
Abstract
Organoids are in vitro 3D self-organizing tissues that mimic embryogenesis. Organoid research is advancing at a tremendous pace, since it offers great opportunities for disease modeling, drug development and screening, personalized medicine, as well as understanding organogenesis. Mechanobiology of organoids is an unexplored area, which can shed light to several unexplained aspects of self-organization behavior in organogenesis. It is becoming evident that collective cell behavior is distinctly different from individual cells’ conduct against certain stimulants. Inherently consisting of higher number of degrees of freedom for cell motility and more complex cell-to-cell and cell-to-extracellular matrix behavior, understanding mechanotransduction in organoids is even more challenging compared with cell communities in 2D culture conditions. Yet, deciphering mechanobiology of organoids can help us understand effects of mechanical cues in health and disease, and translate findings of basic research toward clinical diagnosis and therapy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Since ancient times, scientists were observing and studying moving objects, absolute or relative motions of almost everything around them, including the human body. Especially the latest, including human locomotion, growing from embryo to the dying age, assisted by anatomical observations, resulted into a scientific discipline, drawing from various disciplines (as disciplines were separated at later times) named biomechanics. Until the discovery of the cell, as the unit of living bodies, biomechanics had as its studying object the whole body, tissues, and some organs.
With the great strides that biology took in studying the cellular function, the cellular movements (cell division, migration, differentiation) at many levels of organization, including the molecular one, it became evident that the physical environment, notably the mechanical one, had to be considered in detail, as a basic contributing factor to understanding cellular function. In the last 2–3 decades, slowly but steadily, the concept of mechanobiology emerged as a discipline that explores the role of mechanical forces in cellular development, physiology, and disease (Sunyer and Trepat 2017).
Animal models, tissue explants, two-dimensional (2D) cell culture systems, and three-dimensional (3D) tissue-engineered culture systems have been studied in order to mimic the functions of human organs to make diverse number of tests on these models, involving basic tissue morphogenesis, disease modeling and progress, pharmaceutical screening, and even personalized medicine (Jackson and Lu 2016). Although animal models have been successful in mimicking physiology in vivo, anatomical and physiological differences of animals and humans, unknown variables, limited imaging capabilities, and limited animal use due to ethical rules restrict the use of these models (Yin et al. 2016). Even though tissue explants from human beings show the cellular organization and functions of human organs in the best way, these structures cannot be cultured for a long time and they lose the tissue phenotype quickly in vitro (Gähwiler et al. 1997). Researchers have tried to eliminate all these negativities with 2D cell culture systems using human cells for many years. In conventional cell culture studies, cells grow on a homogeneous, stiff surface instead of a heterogeneous and soft extracellular matrix (ECM) as it is in vivo. In 2D cell culture systems, due to the fact that cell-cell and cell-matrix interactions are different from in vivo because of the lack of ECM, as well as spatial organization among the cells, there are greater problems in these systems, such as silencing of some signaling pathways, changing of gene and protein expressions, cellular phenotype, and functions (Kapałczyńska et al. 2018). In order to increase the similarity to the tissues and organs of interest, to mimic in vivo cellular organization and tissue functions, researchers focus on 3D tissue engineering products, where cells can interact more realistically with each other and the ECM.
Cells can adhere to the substrate through four different mechanisms: focal complexes, focal adhesions, fibrillary adhesions, and 3D matrix adhesions (Cukierman et al. 2001). Focal complexes are transient adhesion structures seen in the early stages of the adhesion process, and usually occur while cells migrate and spread. Focal complexes are multi-molecular structures, and integrin, talin, paxillin, vinculin, and actin proteins play a role in the formation of these structures (Geiger and Yamada 2011; Zaidel-Bar et al. 2003). Focal complexes are transformed into focal adhesions by the incorporation of zyxin into these proteins and tyrosine phosphorylation (Beningo et al. 2001; Li et al. 2016). Focal adhesions are more stable and well-regulated structures than focal complexes. Focal adhesions transform into fibrillary adhesions and become more stable. Cell adhesion, migration, and spreading in 2D substrates are limited by the x-y plane. Cell polarity occurs automatically to the extent allowed by the continuous, stiff, and flat surface. In 3D matrix adhesion, in contrast to 2D, cells are spatially associated with the 3D matrix in all directions. Cell behavior, structure, and properties in 3D are very different from 2D culture systems, and this type of adhesion is the closest way of simulating cell adhesion in vivo (Li et al. 2016). 3D environments provide more degrees of freedom; thus, the cells can increase their communication with the surrounding cells, as well as the ECM (Duval et al. 2017). In models where cells are embedded in the ECM in 3D systems, the basal-apical polarization occurs simultaneously, and thus cells can show their true phenotypes. In addition, the growth factors produced by the cells in 2D cultures are removed while changing the medium; however, in 3D systems, the growth factors bind to ECM proteins and contribute to the formation of the gradient within the ECM (Muncie and Weaver 2018). Gradients of soluble biomolecules, nutrients, and oxygen in ECM affect stem cell differentiation and morphogenesis (Baker and Chen 2012).
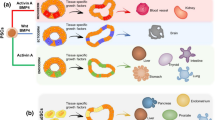
Organoids are 3D small tissues that are derived from stem cells, and that can self-organize and form organ-like structures in appropriate conditions of the microenvironment (Lancaster and Knoblich 2014). This collective self-organization leads into structures that closely resemble the embryonic architecture. “Self-organization” is an important and common tool of nature at many different levels, from molecular to population dynamics. From a physical point of view, it is defined as a system that starts with its parts separate (so that the behavior of each is independent of others’ states) and whose parts then form connections (Ashby 1962). Organoid formation, therefore, may be defined as a self-connecting, organized growth of a starting group of cells, in order to form and differentiate toward an ordered tissue structure based on local interactions.
In order to be named as an “organoid”, the structure must be formed of different cell types; these cells must be organized in accordance with the original organ and must be able to show at least some functions of the organ (Sugawara et al. 2018). For these reasons, organoids are usually obtained from stem cells that have unlimited differentiation and proliferation capacity. Although the term “organoid” is sometimes confused with cell “spheroids” or “aggregates”, there are significant differences between them. Spheroids are usually 3D micro- or macro-cell aggregates obtained using one or more cell lines. The cells in these structures are not organized as they are in the original organ and usually do not show organ functions. Although they are more similar to in vivo tissues than 2D cell culture systems, they are not as successful as organoids in this respect. However, organoids are the structures most closely resembling the original tissue both physically and functionally. They are generally formed from pluripotent stem cells (PSCs), adult stem cells, progenitor cells, or induced pluripotent stem cells (iPSCs) (Lancaster and Knoblich 2014; Yin et al. 2016). Stem cells used during organoid formation self-organize and differentiate into desired cell types by harmonic effects of biochemical molecules, appropriate biophysical conditions, external forces, and appropriate extracellular matrix structure and thus cell-matrix and cell-cell communication of the relevant organ (Watt and Huck 2013).
Since organoids are the most similar in vitro systems to the original organ, they have great potential to be used as organ models in drug development and personalized medicine studies. In addition, since they mimic embryonic development during their formation, they provide important information about human embryogenesis and organogenesis (Ho et al. 2018). To date, information about the human embryonic process has been mostly obtained from animal models, but since anatomical and physiological differences exist between species, observing this process with human cells now provides us with more accurate information. By autologous stem cells obtained from the person, organoids can be obtained as a complete match of the person’s own genetic makeup. Drug discovery and developmental studies with patient-specific organoids, which can be formed with iPSCs without needing a biopsy, are considered to be more effective and to give results much faster (Drost and Clevers 2017). While developing organoid disease models, information about the development of the disease can also be obtained. To date, many organoid studies have been carried out, but mostly intestinal (Ho et al. 2018; Nakamura and Sato 2018; Rahmani et al. 2018), brain (Allende et al. 2018; Lancaster et al. 2013; Mariani et al. 2015; Qian et al. 2016, 2019; Seo et al. 2017), liver (Fiorotto et al. 2018; Gaitantzi and Breitkopf-Heinlein 2018; Vyas et al. 2018), kidney (Kim et al. 2018), articular cartilage (Irie et al. 2008; Mizuno et al. 2016), lung, and stomach (Dutta et al. 2017; Xu et al. 2018) organoids have been studied. Because of the fact that it is difficult to control in vitro stem cell differentiation, spatial organization and the interactions of cells with each other and the matrix, most of the obtained organoids have been formed to contain only one or few components of a tissue (Yin et al. 2016). Modeling of individual genetic diseases can be carried out with organoids combined with genetic engineering, genomic editing, transcriptomics, and epigenetic studies of individual PSCs or iPSCs (Ardhanareeswaran et al. 2017). Since individual tumor organoids can be obtained with cells isolated from the patient, even rare-type cancers can be modeled in this way and used in drug development studies (Ho et al. 2018).
Engineering the organoid microenvironment
It is inherently difficult to replicate the organoid studies due to self-organizing character. Each organoid produced cannot attain the same shape and size, as well as the spatial organization of the cells and cell compositions (Karzbrun and Reiner 2019). The engineered microenvironment of the cells during the formation of organoids is the main factor that determines the organoid fate. Culture media is only one of the input signals that determine the outcome from the initial cellular population. The most important components of the microenvironment are growth factors, morphogens, cell-cell interactions, and ECM due to surrounding cell diversity and localization (Eshghi and Schaffer 2008). Although the composition and structure of ECM vary with each tissue, it is a complex matrix structure that generally contains glycoproteins, proteoglycans, and fibrous proteins, such as collagen, laminin, fibronectin, and elastin produced by the endoplasmic reticulum and Golgi apparatus of the cells (Alberts et al. 2002). All these ECM components not only provide structural support to the cells by forming the physical microenvironment of the tissues, but also directly interact with the receptors of the integrins, transmembrane proteins located within the cell membrane, and directly affect cell migration, spreading, differentiation, and proliferation (Gattazzo et al. 2014). Thus, when integrins cannot find adhesion contact in the absence of ECM, the cells respond to this situation with a kind of apoptosis, specifically named anoikis (Frisch and Francis 1994). Therefore, ECM has vital importance on the cells. However, it is not sufficient for 3D cell culture and tissue engineering studies to have a surface for cells to adhere to, where ECM is only partially mimicked. ECM protein isoforms may even cause undesirable effects on the cells. Due to activated different signaling pathways by an undesired ECM ligand or silenced signaling pathways by a lacking ligand in the microenvironment, the cells may lose their phenotypes or tend to go through malignant transformation or apoptosis (Gattazzo et al. 2014). Biochemical signals should be given to the system at the appropriate concentration, place, and time to produce more realistic organoid formation to in vivo (Yin et al. 2016).
In vitro organogenesis has been modeled as a non-linear deterministic system (Dahl-Jensen et al. 2016). Most biological systems are non-linear, since it is impossible to isolate certain input parameters from the rest of the organism and its surroundings, history, genetic make-up, and epigenetic modifications (Turner et al. 2016). Similarly, different culture conditions can lead to preserved stem cell behavior, as well as differentiation toward different lineages (Greggio et al. 2013; Fordham et al., 2013). A third type of outcome is usually a combination of the two cases, where a group of progenitors are conserved, while other groups can be differentiated even toward more than one phenotype—a condition that can be defined as multiple-equilibria (Dahl-Jensen et al. 2016). Small and usually uncontrollable changes in initial conditions act as bifurcation points that increase the number of possible equilibria (Todhunter et al. 2015; Ungrin et al. 2012). This can be treated as the main reason for low reproducibility in current organoid research.
Mechanosensing of the ECM and external forces
In addition to the biochemical composition of the microenvironment, biomechanical properties of the ECM, such as stiffness, viscoelasticity, pore size, porosity, permeability, thickness, size, shape, and topography characteristics are known to be of great importance on cell behavior (Akhmanova et al. 2015). A comprehensive view of mechanobiology may be considered as the one that focuses on the effects of structural micro-environment and force-induced deformations on molecules, cells, and tissues.
Cells sense surface stiffness and elasticity with focal adhesion complexes, including adhesion molecules, such as integrin and cadherin, and these proteins bind ECM to the actomyosin cytoskeleton of the cell. Cytoskeleton contractions transmit this information to the nucleus by mechanosensing and determine how the cell reacts through the nucleus matrix, nucleus envelope, and chromatins (Gattazzo et al. 2014). The elastic modulus of the plastic culture vessels at the GPa level used in 2D cell culture causes changing cell differentiation, migration, spatial organization, and self-renewal properties of mammalian cells, since these cells show appropriate cellular behavior in the elastic modulus of the ECM at the kilopascal level. For example, in a study using human mesenchymal stem cells (MSC), it was shown that MSCs express tissue-specific transcription factors on mimicking ECM stiffness of specific tissues (Engler et al. 2006). In another study using neural stem cells (NSC), it was shown that NSCs showed neuronal differentiation on surfaces < 1 kPa, astrocytic differentiation on surfaces between 1 kPa and 3.5 kPa, and oligodendrocytic differentiation on surfaces > 7 kPa (Leipzig and Shoichet 2009). In addition, increased ECM stiffness can cause diseases such as cancer and fibrosis, because cancer tissues are stiffer than normal tissues (Jansen et al. 2015). For this reason, it is very important to select a material suitable for the ECM stiffness of the desired organ when forming the organoid. Researchers often use hydrogels, which have high water-holding capacity, for this purpose. The most commonly used hydrogel in organoid studies is Matrigel for supporting cells physically and for biochemical signaling cues (Yin et al. 2016). Matrigel can be used for most types of cells as it promotes growth and provides a physiologically adequate medium for soluble biomolecules relative to synthetic scaffolds. However, since the content of Matrigel is not completely known, it may contain biomolecules inappropriate for the used cell type or may not contain specific ECM components. Apart from this, it can show lot-to-lot variability (Fang and Eglen 2017), leading to experimental unrepeatability. It is important to move away from Matrigel and other culture components with spatial and temporal variability (like serum), and establish more standardized media formulations and defined synthetic or natural hydrogels with controllable chemical composition and physical properties, such as porosity, viscoelasticity, and stiffness. The scaffolds can be designed by selecting the polymer according to the properties of the desired tissue and thus, providing cells to show the most similar behavior to the original tissue.
During organogenesis, cells are also exposed to many different external mechanical stimuli. Shear stress, cardiac contraction, fetal breathing movement, and traction force can be given as examples (Park et al. 2019). External forces have an important effect on stem cells. In the absence of these forces, desired stem cell differentiation, spatial organization, and organ functions cannot be observed and mature organoid development cannot be achieved. The forces can be applied directly to the cell or indirectly through the ECM, and thus the cell can mechanosense the forces and react while forming an organoid. The amplitude of the force applied during organoid development is an important parameter; however, the duration of application, being a static or dynamic force, and frequency are also other important parameters. The reactions of the cells as a result of the effects of external forces are called mechanotransduction. For example, continuous blood flow from the circulatory system and airflow through the respiratory system create shear stress on endothelial and epithelial cells, respectively, or compression and tension are imposed on the cells during muscle contraction. Each external force applied to the cell creates tension on the cytoskeleton and has an effect on the geometry, organization, and behavior of the cell (Park et al. 2019).
Mechanotransduction is initiated at the local cell membrane-force interface by inducing local conformational changes of proteins, similar to soluble ligand-induced signal transduction (Na et al. 2008). It is generally postulated that the initial event of mechanotransduction results in a biochemical cascade of events that reaches the nucleus; such that, the external forces are detected by focal adhesion proteins and mechanosensed by the nucleus through mechanotransduction signaling pathways, such as Ras/MAPK, P13K/Akt, RhoA/ROCK, Wnt/β-Catenin, and TGF-β (Chan et al. 2017; Gattazzo et al. 2014).
However, most important is the notion that there is a direct mechanical signal movement from the focal adhesion deformation through the cytoskeleton all the way to the chromatin inside the nucleus (Tajik et al. 2016; Swift et al. 2013; Kirby and Lammerding 2018; Mammoto et al. 2013; Stachowiak et al. 2014; Miroshnikova et al. 2017). It is more plausible that these two pathways, i.e., biochemical and mechanical (excluding here possible electrical pathways), interact and modify each other on their way to the nucleus (Na et al. 2008; Stachowiak et al. 2014).
Different ion channels, including NMDA receptors, TREK-1, and TRPC1, are shown to be mechanosensitive, and be activated by the tension developed in the cell membrane on a millisecond time scale. Particularly MEC-4/MEC-10, DEG/ENaC, TRP, and Piezo channels are shown to act as cytoskeleton-extracellular matrix linkages (Martinac 2014). Piezo1 and Piezo2 proteins are also shown to act as pore-forming subunits of mechanically activated ion channels (Coste et al. 2012; Qiu and Müller, 2018). It has been well established that stretch-activated ion channels alter cardiac electrical activity and the influx of Ca2+ and Na+ ions, and regulate cardiac contractility via protein phosphorylation (Baumgarten 2013; Kohl 2018).
Simplified schematics of the connectivity of the various proteinous structural elements, both inside the cytosol and the nucleus, comprising actin filaments, microtubules, intermediate filaments, the LINC complex, lamins, and nuclear actins, to name a few, are found in many reviews (Tajik et al. 2016; Swift et al. 2013; Kirby and Lammerding 2018; Mammoto et al. 2013; Miroshnikova et al. 2017). A compilation of the existing knowledge suggests that the mechanical signal transduction is very rapid, as reported by the activation of various cytosolic reporters in living cells (Na et al. 2008; Pertz et al. 2006). A postulation has been put forward recently, named mechanoepigenetics, suggesting that all mechanotransduction pathways, originating at cell-matrix interfaces, cell-cell interfaces, and flowing media-cell interfaces are integrated in their way to the nucleus and chromatin, where they mechanically rearrange the chromatin for epigenetic manifestations (Missirlis 2016).
Another key question that is in need of urgent answers is how the cellular mechanics and signaling transduction are influencing tissue mechanics and intercellular mechanical communication during embryogenesis and development in general. Most probably there is a two-way communication. In recent years, organoids, as mentioned earlier, are extensively studied, in 3D experimental conditions, and, by their nature, get a lot of information and maybe feedback from basic knowledge of developmental biology, of growth and patterning. It seems that the molecules, termed morphogens, can be transported in several different ways across cells, while both agreements and controversies still exist on how morphogen gradients are formed and interpreted, which signaling pathways feedback on morphogenesis, how patterns emerge, and how growth is controlled, just to name a few (Chan et al. 2017; Stathopoulos and Iber 2013). Outside forces are sensed by cells through mechanotransduction; however, specific forces are also generated within the cells themselves, as they are necessary for many physiological processes, such as cell division, migration, and cell attachment. Such forces, produced within or outside the nucleus, may be subtler than the externally transduced ones. Very little is known on the significance of this chorus of forces interrogated by the nucleus, which may be considered as a mechanosensor of its own (Kirby and Lammerding 2018). However, some insights are already proposed, either by coupling mechanical events (Suzuki et al. 2016) or by gaining new knowledge on the function of specific mechanical structures, like the microtubules (Zijl and Lomakin (2019); Portrian et al. 2017).
With regard to the sensing of mechanical cues by cells in organoids, the situation is quite complex and only recently a combination of scant experimental results, complemented by theoretical models, starts to give hints of possible operating mechanisms. As mentioned before in single cells (Chan et al. 2017), several distinct proteins sense and respond to different mechanical cues, for example, zyxin and paxillin in focal adhesions alter binding kinetics, E-cadherin bounds β-catenin is active in cell-cell communication, stretched Piezo 1 and 2 channels regulate calcium ion movement, and the Yap (Yes-associated protein)/Taz (transcriptional coactivator with PDZ-binding motif, WWTR1) system interacts with specific transcription factors to regulate genetic/epigenetic modifications (Martino et al. 2018; Dupont et al. 2011), and all these cues are orchestrated in ways that are still being investigated. It is possible that transmission of forces across multiple scales is essential for positional information and responsible for the reaction-diffusion model (Chan et al. 2017). Also, during morphogenesis, mechanical constraints may arise due to tissue bending or buckling, which generate long-range tissue stress. One probable corollary of this state is that there is an inter-play between mechanical signals and cellular polarity (Chan et al. 2017). During physiological morphogenesis, as well as organoid formation, the growing number of cells is communicating among themselves in order to achieve the spectacular spatiotemporal arrangement that constitutes the developing tissues and organs. Cells often migrate collectively, in a coordinated polarity, while maintaining their cell-to-cell contacts (Sunyer and Trepat 2017). Traction force microscopy has revealed that apart from the role of actin-based mechanical forces, microtubules also play an important role in maintaining the polarized shape of the cells and in coordinating cell behavior during tissue remodeling (Singh et al. 2018). In a recent publication, it is demonstrated that during epithelial morphogenesis, the apico-basal forces in cells (including here the apoptotic cells, which contribute to the process mechanically) rely on the contractile actomyosin structure that connects the apical surface to the nucleus, while the nucleus is anchored to basal adhesions, participating itself in the force production necessary for the morphogenetic actions (Ambrosini et al. 2019).
Another investigation on effect of polarity and cell division dynamics on cystogenesis has shown that cell-to-cell contact topology can discriminate mechanical equilibrium and non-equilibrium states (Cerruti et al. 2013). Their in vitro and in vivo observations, backed up with mathematical modeling, show that cystogenesis occur under energetically unfavorable conditions, and in a mechanical non-equilibrium state, where cell rearrangements extend further from neighboring exchange interactions toward long-range motility. High cell division rate was also associated with multiple lumen formation and fast cyst growth. It is important to note that growth of healthy tissues, such as optic cup, cornea, and retina have been associated with highly ordered close-to-equilibrium states (Eiraku et al. 2012; Bhat 2001; Jalbert et al. 2003; Hofer et al. 2005), where short-range cell-to-cell interactions are predominant (Amonlirdviman et al. 2005; Classen et al. 2005).
As it is demonstrated in an interesting publication (Ellison et al. 2016), the cell-cell communication enhances the overall cell assembly (organoid for example) to sense shallow gradients during morphogenesis. As reliable gradient sensing is of paramount importance during, for example, epithelial branching morphogenesis, it was experimentally demonstrated that extremely weak gradients of a growth factor is sensed and utilized by cell ensembles but not by individual cells (Ellison et al. 2016). This “collective sensing” is most likely mediated by cell-cell communications through gap junctions and local feedback systems. An educated guess is that also extremely weak forces, not sensed by individual cells, may be robustly sensed and transduced by cell ensembles. This is an issue for exploration.
In silico organoid models
Control of biochemical, mechanical, and chemical stimuli is known to have many effects on stem cells during organoid formation is essential. Yet, the orchestrated effects of these mechanisms are not fully understood and are very difficult to control; and undesirable changes in stem cells during organoid formation may occur under unpredictable effects. Realistic in silico organoid models can be a powerful tool to develop a better understanding of organogenesis and to develop therapeutic strategies faster, and in a controlled, efficient, and cost-effective manner (Karolak et al. 2018). With the help of in silico models, better controlled systems can be constructed and experimental design optimization can be provided for in vitro organoid formation (Poli et al. 2019). Furthermore, with the help of these models, organoid growth, cell differentiation, and functionality can be predicted while effects of certain manipulations can also be foreseen. Up to date, limited studies have been carried out with in silico models, such as modeling transport, consumption, and diffusion of oxygen in organoids (Berger et al. 2018; Buske et al. 2012), modeling the effects of short-range growth inhibitory signals (Dahl-Jensen et al. 2016), shape transformations of epithelial cells (Misra et al. 2016) or behavior of multi-cellular systems by cellular growth, signaling, and migration (Milde et al. 2014). In addition to such studies that provide a better understanding of in vivo and in vitro organogenesis, tumor development, progression, and responses to the treatment studies have been performed with in silico tumor organoids (Karolak et al. 2018). In silico organoid models have many deficiencies at the moment, but it is foreseen that they will develop rapidly in parallel to in vitro organoid studies.
Future perspectives
Organoids are important tools for drug development, disease modeling, and personalized medicine studies, since they simulate the self-organization of tissues during embryogenesis. But we still need to accept that there are major differences between in vivo embryogenesis or organogenesis and in vitro organoid formation, since in vitro conditions, no matter how well-controlled, are extremely different than real, regionally defined physiological conditions. On the other hand, organoids allow researchers to select and isolate certain biochemical and/or physical cues, and study their respective and coordinated effects on tissue or disease development and regeneration. In particular, they can also serve as great tools for studying the effects external and internal mechanical forces and their interactions with molecular signaling pathways during embryogenesis and organogenesis, which is still an unexplored field.
It is not trivial to design a culture system that is well-controlled to produce reproducible results, while preserving self-organizing capacity. Newly emerging microfabrication, 3D printing, and bioprinting techniques combined with appropriately designed dynamic systems, including microfluidics, may offer opportunities to improve organoid research.
References
Akhmanova M, Osidak E, Domogatsky S, Rodin S, Domogatskaya A (2015) Physical, spatial, and molecular aspects of extracellular matrix of in vivo niches and artificial scaffolds relevant to stem cells research. Stem Cells Int 35
Alberts B, Johnson A, Lewis J, Raff M, Roberts K and Walter P (2002) The extracellular matrix of animals. In: Molecular biology of the cell. 4th edition. New York Garland Science. https://www.ncbi.nlm.nih.gov/books/NBK26810/. Accessed 24 June 2019
Allende ML et al (2018) Cerebral organoids derived from Sandhoff disease-induced pluripotent stem cells exhibit impaired neurodifferentiation. J Lipid Res 59:550–563
Ambrosini A et al (2019) Mechanical function of the nucleus in force generation during epithelial morphogenesis. Dev Cell 50:1–15
Amonlirdviman K, Khare NA, Tree DR, Chen WS, Axelrod JD, Tomlin CJ (2005) Mathematical modeling of planar cell polarity to understand domineering nonautonomy. Science 307:423–426
Ardhanareeswaran K, Mariani J, Coppola G, Abyzov A, Vaccarino FM (2017) Human induced pluripotent stem cells for modelling neurodevelopmental disorders. Nat Rev Neurol 13:265–278
Ashby WR (1962) Principles of the self-organizing system. In: Von Foerster H, Zopf GW (eds) Principles of self-organization: transactions of the University of Illinois Symposium. Pergamon Press, London, UK, pp 255–278
Baker BM, Chen CS (2012) Deconstructing the third dimension–how 3D culture microenvironments alter cellular cues. J Cell Sci 125:3015–3024
Baumgarten CM (2013) Origin of mechanotransduction: stretch-activated ion channels. In: Madame curie bioscience database [Internet]. Austin (TX), Landes Bioscience, pp 2000–2013 Available from: https://www.ncbi.nlm.nih.gov/books/NBK6374/. Accessed 27 June 2019
Beningo KA, Dembo M, Kaverina I, Small JV, Wang Y-l (2001) Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J Cell Biol 153:881–888
Berger E et al (2018) Millifluidic culture improves human midbrain organoid vitality and differentiation. Lab Chip 18:3172–3183
Bhat SP (2001) The ocular lens epithelium. Biosci Rep 21:537–563. https://doi.org/10.1023/A:1017952128502
Buske P, Przybilla J, Loeffler M, Sachs N, Sato T, Clevers H, Galle J (2012) On the biomechanics of stem cell niche formation in the gut–modelling growing organoids. FEBS J 279:3475–3487
Cerruti B et al (2013) Polarity, cell division, and out-of-equilibrium dynamics control the growth of epithelial structures. J Cell Biol 203(2):359–372
Chan CJ, Heisenberg C-P, Hiiragi T (2017) Coordination of morphogenesis and cell-fate specification in development. Curr Biol 27:R1024–R1035
Classen AK, Anderson KI, Marois E, Eaton S (2005) Hexagonal packing of Drosophila wing epithelial cells by the planar cell polarity pathway. Dev Cell 9:805–817. https://doi.org/10.1016/j.devcel.2005.10.016
Coste et al (2012) Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 483:176–181
Cukierman E, Pankov R, Stevens DR, Yamada KM (2001) Taking cell-matrix adhesions to the third dimension. Science 294:1708–1712
Dahl-Jensen SB, Figueiredo-Larsen M, Grapin-Botton A, Sneppen K (2016) Short-range growth inhibitory signals from the epithelium can drive non-stereotypic branching in the pancreas. Phys Biol 13:016007
Drost J, Clevers H (2017) Translational applications of adult stem cell-derived organoids. Development 144:968–975
Dupont et al (2011) Role of YAP/TAZ in mechanotransduction. Nature 474:179–183
Dutta D, Heo I, Clevers H (2017) Disease modeling in stem cell-derived 3D organoid systems. Trends Mol Med 23:393–410
Duval K, Grover H, Han L-H, Mou Y, Pegoraro AF, Fredberg J, Chen Z (2017) Modeling physiological events in 2D vs. 3D cell culture. Physiology 32:266–277
Eiraku M, Adachi T, Sasai Y (2012) Relaxation-expansion model for self-driven retinal morphogenesis. Bioessays 34(1):17–25
Ellison D et al (2016) Cell-cell communication enhances the capacity of cell ensembles to sense shallow gradients during morphogenesis. Proceedings of the National Academy of Sciences of the United States of America (PNAS) 113(6):E679–88
Engler AJ, Sen S, Sweeney HL, Discher DE (2006) Matrix elasticity directs stem cell lineage specification. Cell 126:677–689
Eshghi S, Schaffer DV (2008) Engineering microenvironments to control stem cell fate and function. In: StemBook [Internet]. Ed. Lisa Girard Harvard Stem Cell Institute. https://doi.org/10.3824/stembook.1.5.1
Fang Y, Eglen RM (2017) Three-dimensional cell cultures in drug discovery and development. Slas discovery: Advancing Life Sciences R&D 22:456–472
Fiorotto R, Amenduni M, Mariotti V, Fabris L, Spirli C, Strazzabosco M (2018) Liver diseases in the dish: iPSC and organoids as a new approach to modeling liver diseases. Biochim Biophys Acta, Mol Basis Dis 1865(5):920–928. https://doi.org/10.1016/j.bbadis.2018.08.038
Fordham RP, Yui S, Hannan NRF, Soendergaard C, Madgwick A et al (2013) Transplantation of expanded fetal intestinal progenitors contributes to colon regeneration after injury. Cell Stem Cell 13:734–744
Frisch SM, Francis H (1994) Disruption of epithelial cell-matrix interactions induces apoptosis. J Cell Biol 124:619–626
Gähwiler B, Capogna M, Debanne D, McKinney R, Thompson S (1997) Organotypic slice cultures: a technique has come of age. Trends Neurosci 20:471–477
Gaitantzi H, Breitkopf-Heinlein K (2018) Spontaneous self-assembly of liver organoids from differentiated human cells: human liver organoids. In: Davies JA, Lawrence ML (eds) Organs and organoids. Elsevier, Part 2, Chapter 7 145–156
Gattazzo F, Urciuolo A, Bonaldo P (2014) Extracellular matrix: a dynamic microenvironment for stem cell niche. Biochim Biophys Acta, Gen Subj 1840:2506–2519
Geiger B, Yamada KM (2011) Molecular architecture and function of matrix adhesions. Cold Spring Harb Perspect Biol 3(5):a005033. https://doi.org/10.1101/cshperspect.a005033
Greggio C, De Franceschi F, Figueiredo-Larsen M, Gobaa S, Ranga A, Semb H, Lutolf M, Grapin-Botton A (2013) Artificial three-dimensional niches deconstruct pancreas development in vitro. Development 140:4452–4462
Ho B, Pek N, Soh B-S (2018) Disease modeling using 3D organoids derived from human induced pluripotent stem cells. Int J Mol Sci 19:936
Hofer H, Carroll J, Neitz J, Neitz M, Williams DR (2005) Organization of the human trichromatic cone mosaic. J Neurosci 25:9669–9679. https://doi.org/10.1523/JNEUROSCI.2414-05.2005
Irie Y, Mizumoto H, Fujino S, Kajiwara T (2008) Development of articular cartilage grafts using organoid formation techniques. Transplant Proc 40(2):631–633
Jackson EL, Lu H (2016) Three-dimensional models for studying development and disease: moving on from organisms to organs-on-a-chip and organoids. Integr Biol 8:672–683
Jalbert I, Stapleton F, Papas E, Sweeney DF, Coroneo M (2003) In vivo confocal microscopy of the human cornea. Br J Ophthalmol 87:225–236. https://doi.org/10.1136/bjo.87.2.225
Jansen KA, Donato DM, Balcioglu HE, Schmidt T, Danen EH, Koenderink GH (2015) A guide to mechanobiology: where biology and physics meet. Biochim Biophys Acta, Mol Cell Res 1853(11B):3043–3052
Kapałczyńska M et al (2018) 2D and 3D cell cultures–a comparison of different types of cancer cell cultures. Arch Med Sci 14(4):910–919. https://doi.org/10.5114/aoms.2016.63743
Karolak A, Markov DA, McCawley LJ, Rejniak KA (2018) Towards personalized computational oncology: from spatial models of tumour spheroids, to organoids, to tissues. J R Soc Interface 15:20170703. https://doi.org/10.1098/rsif.2017.0703
Karzbrun E, Reiner O (2019) Brain organoids—a bottom-up approach for studying human neurodevelopment. Bioengineering 6(1):9. https://doi.org/10.3390/bioengineering6010009
Kim YK, Nam SA, Yang CW (2018) Applications of kidney organoids derived from human pluripotent stem cells. Korean J Intern Med 33(4):649–659
Kirby TJ, Lammerding J (2018) Emerging views of the nucleus as a cellular mechanosensor. Nat Cell Biol 20:373–381
Kohl P (2018) Cardiac stretch-activated channels and mechano-electric coupling. In: Zipes DP, Jalife J, Stevenson WG (eds) Cardiac electrophysiology: from cell to bedside (Seventh Edition). Elsevier Inc. Chapter 14 128–139
Lancaster MA, Knoblich JA (2014) Organogenesis in a dish: modeling development and disease using organoid technologies. Science 345(6194):1247125. https://doi.org/10.1126/science.1247125
Lancaster MA et al (2013) Cerebral organoids model human brain development and microcephaly. Nature 501:373–379
Leipzig ND, Shoichet MS (2009) The effect of substrate stiffness on adult neural stem cell behavior. Biomaterials 30:6867–6878
Li Z, Lee H, Zhu C (2016) Molecular mechanisms of mechanotransduction in integrin-mediated cell-matrix adhesion. Exp Cell Res 349:85–94. https://doi.org/10.1016/j.yexcr.2016.10.001
Mammoto T, Mammoto A, Ingber DE (2013) Mechanobiology and developmental control. Annu Rev Cell Dev Biol 29:27–61
Mariani J et al (2015) FOXG1-dependent dysregulation of GABA/glutamate neuron differentiation in autism spectrum disorders. Cell 162:375–390
Martinac B (2014) The ion channels to cytoskeleton connection as potential mechanism of mechanosensitivity. Biochim Biophys Acta 1838:682–691
Martino F, Perestrelo AR, Vinarský V, Pagliari S, Forte G (2018) Cellular mechanotransduction: from tension to function. Front Physiol 9:824
Milde F, Tauriello G, Haberkern H, Koumoutsakos P (2014) SEM++: a particle model of cellular growth, signaling and migration. Comput Particle Mech 1(2):211–227
Miroshnikova YA, Nava MM, Wickstroem SA (2017) Emerging roles of mechanical forces in chromatin regulation. J Cell Sci 130:2243–2250
Misra M, Audoly B, Kevrekidis IG, Shvartsman SY (2016) Shape transformations of epithelial shells. Biophys J 110(7):1670–1678. https://doi.org/10.1016/j.bpj.2016.03.009
Missirlis YF (2016) Mechanoepigenetics. Front Cell Dev Biol 4:113
Mizuno S, Takada E, Fukai N (2016) Spheroidal organoids reproduce characteristics of longitudinal depth zones in bovine articular cartilage. Cells Tissues Organs 202:382–392
Muncie JM, Weaver VM (2018) The physical and biochemical properties of the extracellular matrix regulate cell fate. Curr Top Dev Biol 130:1–37
Na S et al (2008) Rapid signal transduction in living cells is a unique feature of mechanotransduction. PNAS 105(18):6626–6631
Nakamura T, Sato T (2018) Advancing intestinal organoid technology toward regenerative medicine. Cell Mol Gastroenterol Hepatol 5:51–60
Park SE, Georgescu A, Huh D (2019) Organoids-on-a-chip. Science 364:960–965
Pertz O et al (2006) Spatiotemporal dynamics of RhoA activity in migrating cells. Nature 440:1069–1072
Poli D, Magliaro C, Ahluwalia A (2019) Experimental and computational methods for the study of cerebral organoids: a review. Front Neurosci 13:162
Portrian D et al (2017) Tubulin acetylation protects long-lived microtubules against mechanical ageing. Nat Cell Biol 19:391–398
Qian X et al (2016) Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell 165:1238–1254
Qian X, Song H, Ming G-L (2019) Brain organoids: advances, applications and challenges. Development 146:dev166074. https://doi.org/10.1242/dev.166074
Qiu X, Müller U (2018) Mechanically gated ion channels in mammalian hair cells. Front Cell Neurosci 12:100
Rahmani S, Breyner NM, Su H-M, Verdu EF, Didar TF (2018) Intestinal organoids: a new paradigm for engineering intestinal epithelium in vitro. Biomaterials 194:195–214
Seo J et al (2017) Inhibition of p25/Cdk5 attenuates tauopathy in mouse and iPSC models of frontotemporal dementia. J Neurosci 37:9917–9924
Singh A et al (2018) Polarized microtubule dynamics directs cell mechanics and coordinates forces during epithelial morphogenesis. Nat Cell Biol 20:1126–1133
Stachowiak MR et al (2014) A mechanical-biochemical feedback loop regulates remodeling in the actin cytoskeleton. PNAS 111(49):17528–17533
Stathopoulos A, Iber D (2013) Studies of morphogens: keep calm and carry on. Development 140:4119–4124
Sugawara T, Sasaki K, Akutsu H (2018) Organoids recapitulate organs? Stem Cell Investig 5:3. https://doi.org/10.21037/sci.2018.01.02
Sunyer R, Trepat X (2017) Mechanobiology of collective cell systems. Biofisica 7:1–7 http://biofisica.info. Accessed 18 June 2019
Suzuki A et al (2016) How the kinetochore couples microtubule force and centromere stretch to move chromosomes. Nat Cell Biol 18:382–392
Swift J et al (2013) Nuclear lamin-A scales with tissue stiffness and enhances matrix-directed differentiation. Science 341(6149):1240104. https://doi.org/10.1126/science.1240104
Tajik A et al (2016) Transcription upregulation via force-induced direct stretching of chromatin. Nat Mater 15:1287–1296
Todhunter ME, Jee NY, Hughes AJ, Coyle MC, Cerchiari A, Farlow J, Garbe JC, LaBarge MA, Desai TA, Gartner ZJ (2015) Programmed synthesis of three-dimensional tissues. Nat Methods 12:975–981
Turner DA, Baillie-Johnson P, Martinez Arias A (2016) Organoids and the genetically encoded self-assembly of embryonic stem cells. BioEssays 38:181–191
Ungrin MD, Clarke G, Yin T, Niebrugge S, Nostro MC, Sarangi F, Wood G, Keller G, Zandstra PW (2012) Rational bioprocess design for human pluripotent stem cell expansion and endoderm differentiation based on cellular dynamics. Biotechnol Bioeng 109:853–866
Vyas D et al (2018) Self-assembled liver organoids recapitulate hepatobiliary organogenesis in vitro. Hepatology 67:750–761
Watt FM, Huck WT (2013) Role of the extracellular matrix in regulating stem cell fate. Nat Rev Mol Cell Biol 14(8):467–473
Xu H, Jiao Y, Qin S, Zhao W, Chu Q, Wu K (2018) Organoid technology in disease modelling, drug development, personalized treatment and regeneration medicine. Exp Hematol Oncol 7:30. https://doi.org/10.1186/s40164-018-0122-9
Yin X, Mead BE, Safaee H, Langer R, Karp JM, Levy O (2016) Engineering stem cell organoids. Cell Stem Cell 18:25–38
Zaidel-Bar R, Ballestrem C, Kam Z, Geiger B (2003) Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J Cell Sci 116:4605–4613
Zijl S, Lomakin AJ (2019) The “nuclear physics” behind genetic and epigenetic control of cell fate. Exp Cell Res 376(2):236–239
Acknowledgments
The authors acknowledge COST Action 16122 (BIONECA) for financing Prof. Dr. Yannis F. Missirlis for an STSM to visit and collaborate with the rest of the authors at Ege University; COST Action 16217 (ENIUS) for designating him as a disseminator to “promote” the Action at 4th international Symposium on Nanoengineering for Mechanobiology (N4M), where some of the ideas presented in this review were discussed; and Republic of Turkey Ministry of Development [EGEMATAL;2010K120810] for financing Dr. Ece Bayir.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bayir, E., Sendemir, A. & Missirlis, Y.F. Mechanobiology of cells and cell systems, such as organoids. Biophys Rev 11, 721–728 (2019). https://doi.org/10.1007/s12551-019-00590-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12551-019-00590-7