
Abstract
Populations in malaria endemic areas are frequently exposed to mycotoxin-contaminated diets. The possible toxicological outcome of co-exposure to dietary aflatoxin B1 (AFB1) and artemisinin-based combination therapy warrants investigation to ascertain amplification or attenuation of cellular injury. Here, we investigated the neurobehavioral and biochemical responses associated with co-exposure to anti-malarial drug coartem, an artemether-lumefantrine combination (5 mg/kg body weight, twice a day and 3 days per week) and AFB1 (35 and 70 µg/kg body weight) in rats. Motor deficits, locomotor incompetence, and anxiogenic-like behavior induced by low AFB1 dose were significantly (p < 0.05) assuaged by coartem but failed to rescue these behavioral abnormalities in high AFB1-dosed group. Coartem administration did not alter exploratory deficits typified by reduced track plot densities and greater heat map intensity in high AFB1-dosed animals. Furthermore, the reduction in cerebral and cerebellar acetylcholinesterase activity, anti-oxidant enzyme activities, and glutathione and thiol levels were markedly assuaged by coartem administration in low AFB1 group but not in high AFB1-dosed animals. The significant attenuation of cerebral and cerebellar oxidative stress indices namely reactive oxygen and nitrogen species, xanthine oxidase activity, and lipid peroxidation by coartem administration was evident in low AFB1 group but not high AFB1 dose. Although coartem administration abated nitric oxide level, activities of myeloperoxidase, caspase-9, and caspase-3 in animals exposed to both doses of AFB1, these indices were significantly higher than the control. Coartem administration ameliorated histopathological and mophometrical changes due to low AFB1 exposure but not in high AFB1 exposure. In conclusion, contrary to AFB1 alone, behavioral and biochemical responses were not altered in animals singly exposed to coartem. Co-exposure to coartem and AFB1 elicited no additional risk but partially lessened neurotoxicity associated with AFB1 exposure.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The discovery of efficient anti-malarial drugs has contributed significantly to the decrease of malaria infection and its associated morbidity and mortality. The derivatives of artemisinin exhibit greater anti-malarial activity than the parent artemisinin because they pass through the blood–brain barrier easily to elicit its therapeutic action (Gautam et al. 2009; Zhang et al. 2021). Moreover, artemisinin derivatives reportedly exhibit several health beneficial effects namely anti-inflammatory, anti-microbial, anti-viral, immunomodulatory, anti-cancer, and neuroprotective effects (Ho et al. 2014; Meng et al. 2021). Unfortunately, due to the short half-lives of these drugs, the resistance of the parasite Plasmodium falciparum to the anti-malarial drugs has been reported (Li and Hickman 2011; Balikagala et al. 2021). However, effort to thwart the development of resistance to monotherapy involves the combination of artemisinin derivatives leading to the synthesis of longer-acting anti-malarial drugs commonly called artemisinin-based combination therapy (ACT) (Hernandez Maldonado and Grundmann 2022; Pernaute-Lau et al. 2022). The ACT remains the recommended first-line treatment for malaria by the World Health Organization (WHO 2020).
Coartem™, an anti-malarial drug comprising artemether and lumefantrine, represents the first ACT drug to be fully registered (Skelton-Stroud and Mull 1998). Coartem administration is an effective treatment for drug-resistant malaria principally caused by Plasmodium falciparum or mixed infections in both children and adults. Previous laboratory studies with animals demonstrated that administration of artemisinin derivatives to rats (12.5 mg/kg for 7 days), dogs (6 mg/kg for 28 days) and rhesus monkeys (8 mg/kg for 14 days) was associated with neurotoxicity characterized by balance deficits, altered movement, spasticity, brainstem tissue damage, and death (Classen et al. 1999; Kamchonwongpaisan et al. 1997; Petras et al. 2000; Li and Hickman 2011). The reported adverse effects of these drugs on the pons and medulla of rats, dogs, and monkeys have generated a major health concern, even though there are rare clinical reports related to neurotoxicity of ACT (Ramos-Martín et al. 2014).
Interestingly, in malaria endemic countries, there is a high rate of dietary contamination with mycotoxins produced by fungi before harvesting or during post-harvest storage (Wagacha and Muthomi 2008; Benkerroum 2020). The occurrence of aflatoxin B1 (AFB1), the most potent mycotoxins, in both animal and human foods is indeed associated with major health consequences (Rushing and Selim 2019, Sullivan 2022). For instance, acute aflatoxin exposure in humans resulted in abdominal pain, vomiting, convulsions, coma, and finally death with cerebral edema (Benkerroum 2020; Marin et al. 2013). AFB1 elicits neurotoxicity by disrupting the blood–brain barrier leading to the poisoning of vascular endothelial cells and the astrocytes which are essential for the brain environment homeostasis, neuronal survival, and synaptic information transmission (Baldissera et al. 2018; Qureshi et al. 2015). The neurobiochemical changes in laboratory animals exposed to AFB1 include induction of oxidative stress, impairment of antioxidant defense mechanisms, diminished cholinergic neurotransmission, and oxidative phosphorylation (Bahey et al. 2015; Souto et al. 2018; Adedara et al. 2021). The complete avoidance of dietary mycotoxin exposure is difficult due to the widespread agricultural foodstuff exchange, lack of visible warning labels on raw materials and finished products as well as ineffectual supervision on food security (IARC 2000; Adedara et al. 2014). Thus, dietary exposure to mycotoxins remains a major threat to animal and human health globally.
Owing to the prevalence of malaria, individuals undergoing ACT treatment may be exposed to dietary AFB1, thus predisposing such individuals to cellular responses which are not currently known. Chemotherapy has a good measure employed to fight against malaria is commonly abused via self-medication due to repeated ingestion of drugs in developing countries. Indeed, humoral and cellular responses in serum samples were assessed in vitro to verify the relationships between AFB1 exposure and the prevalence of malaria among children (Allen et al. 1992). Thus, it is crucial to provide necessary scientific evidence on the cellular responses associated with co-artem and AFB1 exposure to expand the extant knowledge about strategic risk exposure evaluation of xenobiotics. The systemic responses during exposure to more than one chemical can be heightened producing additive or synergistic effects or assuaged due to inhibition or antagonism when compared with each chemical effects (Karri et al. 2016; Adedara et al. 2017a, b). Thus, the present investigation was carried out to characterize, for the first time, the impact of co-exposure to co-artem and AFB1 on the neurobehavioral physiognomies, oxido-inflammatory stress, antioxidant defense mechanisms, and histological architecture in the cerebrum and cerebellum of rats singly or jointly exposed to co-artem and AFB1.
Materials and methods
Chemicals and assay kits
Coartem™ was obtained from Novartis Pharmaceutical Corporation, Horsham, UK. Thiobarbituric acid (TBA), 2′,7′-dichlorofluorescein diacetate (DCFH-DA), and 5,5-dithio-bis-2-nitrobenzoic acid (DTNB) were bought from Sigma Aldrich (St. Louis, MO, USA). Evaluation of caspase-9 and caspase-3 activities was performed using enzyme-linked immunosorbent assay kits (Elabscience Biotechnology Company, Beijing, China).
Housing and treatment of animals
Thirty (30) pubertal male Wistar rats collected from the Faculty of Veterinary Medicine, University of Ibadan were maintained under controlled housing conditions within the vivarium at the Department of Biochemistry. The animals had unhindered access to rodent chow and water. The investigation protocols were executed in accordance to the approved guidelines of the University of Ibadan Ethical Committee. The study composes of five groups of six animals each.
-
1.
Control group: rats orally dosed with corn oil at 2 mL/kg body weight.
-
2.
AFB1 alone: rats orally dosed with AFB1 alone at 70 µg/kg body weight.
-
3.
Coartem alone: rats orally dosed with coartem alone at 5 mg/kg (twice a day, every 12 h).
-
4.
Coartem + AFB11: rats orally dosed with coartem at 5 mg/kg (twice a day, every 12 h) and AFB1 at 35 µg/kg.
-
5.
Coartem + AFB12: rats orally dosed with coartem at 5 mg/kg (twice a day, every 12 h) and AFB1 at 70 µg/kg.
Coartem and AFB1 were separately dissolved in corn oil as a vehicle. Administration of coartem was 3 days per week, whereas AFB1 dosing was daily for 4 consecutive weeks. The doses of coartem and AFB1 as well as the duration of exposure used in this research were selected from previous pilot and published findings from our laboratory and other investigators (Adedara et al. 2021; El-Dakdoky 2009; Owumi et al. 2015; Nuwagira et al. 2021). Moreover, the 35 and 70 µg AFB1/kg are environmentally relevant and correspond to about 0.043 and 0.087 mg/kg in agricultural products. For instance, in Italy, the maximum level of AFB1 contamination was 728.8 and 1074 µg/kg in cotton and maize, respectively (Ferrari et al. 2022).
Behavioral testing
The assessment of behavioral characteristics was carried out in a novel wooden box (56 cm width by 56 cm length by 20 cm height), according to established protocol (Adedara et al. 2017b). In short, experimental animals were randomly chosen, placed inside the novel box, and allowed to freely explore the apparatus. A webcam (DNE webcam, Porto Alegre, Brazil) fixed directly above the box was used to video capture the activities of the rats for 7 min. Subsequently, the video-tracking software (ANY-maze, Stoelting Co, USA) was used to analyze the behavioral characteristics of the animals. To prevent interference, the box was cleaned using 70% ethanol and cotton wool each time a test is completed.
Exploratory, motor, and locomotor activities’ measurement
Adaptation of the animals to novelty stress was measured by specific exploratory, motor, and locomotor parameters. Behavioral parameters such as time of mobility, maximum speed, distance traveled, path efficiency, total body rotation, and absolute turn angle were used to evaluate locomotor and motor fitness. Moreover, the track plots and heat maps generated using the video-tracking software was used to evaluate the exploratory competence of the animals in the novel environment.
The limb strength of the animals was evaluated using the forelimb grip test in accordance with established method (Folarin et al. 2016; Adedara et al. 2019), while the negative geotaxis test was performed to measure the motor fitness according to established protocol (Motz and Alberts 2005; Adedara et al. 2019). The total time freezing, counting of fecal bolus, and the frequency of urination were quantified at the end of each trial to estimate fear and anxiety-related behavior in accordance with established procedure (Motz and Alberts 2005; Zimcikova et al. 2017; Owumi et al. 2022).
Preparation of sample for biochemical assays
Experimental animals were sacrificed following light ether anesthesia. The brains were carefully excised and parted into cerebellum and cerebrum under a cold condition (4 °C). Subsequently, the cerebellar and cerebral samples were homogenized in isolation using 50 mM Tris–HCl buffer of pH 7.4. The supernatant obtained after cold (4 °C) centrifugation at 12,000 g for 15 min was afterward used for biochemical assays using a SpectraMax plate reader (Molecular Devices, CA, USA). The protein concentration of the cerebellum and cerebrum samples was assessed, as previously reported (Bradford 1976).
Assessment of neurotoxicity and antioxidant status
Acetylcholinesterase (AChE) activity, an index of neurotoxicity, was analyzed by measuring the rate of DTNB conversion to thiolate dianion, as previously reported (Ellman et al. 1961). The antioxidant defense mechanism capability was evaluated using superoxide dismutase (SOD) and catalase (CAT) activities, according to Misra and Fridovich (Misra and Fridovich 1972) and Claiborne (Claiborne 1995), respectively, whereas glutathione peroxidase (GPx) and glutathione-S-transferase (GST) activities were analyzed in accordance with Rotruck et al. (1973) and Habig et al. (1974), respectively. The glutathione (GSH) and total thiol (TSH) levels were analyzed, as previously reported by Jollow et al. (1974) and Ellman (1959), respectively. Cellular oxidative status was assessed using xanthine oxidase (XO) activity as well as reactive oxygen and nitrogen species (RONS) and lipid peroxidation (LPO) levels in accordance with established methods (Adedara et al. 2016; Owumi et al. 2022).
Inflammatory and apoptotic indices
The cerebellar and cerebral levels of inflammatory stress and apoptosis were analyzed in the experimental animals. Myeloperoxidase (MPO) activity and nitric oxide (NO) level were assayed as inflammatory endpoints according to established protocol (Green et al. 1982; Granell et al. 2003), whereas caspase-9 and caspase-3 activities were spectrophotometrically evaluated as apoptotic enzyme endpoints using enzyme-linked immunosorbent assay kits.
Microscopic examination and histomorphometry
Histological evaluation and histomorphometry of the cerebral and cerebellar samples were carried out following fixing of samples with 10% phosphate-buffered formalin for 72 h by pathologists. The 5-μm sections of the samples pre-stained with hematoxylin and eosin were examined with a light microscope and image captured using a digital camera, as earlier reported (Bancroft and Gamble 2008). The degenerated Purkinje and cortical neurons in the brain sections were quantified following standard technique (Owoeye et al. 2018).
Statistics
Statistical analysis of the data was performed using one-way analysis of variance and Bonferroni’s post hoc test with Graphpad Prism 7 Software (Graph Pad Software Inc, USA). Statistical significance was set at values of p of less than 0.05.
Results
Outcome of exposure to coartem and AFB1 on neurobehavioral accomplishments
The locomotor, motor, and exploratory statuses of animals singly or jointly dosed with coartem and AFB1 in a novel environment are displayed in Figs. 1 and 2. Animals dosed with AFB1 alone exhibited enfeebled locomotor accomplishments, as shown by a significant reduction in the mobility time, maximum speed, and distance traveled, whereas animals dosed with COART alone exhibited no treatment-related effects on the locomotor accomplishments when compared with the control. Similarly, dosing of animals with AFB1 alone caused a marked decline in motor and turning fitness as typified by increased negative geotaxis and forelimb strength with reduced path efficiency (movement in a straight route), body rotation, and turn angle. Moreover, animals dosed with AFB1 alone exhibited increased anxiogenic behaviors, as shown by significant elevation in the frequency of urination, fecal bolus discharge and freezing time, whereas animals dosed with COART alone exhibited no treatment-related alterations in the motor and turning activities as well as anxiogenic behavior when compared with the control. Data from co-exposure to coartem and AFB1 showed that all the neurobehavioral parameters significantly improved and were similar to control in animals co-exposed to COART + AFB11 (35 µg/kg), whereas they were significantly decreased in animals dosed with COART + AFB12 (70 µg/kg) when compared with control and COART alone.

Influence of AFB1 and coartem on locomotor activity in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)

Influence of AFB1 and coartem co-exposure on motor activity and anxiety-like behavior in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)
In addition, the exploratory competence of animals singly or jointly dosed with coartem and AFB1 was verified by the track plots and heat maps which represent the roaming traces and cumulative period the animals spent in different areas of the novel environment, respectively (Fig. 3). Animals dosed with AFB1 alone exhibited disrupted exploratory behavior, as shown by diminished track plot density and greater heat map intensity, whereas animals dosed with COART alone exhibited no treatment-related alterations in exploratory behavior when compared with the control. Animals co-exposed to COART + AFB11 (35 µg/kg) exhibited considerable improvement in the exploratory competence comparable to control, whereas animals dosed with COART + AFB12 (70 µg/kg) exhibited noticeable reduction in the exploratory competence when compared with control and COART alone.

Influence of AFB1 and coartem co-exposure on exploratory behavior in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg respectively
Outcome of exposure to coartem and AFB1 on AChE activity and antioxidant status
The cerebral and cerebellar AChE activity and antioxidant status of animals singly or jointly dosed with coartem and AFB1 are displayed in Figs. 4 and 5. The AChE activity of animals dosed with coartem alone was not statistically different when compared with control, whereas AFB1 alone animals exhibited significant (p < 0.05) decreased in AChE activity, compared to control and coartem alone. Animals co-exposed to COART + AFB11 (35 µg/kg) exhibited significant restoration in the AChE activity similar to control, whereas animals dosed with COART + AFB12 (70 µg/kg) exhibited no improvement in the cerebral and cerebellar AChE activity when compared with control and COART alone. Furthermore, animals dosed with AFB1 alone exhibited significant impairment of the antioxidant enzymes, as shown by a significant reduction in GSH level together with GPx, GST, CAT, and SOD activities in the brain structures, whereas they were not affected in animals dosed with COART alone in comparison with control. Animals co-exposed to COART + AFB11 (35 µg/kg) demonstrated a significant improvement in the GSH level and activities of GPx, GST, CAT, and SOD in comparison with AFB1 alone. Animals dosed with COART + AFB12 (70 µg/kg) were mostly similar to AFB1 alone group and exhibited significant decrease in the antioxidant status when compared with control and COART alone.

Influence of AFB1 and coartem co-exposure on AChE, SOD, and CAT activities in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)

Influence of AFB1 and coartem co-exposure on GST and GPx activities cum GSH level in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg, respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)
Outcome of exposure to coartem and AFB1 on oxido-inflammatory stress
The activities of XO and MPO as well as the levels of NO, TSH, RONS, and LPO in the cerebrum and cerebellum of experimental animals singly or jointly dosed with coartem and AFB1 are shown in Figs. 6 and 7. Animals dosed with AFB1 alone exhibited a significant increase in inflammatory response typified by elevated MPO activity and NO level in addition to diminished TSH level and upsurge of oxidative stress indices namely XO, RONS, and LPO in the cerebrum and cerebellum when compared with control. Animals dosed with COART alone presented no treatment-related alterations in the oxidative and inflammatory stress indices when compared with the control. Animals co-exposed to COART + AFB11 (35 µg/kg) demonstrated significant improvement in the TSH level and activities of GPx, GST, CAT, and SOD in comparison with COART alone, whereas animals dosed with COART + AFB12 (70 µg/kg) exhibited a significant decrease in the antioxidant status when compared with control and COART alone. The significant reduction in the oxido-inflammatory stress indices was comparable to control in animals co-exposed to COART + AFB11 (35 µg/kg), whereas they were significantly elevated in animals co-exposed to COART + AFB12 (70 µg/kg) in comparison with control and COART alone.

Influence of AFB1 and coartem co-exposure on XO and MPO activities cum NO level in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg, respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)

Influence of AFB1 and coartem co-exposure on TSH, RONS, and LPO levels in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg, respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)
Outcome of exposure to coartem and AFB1 on caspase activation
The cerebral and cerebellar caspase-9 and caspase-3 activities in animals singly or jointly dosed with coartem and AFB1 are displayed in Fig. 8. Animals dosed with AFB1 alone exhibited a significant increase in apoptotic indices, specifically caspase-9 and caspase-3 activities, whereas animals dosed with COART alone exhibited no treatment-related changes in caspase-9 and caspase-3 activities when compared with the control. The cerebral and cerebellar activities of caspase-9 and caspase-3 were significantly abated in both groups of animals co-exposed to COART + AFB1, compared with control but were significantly higher than the control.

Influence of AFB1 and coartem co-exposure on caspase-9 and caspase-3 activities in rats. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg, respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)
Outcome of exposure to coartem and AFB1 on brain histology and histomorphometry
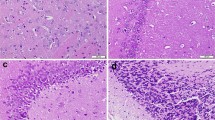
Representative photomicrographs and histomorphometric data of the brain structures of animals singly or jointly exposed to coartem and AFB1 are shown in Figs. 9 and 10. Animals dosed with COART alone exhibited no treatment-related changes in the histological appearance and histomorphometry of the cerebellum and cerebrum compared with control. Conversely, animals dosed with AFB1 alone exhibited marked neuronal degeneration associated with pyknosis, congestion, and vacuolation of the neuropil in the cerebral cortex, whereas Purkinje cell layer neuronal degeneration in the cerebellum was associated with diminished nuclear material. Histological injuries were significantly abated in both groups of animals co-exposed to COART + AFB1 when compared with control. However, neuronal degeneration in animals co-exposed to COART + AFB12 (70 µg/kg) were significantly higher than the control and COART alone.

Histological and histomorphometric data of the cerebrum. Sections from the control and coartem alone appear normal. AFB1 alone-dosed section showing neuronal degeneration associated with pyknosis, congestion, and vacuolation of the neuropil (green arrows). Coartem minimized neuronal degeneration in animals co-exposed to low AFB1 dose than high dose. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg, respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)

Histological and histomorphometric data of the cerebellum. Sections from the control and coartem alone appear normal. Animals dosed with AFB1 alone showing neuronal degeneration associated with reduced nuclear material (red arrows) in the Purkinje cell layer. Coartem minimized neuronal degeneration in animals co-exposed to low AFB1 dose than high dose. AFB1 indicates aflatoxin B1. AFB1 denotes 70 µg/kg; AFB11 and AFB12 denote 35 and 70 µg/kg, respectively. COART denotes coartem at 5 mg/kg respectively. Each bar represents mean ± S.D. for 8 rats per group. a: Values differ significantly from control (p < 0.05). b: Values differ significantly from AFB1 alone (p < 0.05). c: Values differ significantly from COART and AFB11 (p < 0.05). d: Values differ significantly from COART and AFB11 (p < 0.05)
Discussion
The present investigation is the first report on the impact of co-exposure to anti-malarial drug coartem and AFB1, a dietary contaminant, on the behavioral and neurochemical endpoints in rats. The data herein showed the mitigating effect of coartem administration on impaired motor and turning fitness parameters (i.e., body rotation, turn angle, and grip strength) in animals exposed to low dose of AFB1 but failed in animals that were dosed with high dose of AFB1. The adverse effect of AFB1 on motor and locomotor fitness has been reported (Bahey et al. 2015; Wu et al. 2019). The significant improvement in motor fitness parameters connotes the efficiency of coartem administration to modify the deficits in the coordination between the muscle and the nervous system in low dose AFB1-intoxicated animals. Furthermore, coartem administration markedly attenuated locomotor insufficiencies characterized by diminished mobility time, maximum speed, distance traveled, and straight path movement in rats exposed to low dose of AFB1. The persistent decrease in locomotor parameters in animals treated with high AFB1 dose clearly connotes the failure of coartem administration to modify the locomotor deficits elicited by this high dose of AFB1.
Moreover, our findings indicate that coartem administration partially abated anxiogenic behaviors such as fecal bolus discharge, frequency of urination, geotaxis, and freezing in animals that were dosed with AFB1. This clearly indicates the inability of coartem to protect against induction of fear and anxiety caused by these investigated doses of AFB1. Previous investigators have reported that AFB1 induces anxiety and depression-like behaviors in rats (Aytekin Sahin et al. 2022). Exploratory behavior is an important survival skill by which organism gains information about its spatial environments (Gorny et al. 2002; Bingman and Sharp 2006). The obvious improvement in track plot densities with reduced heat map intensity in animals administered coartem, and low dose of AFB1 indicates the effectiveness of coartem to modify the exploratory and spatial behavior deficits in the animals. However, coartem failed to protect against exploratory behavior deficits associated with high dose of AFB1.
The current investigation further unravels the impact of co-exposure to AFB1 and coartem on the cholinergic system, antioxidant defense mechanisms, inflammatory stress, and apoptotic response in the cerebrum and cerebellum of exposed animals. The cholinergic nerve cells utilize acetylcholine during neurotransmission to generate and control locomotor activity in vertebrate species (Le Ray et al. 2022). The hydrolysis of acetylcholine to choline and acetate by AChE is essential for the termination of synaptic neurotransmission. The current finding on AFB1-mediated suppression of cholinergic signaling corroborates previous studies (Tanaka et al. 2015; Adedara et al. 2021). The possible beneficial role of coartem in cholinergic neurotransmission was observed in rats treated with low AFB1 dose. However, the persistent suppression of AChE activity in high AFB1-dosed animals may elicit neuronal dysfunction as a result of acetylcholine buildup at the synaptic nerve which reportedly is associated with downregulation and desensitization of cholinergic receptors (van Koppen and Kaiser 2003). This finding further indicates the unbeneficial effect of coartem on impaired cholinergic function, and consequently, the behavioral deficits associated with exposure to high AFB1 dose.
Antioxidant defense mechanism is fundamental for the maintenance of cellular integrity because it directly and/or indirectly removes RONS and suppresses cellular oxidative injury (Cenini et al. 2019). Here, the diminutions in the enzymatic and non-enzymatic antioxidants persisted along with elevated indices of oxidative stress in the cerebrum and cerebellum of animals co-exposed to coartem and both doses of AFB1. The upsurge in the lipid peroxidation level in brains of AFB11-treated rats corroborates earlier findings (Aytekin Sahin et al. 2022). Animals in coartem and AFB1 exhibited lesser oxidative damage than animals exposed to AFB1 alone. However, these findings clearly indicate unprofitable effect of coartem on dysfunctional antioxidant defense mechanisms which is associated with RONS buildup and subsequently cellular oxidative damage in the AFB1-exposed animals.
Moreover, we investigated the impact of coartem and AFB1 co-exposure on inflammatory and apoptotic response in the cerebrum and cerebellum of experimental animals. Low physiological NO concentration is essential for the nervous system function and immunity, whereas elevated NO level mediates inflammatory response and induction of nitrosative stress leading to protein, nucleic acids, and lipid modification (Kapil et al. 2020). The MPO exhibits cytokine-related activity and is involved in neutrophil stimulation to produce ROS and inflammatory response (Lau et al. 2005). The upsurge in NO level and MPO activity as well as caspase-9 and caspase-3 activities persisted in animals co-exposed to coartem and both doses of AFB1. The activation of initiator caspase-9 and executioner caspase-3 is essential in the intrinsic pathway of apoptotic cell death (Owumi et al. 2022; Unnisa et al. 2022). Although animals in the co-exposure group exhibited lesser oxidative damage than animals in the AFB1 alone, the marked increases in these inflammatory and apoptotic indices evidently indicate unbeneficial effect of coartem on inflammation and cell death associated with AFB1 exposure. Moreover, histopathological and mophormetrical findings corroborate biochemical and behavioral findings that coartem administration was ineffective in mitigating neurotoxicity associated with high dose of AFB1 exposure. The histopathological injury observed in brains of AFB11-treated rats in this study is consistent with earlier findings (Aytekin Sahin et al. 2022).
In conclusion, the current data demonstrated that co-exposure to AFB1 and coartem posed no additional risk on the behavioral and biochemical responses when compared with separate exposure to AFB1. Coartem partially lessened neurotoxicity associated with AFB1 exposure in rats. Thus, the scenario of co-exposure to coartem and AFB1-contaminated diet may not worsen the health condition for such individual undergoing malaria treatment.
Data availability
The original data and materials of the current study are available with the corresponding author and would be made available on justifiable request.
References
Adedara IA, Abolaji AO, Awogbindin IO, Farombi EO (2017a) Suppression of the brain-pituitary-testicular axis function following acute arsenic and manganese co-exposure and withdrawal in rats. J Trace Elem Med Biol 39:21–29
Adedara IA, Abolaji AO, Idris UF, Olabiyi BF, Onibiyo EM, Ojuade TD, Farombi EO (2017b) Neuroprotective influence of taurine on fluoride-induced biochemical and behavioral deficits in rats. Chem Biol Interact 261:1–10
Adedara IA, Abolaji AO, Rocha JB, Farombi EO (2016) Diphenyl diselenide protects against mortality, locomotor deficits and oxidative stress in Drosophila melanogaster model of manganese-induced neurotoxicity. Neurochem Res 41:1430–1438
Adedara IA, Fasina OB, Ayeni MF, Ajayi OM, Farombi EO (2019) Protocatechuic acid ameliorates neurobehavioral deficits via suppression of oxidative damage, inflammation, caspase-3 and acetylcholinesterase activities in diabetic rats. Food Chem Toxicol 125:170–181
Adedara IA, Nanjappa MK, Farombi EO, Akingbemi BT (2014) Aflatoxin B1 disrupts the androgen biosynthetic pathway in rat Leydig cells. Food Chem Toxicol 65:252–259
Adedara IA, Owumi SE, Oyelere AK, Farombi EO (2021) Neuroprotective role of gallic acid in aflatoxin B1-induced behavioral abnormalities in rats. J Biochem Mol Toxicol 35:e22684. https://doi.org/10.1002/jbt.22684
Allen SJ, Wild CP, Wheeler JG, Riley EM, Montesano R, Bennett S, Whittle HC, Hall AJ, Greenwood BM (1992) Aflatoxin exposure, malaria and hepatitis B infection in rural Gambian children. Trans R Soc Trop Med Hyg 86:426–430
Aytekin Sahin G, Karabulut D, Unal G, Sayan M, Sahin H (2022) Effects of probiotic supplementation on very low dose AFB1-induced neurotoxicity in adult male rats. Life Sci 306:120798. https://doi.org/10.1016/j.lfs.2022.120798
Bahey NG, Abd Elaziz HO, Gadalla KK (2015) Toxic effect of aflatoxin B1 and the role of recovery on the rat cerebral cortex and hippocampus. Tissue Cell 47:559–566
Baldissera MD, Souza CF, Zeppenfeld CC, Descovi SN, Moreira KLS, da Rocha MIUM, da Veiga ML, da Silva AS, Baldisserotto B (2018) Aflatoxin B1-contaminated diet disrupts the blood-brain barrier and affects fish behavior: involvement of neurotransmitters in brain synaptosomes. Environ Toxicol Pharmacol 60:45–51
Balikagala B, Fukuda N, Ikeda M, Katuro OT, Tachibana SI, Yamauchi M, Opio W, Emoto S, Anywar DA, Kimura E, Palacpac NMQ, Odongo-Aginya EI, Ogwang M, Horii T, Mita T (2021) Evidence of artemisinin-resistant malaria in Africa. N Engl J Med 385(13):1163–1171
Bancroft JD, Gamble M (2008) Theory and practice of histology techniques, 6th ed. Churchill Livingstone Elsevier, pp. 83–134
Benkerroum N (2020) Aflatoxins: producing-molds, structure, health issues and incidence in Southeast Asian and Sub-Saharan African countries. Int J Environ Res Public Health 17:1215
Bingman VP, Sharp PE (2006) Neuronal implementation of hippocampal-mediated spatial behavior: a comparative evolutionary perspective. Behav Cogn Neurosci Rev 5:80–91
Bradford MM (1976) Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Cenini G, Lloret A, Cascella R (2019) Oxidative stress in neurodegenerative diseases: from a mitochondrial point of view. Oxid Med Cell Longev 2019:2105607. https://doi.org/10.1155/2019/2105607
Claiborne A (1995) Catalase activity. In: Greewald AR (ed) Handbook of methods for oxygen radical research. CRC Press, Boca Raton, pp 237–242
Classen W, Altmann B, Gretener P, Souppart C, Skelton-Stroud P, Krinke G (1999) Differential effects of orally versus parenterally administered qinghaosu derivatives artemether in dogs. Exp Toxicol Pathol 51:507–516
El-Dakdoky MH (2009) Evaluation of the developmental toxicity of artemether during different phases of rat pregnancy. Food Chem Toxicol 47:1437–1441
Ellman GL, Courtney KD, Andres V Jr, Feather-Stone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7:88–95
Ellman GL (1959) Tissue sulfhydryl groups. Arch Biochem Biophys 82:70–77
Ferrari L, Fumagalli F, Rizzi N, Grandi E, Vailati S, Manoni M, Ottoboni M, Cheli F, Pinotti L (2022) An eight-year survey on aflatoxin B1 indicates high feed safety in animal feed and forages in Northern Italy. Toxins (basel) 14(11):763. https://doi.org/10.3390/toxins14110763
Folarin O, Olopade F, Onwuka S, Olopade J (2016) Memory deficit recovery after chronic vanadium exposure in mice. Oxid Med Cell Longev 2016:4860582. https://doi.org/10.1155/2016/4860582
Gautam A, Ahmed T, Batra V, Paliwal J (2009) Pharmacokinetics and pharmacodynamics of endoperoxide antimalarials. Curr Drug Metab 10:289–306
Gorny JH, Gorny B, Wallace DG, Whishaw IQ (2002) Fimbria-fornix lesions disrupt the dead reckoning (homing) component of exploratory behavior in mice. Learn Mem 9:387–394
Granell S, Gironella M, Bulbena O, Panés J, Mauri M, Sabater L, Aparisi L, Gelpí E, Closa D (2003) Heparin mobilizes xanthine oxidase and induces lung inflammation in acute pancreatitis. Crit Care Med 31:525–530
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR (1982) Analysis of nitrate, nitrite and [15N]nitrate in biological fluids. Anal Biochem 126:131–138
Habig WH, Pabst MJ, Jakoby WB (1974) Glutathione S-transferase. The first enzymatic step in mercapturic acid formation. J Biol Chem 249:7130–7139
Hernandez Maldonado J, Grundmann O (2022) Drug-drug interactions of artemisinin-based combination therapies in malaria treatment: a narrative review of the literature. J Clin Pharmacol 62:1197–1205
Ho WE, Peh HY, Chan TK, Wong WS (2014) Artemisinins: pharmacological actions beyond anti-malarial. Pharmacol Ther 142:126–139
International Agency for Research on Cancer (2000) Monographs on aflatoxins, vol 82. IARC Press, Lyon, pp 171–300
Jollow DJ, Mitchell JR, Zampaglione N, Gillette JR (1974) Bromobenzene induced liver necrosis: protective role of glutathione and evidence for 3,4 bromobenzene oxide as the hepatotoxic metabolite. Pharmacology 11:151–169
Kamchonwongpaisan S, McKeever P, Hossler P, Ziffer H, Meshnick SR (1997) Artemisinin neurotoxicity: neuropathology in rats and mechanistic studies in vitro. Am J Trop Med Hyg 56:7–12
Kapil V, Khambata RS, Jones DA, Rathod K, Primus C, Massimo G, Fukuto JM, Ahluwalia A (2020) The noncanonical pathway for in vivo nitric oxide generation: the nitrate-nitrite-nitric oxide pathway. Pharmacol Rev 72:692–766
Karri V, Schuhmacher M, Kumar V (2016) Heavy metals (Pb, Cd, As and MeHg) as risk factors for cognitive dysfunction: a general review of metal mixture mechanism in brain. Environ Toxicol Pharmacol 48:203–213
Lau D, Mollnau H, Eiserich JP, Freeman BA, Daiber A, Gehling UM, Brümmer J, Rudolph V, Münzel T, Heitzer T, Meinertz T, Baldus S (2005) Myeloperoxidase mediates neutrophil activation by association with CD11b/CD18 integrins. Proc Natl Acad Sci USA 102:431–436
Le Ray D, Bertrand SS, Dubuc R (2022) Cholinergic modulation of locomotor circuits in vertebrates. Int J Mol Sci 23:10738. https://doi.org/10.3390/ijms231810738
Li Q, Hickman M (2011) Toxicokinetic and toxicodynamic (TK/TD) evaluation to determine and predict the neurotoxicity of artemisinins. Toxicology 279(1–3):1–9
Marin S, Ramos AJ, Cano-Sancho G, Sanchis V (2013) Mycotoxins: occurrence, toxicology, and exposure assessment. Food Chem Toxicol 60:218–237
Meng Y, Ma N, Lyu H, Wong YK, Zhang X, Zhu Y, Gao P, Sun P, Song Y, Lin L, Wang J (2021) Recent pharmacological advances in the repurposing of artemisinin drugs. Med Res Rev 41:3156–3181
Misra HP, Fridovich I (1972) The role of superoxide anion in the autooxidation of epinephrine and a simple assay for superoxide dismutase. J Biol Chem 247:3170–3175
Motz BA, Alberts JR (2005) The validity and utility of geotaxis in young rodents. Neurotoxicol Teratol 27:529–533
Nuwagira C, Peter EL, Ajayi CO, Adriko J, Kagoro GR, Olet EA, Ogwang PE, Tolo CU (2021) Developmental stages influence in vivo antimalarial activity of aerial part extracts of Schkuhria pinnata. J Ethnopharmacol 279:114341. https://doi.org/10.1016/j.jep.2021.114341
Owoeye O, Adedara IA, Farombi EO (2018) Pretreatment with taurine prevented brain injury and exploratory behaviour associated with administration of anticancer drug cisplatin in rats. Biomed Pharmacother 102:375–384
Owumi SE, Adedara IA, Oyelere AK (2022) Indole-3-propionic acid mitigates chlorpyrifos-mediated neurotoxicity by modulating cholinergic and redox-regulatory systems, inflammatory stress, apoptotic responses and DNA damage in rats, Environ. Toxicol Pharmacol 89:103786. https://doi.org/10.1016/j.etap.2021.103786
Owumi SE, Gbadegesin MA, Odunola OA, Adegoke AM, Uwaifo AO (2015) Toxicity associated with repeated administration of artemether-lumefantrine in rats. Environ Toxicol 30:301–307
Pernaute-Lau L, Camara M, Nóbrega de Sousa T, Morris U, Ferreira MU, Gil JP (2022) An update on pharmacogenetic factors influencing the metabolism and toxicity of artemisinin-based combination therapy in the treatment of malaria. Expert Opin Drug Metab Toxicol 18:39–59
Petras JM, Young GD, Bauman RA, Kyle DE, Gettayacamin M, Webster HK, Corcoran KD, Peggins JO, Vane MA, Brewer TG (2000) Brewer, Arteether-induced brain injury in Macaca mulatta I. The precerebellar nuclei: the lateral reticular nuclei, paramedian reticular nuclei, and perihypoglossal nuclei. Anat Embryol (berl) 201:383–397
Qureshi H, Hamid SS, Ali SS, Anwar J, Siddiqui AA, Khan NA (2015) Cytotoxic effects of aflatoxin B1 on human brain microvascular endothelial cells of the blood-brain barrier. Med Mycol 53:409–416
Ramos-Martín V, González-Martínez C, Mackenzie I, Schmutzhard J, Pace C, Lalloo DG, Terlouw DJ (2014) Neuroauditory toxicity of artemisinin combination therapies-have safety concerns been addressed? Am J Trop Med Hyg 91:62–73
Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, Hoekstra WG (1973) Selenium: biochemical role as a component of glutathione peroxidase. Science 179:588–590
Rushing BR, Selim MI (2019) Aflatoxin B1: a review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem Toxicol 124:81–100
Skelton-Stroud P, Mull R (1998) Positioning, labelling, and medical information control of co-artemether tablets (CPG 56697): a fixed novel combination of artemether and benflumetol. Novartis Co-Artemether International Development Team. Med Trop (mars) 58:77–81
Souto NS, Claudia Monteiro Braga A, Lutchemeyer de Freitas M, Rechia Fighera M, Royes LFF, Schneider Oliveira M, Furian AF (2018) Aflatoxin B1 reduces non-enzymatic antioxidant defenses and increases protein kinase C activation in the cerebral cortex of young rats. Nutr Neurosci 21:268–275
Sullivan AP (2022) Mycotoxin illness: recognition and management from functional medicine perspective. Phys Med Rehabil Clin N Am 33:647–663
Tanaka T, Mizukami S, Hasegawa-Baba Y, Onda N, Sugita-Konishi Y, Yoshida T, Shibutani M (2015) Developmental exposure of aflatoxin B1 reversibly affects hippocampal neurogenesis targeting late-stage neural progenitor cells through suppression of cholinergic signaling in rats. Toxicology 336:59–69
Unnisa A, Greig NH, Kamal MA (2022) Inhibition of caspase 3 and caspase 9 mediated apoptosis: a multimodal therapeutic target in traumatic brain injury. Curr Neuropharmacol 27. https://doi.org/10.2174/1570159X20666220327222921
van Koppen CJ, Kaiser B (2003) Regulation of muscarinic acetylcholine receptor signaling. Pharmacol Ther 98:197–220
Wagacha JM, Muthomi JW (2008) Mycotoxin problem in Africa: current status, implications to food safety and health and possible management strategies. Int J Food Microbiol 124:1–12
World Health Organization (2020) World Malaria Report. www.who.int/publications/i/item/9789240015791
Wu TS, Cheng YC, Chen PJ, Huang YT, Yu FY, Liu BH (2019) Exposure to aflatoxin B1 interferes with locomotion and neural development in zebrafish embryos and larvae. Chemosphere 217:905–913
Zhang T, Zhang X, Lin C, Wu S, Wang F, Wang H, Wang Y, Peng Y, Hutchinson MR, Li H, Wang X (2021) Artemisinin inhibits TLR4 signaling by targeting co-receptor MD2 in microglial BV-2 cells and prevents lipopolysaccharide-induced blood-brain barrier leakage in mice. J Neurochem 157:611–623
Zimcikova E, Simko J, Karesova I, Kremlacek J, Malakova J (2017) Behavioral effects of antiepileptic drugs in rats: are the effects on mood and behavior detectable in open-field test? Seizure 52:35–40
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Adedara, I.A., Owumi, S.E. Neurobehavioral and biochemical responses to artemisinin-based drug and aflatoxin B1 co-exposure in rats. Mycotoxin Res 39, 67–80 (2023). https://doi.org/10.1007/s12550-023-00474-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12550-023-00474-6