth, J., & Carpenter, P. A. (2003). Differential effects of syntactic and semantic processing on the subregions of Broca’s area. Cognitive Brain Research, 16(2), 297–307. https://doi.10.1016/s0926-6410(02)00285-9
Ni, W., Constable, R. T., Mencl, W. E., Pugh, K. R., Fulbright, R. K., Shaywitz, S. E. … Shankweiler, D. (2000). An event-related neuroimaging study distinguishing form and content in sentence processing. Journal of Cognitive Neuroscience, 12(1), 120–133. https://doi.10.1162/08989290051137648
Noldy, N. E., Stelmack, R. M., & Campbell, K. B. (1990). Event related potentials and recognition memory for pictures and words: The effects of intentional and incidental learning. Psychophysiology, 27(4), 417–428. https://doi.org/10.1111/j.1469-8986.1990.tb02337.x
Olichney, J. M., Chan, S., Wong, L. M., Schneider, A., Seritan, A., Niese, A. … Hagerman, R. (2010). Abnormal N400 word repetition effects in fragile X-associated tremor/ataxia syndrome. Brain, 133(5), 1438–1450. https://doi.org/10.1093/brain/awq077
Olichney, J. M., Morris, S. K., Ochoa, C., Salmon, D. P., Thal, L. J., Kutas, M., & Iragui, V. J. (2002). Abnormal verbal event related potentials in mild cognitive impairment and incipient Alzheimer’s disease. Journal of Neurology Neurosurgery and Psychiatry, 73(4), 377–384. https://doi.org/10.1136/jnnp.73.4.377
Olichney, J. M., Van Petten, C., Paller, K. A., Salmon, D. P., Iragui, V. J., & Kutas, M. (2000). Word repetition in amnesia: Electrophysiological measures of impaired and spared memory. Brain, 123(9), 1948–1963. https://doi.org/10.1093/brain/123.9.1948
Ortells, J. J., Vellido, C., Daza, M. T., & Noguera, C. (2006). Semantic priming effects with and without perceptual awareness. Psicologica, 27(2), 225–242
Osterhout, L., & Holcomb, P. J. (1995). Event related potentials and language comprehension. In M. D. Rugg, & M. G. H. Coles (Eds.), Electrophysiology of mind: Event-related brain potentials and cognition (pp. 171–215). New York: Oxford University Press
Osterhout, L., Holcomb, P. J., & Swinney, D. A. (1994). Brain potentials elicited by garden-path sentences: Evidence of the application of verb information during parsing. Journal of Experimental Psychology: Learning Memory and Cognition, 20(4), 786–803. https://doi.org/10.1037/0278-7393.20.4.786
Öktem, Ö. (1992). A verbal test of memory processes. Arch Neuropsychiatry, 29, 196–206
Posner, M. I., & Snyder, C. R. R. (1975). Attention and cognitive control. In R. Solso (Ed.), Information Processing and Cognition (pp. 55–85). Hillsdale, NJ: Erlbaum
Pulvermüller, F., Shtyrov, Y., & Hauk, O. (2009). Understanding in an instant: Neurophysiological evidence for mechanistic language circuits in the brain. Brain and Language, 110(2), 81–94. https://doi.org/10.1016/j.bandl.2008.12.001
Reitan, R. M., & Wolfson, D. (1985). The Halstead-Reitan Neuropsychological Test Battery: Theory and Interpretattion. Tucson, AZ: Neuropsychology Press
Rosch, E. (1978). Principles of categorization. In E. Rosch, & B. Llyod (Eds.), Cognition and categorization (pp. 27–48). Hillsdale, NJ: Erlbaum
Rossell, S. L., Price, C. J., & Nobre, A. C. (2003). The anatomy and time course of semantic priming investigated by fMRI and ERPs. Neuropsychologia, 41(5), 550–564. https://doi.org/10.1016/S0028-3932(02)00181-1
Rugg, M. D. (1984). Event-related potentials and the phonological processing of words and non-words. Neuropsychologia, 22(4), 435–443. https://doi.org/10.1016/0028-3932(84)90038-1
Rugg, M. D., & Allan, K. (2000). Event-related potential studies of memory. The Oxford handbook of memory, 521, 37
Ruff, I., Blumstein, S. E., Myers, E. B., & Hutchison, E. (2008). Recruitment of anterior and posterior structures in lexical–semantic processing: an fMRI study comparing implicit and explicit tasks. Brain and Language, 105(1), 41–49. https://doi.org/10.1016/j.bandl.2008.01.003
Sachs, O., Weis, S., Zellagui, N., Huber, W., Zvyagintsev, M., Mathiak, K., & Kircher, T. (2008). Automatic processing of semantic relations in fMRI: Neural activation during semantic priming of taxonomic and thematic categories. Brain Research, 1218, 194–205. https://doi.org/10.1016/j.brainres.2008.03.045
Sass, K., Sachs, O., Krach, S., & Kircher, T. (2009). Taxonomic and thematic categories: Neural correlates of categorization in an auditory-to-visual priming task using fMRI. Brain Research, 1270, 78–87. https://doi.org/10.1016/j.brainres.2009.03.013
Savic, O., Savic, A. M., & Kovic, V. (2017). Comparing the temporal dynamics of thematic and taxonomic processing using event-related potentials. PLoS ONE, (12)2, e0189362. https://doi.org/10.1371/journal.pone.0189362
Silva-Pereyra, J., Harmony, T., Villanueva, G., Fernández, T., Rodríguez, M., Galán, L., & Reyes, A. (1999). N400 and lexical decisions: Automatic or controlled processing? Clinical Neurophysiology, 110(5), 813–824. https://doi.org/10.1016/S1388-2457(99)00009-7
Simos, P. G., Basile, L. F., & Papanicolaou, A. C. (1997). Source localization of the N400 response in a sentence-reading paradigm using evoked magnetic fields and magnetic resonance imaging. Brain Research, 762(1–2), 29–39. https://doi.10.1016/s0006-8993(97)00349-1
Steinhauer, K., Royle, P., Drury, J. E., & Fromont, L. A. (2017). The priming of priming: Evidence that the N400 reflects context-dependent post-retrieval word integration in working memory. Neuroscience Letters, 651, 192–197. https://doi.org/10.1016/j.neulet.2017.05.007
Tse, C. Y., Lee, C. L., Sullivan, J., Garnsey, S. M., Dell, G. S., Fabiani, M., & Gratton, G. (2007). Imaging cortical dynamics of language processing with the event-related optical signal. Proceedings of the National Academy of Sciences, 104(43), 17157–17162. https://doi.org/10.1073/pnas.0707901104
van Herten, M., Kolk, H. H., & Chwilla, D. J. (2005). An ERP study of P600 effects elicited by semantic anomalies. Cognitive Brain Research, 22(2), 241–255. https://doi.org/10.1016/j.cogbrainres.2004.09.002
Van Petten, C., & Kutas, M. (1991). Influences of semantic and syntactic context on open and closed-class words. Memory & Cognition, 19, 95–112. https://doi.org/10.3758/BF03198500
van Vliet, M., Manyakov, N. V., Storms, G., Fias, W., Wiersema, J. R., & Van Hulle, M. M. (2014). Response-related potentials during semantic priming: The effect of a speeded button response task on ERPs. Plos One, 9(2), e87650. https://doi.org/10.1371/journal.pone.0087650
Wamain, Y., Pluciennicka, E., & Kalénine, S. (2015). A saw is first identified as an object used on wood: ERP evidence for temporal differences between thematic and functional similarity relations. Neuropsychologia, 71, 28–37. https://doi.org/10.1016/j.neuropsychologia.2015.02.034
Wang, E., Zhou, G., Huang, H., Qian, M., Wang, C., & Wang, Y. (2017). N400 and P600 effect of Chinese words recognition. NeuroQuantology, 15(4), 76–83. https://doi.org/10.14704/nq.2017.15.4.1172
Wible, C. G., Han, S. D., Spencer, M. H., Kubicki, M., Niznikiewicz, M. H., Jolesz, F. A. … Nestor, P. (2006). Connectivity among semantic associates: an fMRI study of semantic priming. Brain and Language, 97(3), 294–305. https://doi.org/10.1016/j.bandl.2005.11.006
Wechsler, D. A. (1987). Wechsler memory scale-revised. New York: Psychological Corporation
Weisbrod, M., Kiefer, M., Winkler, S., Maier, S., Hill, H., Roesch-Ely, D., & Spitzer, M. (1999). Electrophysiological correlates of direct versus indirect semantic priming in normal volunteers. Cognitive Brain Research, 8(3), 289–298. https://doi.org/10.1016/S0926-6410(99)00032-4
Ziegler, J. C., Besson, M., Jacobs, A. M., Nazir, T. A., & Carr, T. H. (1997). Word, pseudoword, and nonword processing: A multitask comparison using event-related brain potentials. Journal of Cognitive Neuroscience, 9(6), 758–775. https://doi.10.1162/jocn.1997.9.6.758
Acknowledgements
The authors would like to acknowledge and thank Assoc. Prof. Soner Akşehirli for his comments and valuable suggestions during writing processing of this study.
Funding
This research was supported by Ankara University Scientific Research Projects Coordination Unit with project number 16L02000001.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Disclosure statement
The authors declared that there is no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Düzenli-Öztürk, S., Hünerli-Gündüz, D., Emek-Savaş, D.D. et al. Taxonomically-related Word Pairs Evoke both N400 and LPC at Long SOA in Turkish. J Psycholinguist Res 51, 1431–1451 (2022). https://doi.org/10.1007/s10936-022-09907-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10936-022-09907-2