
Abstract
The rosetted trace fossil Dactyloidites ottoi (Geinitz, 1849) is discussed based on new occurrences from the Cenomanian Regensburg Formation (glauconitic sandstones, Bavaria) and on a careful re-description of the type material and other specimens from quartz-rich sandstones of the Cenomanian Oberhäslich Formation of Saxony. The feeding trace consists of a fan-shaped spreiten structure originating from a central, vertical to oblique shaft leading downwards into the sediment. The branching radial elements (up to 20, 4–6 mm wide) are subhorizontal protrusive vertical spreiten, mostly forming incompletely circular rosettes with radii of 200°–270° and diameters between 30 and 75 mm. The ichnotaxonomic classification of the trace has been discussed controversially. Gyrophyllites kwassizensis Glocker, 1841 from contemporaneous nearshore glauconitic sandstones in the Bohemian Cretaceous Basin is similar to D. ottoi, but its radial elements are club-shaped and unbranched and do not show any spreiten. The validity of the ichnogenus Haentzschelinia Vyalov, 1964 (type ichnospecies Spongia ottoi) is questioned because the essential behaviour expressed by this form is basically the same as in Dactyloidites Hall, 1886. Different size, age, depositional environment, shape of the rosettes (circular vs. fan-shaped) and number of radial elements are not valid criteria for an ichnogeneric separation. Thus, the classification of S. ottoi in Dactyloidites Hall, 1886 is reasonable. The potential tracemaker of D. ottoi was a worm-like organism systematically reworking the sediment for organic food particles, supported by studies on the feeding behaviour of the modern lugworm Arenicola marina: when nutrient-rich sediments occur at living depth, the lugworm directly ingests the surrounding sediment by generating radial tunnels originating from the central shaft. This feeding strategy results in rosetted structures very similar to D. ottoi. The ichnospecies has a proven range from the Jurassic to the Neogene and predominantly occurs in shallow-water, nearshore to deltaic, nutrient-rich siliciclastic settings (lower Skolithos and upper Cruziana ichnofacies). The narrow palaeoenvironmental window in which the trace may form (shallow-marine settings, local presence of organic-rich sediments at living depth) is furthermore closed by taphonomic constraints: high sedimentation rates towards the top of shallowing-upward cycles support the preservation of D. ottoi, while low accumulation rates during transgressive conditions enhance the chances of their destruction by subsequent bioturbation. Thus, both the formation and the preservation of D. ottoi are related to narrow palaeoenvironmental and specific taphonomic conditions, readily explaining the rarity of the ichnotaxon.
Kurzfassung
Das rosettenförmige Spurenfossil Dactyloidites ottoi (Geinitz, 1849) wird auf Grund neuer Funde aus glaukonitischen Sandsteinen der Regensburg-Formation (Cenoman, Bayern) und einer sorgfältigen Neubeschreibung des Typusmaterials sowie weiterer Exemplare aus flachmarinen Quarzsandsteinen der Oberhäslich-Formation (Cenoman, Sachsen) diskutiert. Die Fressspur besteht aus fächerförmigen Spreitenstrukturen, die von einem zentralen, vertikal bis schräg ins Sediment herabführenden Schacht radial abzweigen. Diese radialen, sich verzweigenden Elemente (bis zu 20 an der Zahl, 4–6 mm breit) bestehen aus subhorizontalen, protrusiven vertikalen Spreiten, die zumeist unvollständige Rosetten mit Radien von etwa 200°–270° und Durchmessern von 30–75 mm bilden. Die ichnotaxonomische Klassifikation der Spur wird kontrovers diskutiert. Gyrophyllites kwassizensis Glocker, 1841 aus zeitgleichen glaukonitischen Sandsteinen des Böhmischen Kreidebeckens ist ähnlich, die radialen Elemente sind allerdings keulenförmig, unverzweigt und zeigen keine Spreiten. Die Gültigkeit der Spurengattung Haentzschelinia Vyalov, 1964 (Typusart Spongia ottoi) wird angezweifelt, da das in ihr ausgedrückte Verhaltensmuster im Grunde identisch ist mit dem der dann prioritären Spurengattung Dactyloidites Hall, 1886. Unterschiedliche Größe, Alter und Ablagerungsverhältnisse begründen ebenso wenig eine Trennung in zwei Spurengattungen wie die Form der Rosetten (kreisförmig vs. fächerförmig) und die Zahl der radialen Elemente. Deshalb wird die Einordnung von Spongia ottoi in Dactyloidites Hall, 1886 hier unterstützt. Potentielle Erzeuger von D. ottoi waren wurmartige Tiere, die systematisch das Sediment nach verwertbaren Nahrungspartikeln durchwühlt haben. Unterstützt wird diese Hypothese durch Studien des Fressverhaltens des rezenten Wattwurms Arenicola marina: wenn genügend Nahrung in der Sedimentschicht der Wohntiefe auftritt, verzichtet das Tier auf die Anlage des normalen U-förmigen Ganges und produziert radiale Fressgänge, die vom zentralen Schacht horizontal abgehen. So entstehen rosettenförmige Spuren, die den fossilen D. ottoi sehr ähnlich sind. D. ottoi hat eine gesicherte Reichweite vom Jura bis ins Neogen und wurde überwiegend in küstennahen, nährstoffreichen, siliziklastischen Flachwassermilieus nachgewiesen (untere Skolithos- und obere Cruziana-Ichnofazies). Das enge paläoökologische Fenster, in dem die Spur enstehen kann (Flachwasser-Bedingungen, Verfügbarkeit von Nährstoffpartikel-reichen Sedimenten in der Wohntiefe des Erzeugers), wird durch taphonomische Gründe weiter eingeengt: hohe Sedimentationsraten zum Top von Verflachungszyklen können die Erhaltung der Spur begünstigen, wohingegen geringe Sedimentationsraten und transgressive Ablagerungsbedingungen sich negativ auf die Überlieferung von D. ottoi auswirken. Somit sind sowohl die Bildung als auch die Erhaltung von D. ottoi an sehr spezifische Bedingungen geknüpft, weshalb die Seltenheit der Spur im stratigraphischen Bericht nicht weiter verwundert.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rosetted trace fossils are common throughout the geological record and have been recorded from deep-water flysch to deltaic deposits. Dactyloidites ottoi (Geinitz, 1849), however, is a relatively rare form, known from (?Triassic–) Jurassic to Neogene, mostly shallow-marine and deltaic siliciclastic environments (Fürsich and Bromley 1985; Agirrezabala and de Gibert 2004).
Dactyloidites ottoi was first described by Geinitz (1849) as Spongia ottoi and interpreted as a sponge. Other authors interpreted the rosetted structure as plant remains or medusoid body fossils. Although Morin (1907) compared D. ottoi with fiddler crab feeding traces, it was not before Häntzschel (1930) that the structure slowly became accepted as trace fossil. Fürsich and Bromley (1985) described well-preserved material from the Upper Cretaceous of Greenland that allowed them to interpret the trace as a feeding structure (fodinichnion), composed of multiply branched radial elements consisting of protrusive vertical spreiten.
The scope of the present contribution is a careful re-description of the type material of S. ottoi Geinitz, 1849 and of additional specimens from sandstones of the Cenomanian Oberhäslich Formation in Saxony (eastern Germany), and a comparison with new occurrences of D. ottoi from the Cenomanian Regensburg Formation of Bavaria (southern Germany). All material is from shallow-marine sandstones of inner shelf environments, and thus, the palaeoenvironmental significance of the trace fossil and its potential producer is discussed as well. Furthermore, ichnotaxonomic issues concerning the ichnogenera Gyrophyllites Glocker, 1841 and Haentzschelinia Vyalov, 1964 are addressed.
Sections
Oberhäslich Formation of the Elbtal Group, Saxony
In Saxony, the area between Meißen and the Czech border is characterized by sedimentary rocks of early Late Cretaceous (Cenomanian–Turonian) age, more or less following the River Elbe Valley. The lithofacies of these Cretaceous strata is dominated by marine siliciclastics and marls or marly limestones (so-called Pläner) that lithostratigraphically have been combined in the Elbtal Group (Tröger and Voigt in Niebuhr et al. 2007). The Elbtal Group forms an important link between the temperate Boreal shelf of northern and northwestern Europe and the Tethyan warm-water areas to the south (Fig. 1). It has been deposited in a relatively narrow strait between the Westsudetic Island and the Bohemian Massif (Osterzgebirge) which acted as source of siliciclastic sediments. For a geological overview of the Elbtal Group, see Tröger (2003).

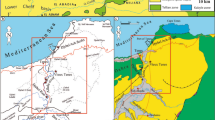
Cenomanian palaeogeographic map showing the position of the two occurrences of Dactyloidites ottoi discussed herein. Map modified after Philip and Floquet (2000)
The first marine transgression recorded in the Elbtal Group came from the north and took place in the late Early Cenomanian, reaching the area of Meißen (conglomeratic bioclastic limestones of the Meißen Formation). The major transgression, however, occurred during the early Late Cenomanian (Calycoceras naviculare Zone) when shallow marine siliciclastics onlapped far southward onto the formerly emergent Osterzgebirge. These conglomerates, sand- and siltstones constitute the Oberhäslich Formation (Tröger and Voigt in Niebuhr et al. 2007). It is usually 10–15 m thick but may reach up to 30 m based on the pre-transgression topography. The predominant lithofacies are compositionally and texturally mature, fine- to medium-grained quartz sandstones. Macrofossils include numerous diverse bivalves, associated with siliceous sponges and echinoids as well as rare starfish and ammonites (Wilmsen and Richardt 2012; Wilmsen and Nagm 2013). Bioturbation is pervasive, with Thalassinoides saxonicus (Geinitz) and Ophiomorpha nodosa Lundgren being common ichnotaxa. D. ottoi, however, is very rare and has only been recorded from the area of Dippoldiswalde (Geinitz 1849, 1871; Häntzschel 1930) and the Tharandter Wald (Göhler 2011). Complete sections do not exist in that area, thus a composite log of the Oberhäslich Formation is displayed in Fig. 2. However, the exact horizon of the type material of D. ottoi is unknown. It is only known that it stems from a single layer at the top of the “Unterquader” (Oberhäslich Formation) at Wendischcarsdorf, 5 km north of Dippoldiswalde (Otto 1852).

Stratigraphic logs of the Oberhäslich and Regensburg formations with inferred levels of Dactyloidites ottoi. The strongly differing lithology of the lower and upper freestone bed in the Ihrlerstein Quarry is illustrated with two thin-section photomicrographs
Regensburg Formation of the Danubian Cretaceous Group, Bavaria
The Regensburg Formation of the type area between Regensburg and Kelheim (Bavaria, southern Germany) is of Early to mid-Late Cenomanian age and has thickness of up to 16 m (Niebuhr et al. 2009). It is a transgressive unit that has been deposited in inner to mid-shelf settings of the north Tethyan margin, close to the emergent Rheno-Bohemian Massif (Fig. 1; see Wilmsen and Niebuhr 2010; Wilmsen et al. 2010; Richardt et al. 2013 for details). The lower part of the formation, the Saal Member (Lower Cenomanian), yielded famous freestones for buildings, known under the names “Regensburger Grünsandstein”, “Ihrlersteiner Grünsandstein” or “Grüner Donausandstein”. Although the freestone has been quarried at numerous sites since more than 400 years ago, only the glauconitic sandstones from the Ihrlerstein Quarry yielded the rosetted trace fossil D. ottoi, as well seen in buildings made of the Ihrlersteiner Grünsandstein, e.g. the stairs of the Befreiungshalle at Kelheim (Rieger and Rieger 1954; Rutte 1982; Endlicher 1984; Hafner 1998) or the flood masonry walls and the bridge arches of the Steinerne Brücke of Regensburg (see Meyer and Schmidt-Kaler 1995). However, even though the Alte Pinakothek, the Glyptothek and the Residenz in Munich are also built of Ihrlersteiner Grünsandstein, D. ottoi is not known from these buildings.
The Ihrlerstein Quarry is located at the western margin of Ihrlerstein, ca. 3 km north of Kelheim and the River Donau. It was opened in 1825 (Hafner 1998) and shows the Regensburg Formation resting on Upper Jurassic “Kelheimer Kalke” and Lower Cretaceous clays and sands of the terrestrial Schutzfels Formation in karst depressions. In 2008, boreholes were drilled in the direct vicinity of the quarry and geochemical analyses were carried out (boreholes Ihrlerstein TRACO B1–B3 of the Bayerisches Landesamt für Umwelt). In 2011, the quarry section was logged in detail by the authors (Fig. 2).
The lower ca. 9 m of the Regensburg Formation in the Ihrlerstein Quarry belong to the Saal Member and yield two levels of freestones (lower and upper Werksandstein). The lower, ca. 3.50-m-thick freestone is a fine- to coarse-grained calcareous sandstone of dark-greenish colour with high glauconite contents and many scattered shell remains (mainly of inoceramids and oysters; Fig. 2). Thin-section analysis showed a quartzose, fine- to coarse-grained bioclastic sandstone with bimodal grain-size distribution and 10–15 % allochthonous glauconite; shell remains up to 50 mm (inoceramid bivalves, oysters, serpulids) occur commonly. Correlation to the Bad Abbach section, about 14 km in the east (see Niebuhr et al. 2009, 20) indicates an Early Cenomanian age for this bed at Ihrlerstein. The upper freestone at the 4–7 m level is subdivided into two fine-grained, less glauconitic sandstones by a soft, argillaceous layer at 5.50 m. It is yellow to light-greenish in colour and has a homogeneous fabric. Thin sections show fine-grained, calcareous quartz sandstone with 5–10 % glauconite and only rare, small bioclasts (Fig. 2). Due to the colour and fabric, the D. ottoi on the stairs of the Befreiungshalle are inferred to stem from the upper freestone bed of Ihrlerstein. The upper part of the Saal Member (6.90–8 m level) is again rich in glauconite. At ca. 8 m, a conspicuous change in lithofacies to the Middle to Upper Cenomanian Bad Abbach Member occurs. The alternation of soft silty marls and nodular marly–silty limestones shows yellow to brownish colours. Referring to the above-mentioned boreholes, the Regensburg Formation at Ihrlerstein has thickness of ca. 15 m, and is followed by the ca. 6-m-thick marly Eibrunn Formation (mid-Upper Cenomanian to lowermost Turonian; see Niebuhr et al. 2009).
Systematic ichnology
Materials and methods
The Saxonian material is housed in the collections of the Senckenberg Naturhistorische Sammlungen Dresden, Museum für Mineralogie und Geologie (MMG, repository SaK). The Bavarian material was studied in the field (stairs of the Befreiungshalle, Kelheim). Measurements were obtained using a sliding calliper.
Ichnogenus Dactyloidites Hall, 1886
Type ichnospecies:
Dactyloidites asteroides (Fitch, 1850).
Dactyloidites ottoi (Geinitz, 1849)

a Holotype of Dactyloidites ottoi (Geinitz, 1849) from the Oberhäslich Formation (Saxony), Wendischcarsdorf near Dippoldiswalde, SaK 478, ×1. b, c Original illustrations of the holotype by Geinitz (1849: pl. 12, figs. 6, 7) as Spongia ottoi. d Illustration of the holotype in Geinitz (1871: pl. 8, fig. 8) as Epitheles furcata Goldfuss

Details of the holotype of Dactyloidites ottoi (Geinitz, 1849) from the Oberhäslich Formation (Saxony), Wendischcarsdorf near Dippoldiswalde (SaK 478). w = width of photomicrograph. a Close-up of the specimen in Fig. 3b showing enrichment of mica flakes between branching radial elements (arrows); w = 35 mm. b Close-up of the specimen in Fig. 3c showing sharply separated radial elements of constant width; w = 35 mm. c Close-up of the distal part of the specimen above Fig. 3b showing branching and enrichment of mica flakes between radial elements (arrows); w = 25 mm. d Cross-section view showing vertical spreiten-like structure (arrows) related to selective weathering of the burrow wall; w = 8 mm. E1 Cross-section view showing potential vertical protrusive spreiten structure, with interpretation in E2; w = 9 mm

Sandstone slab with numerous Dactyloidites ottoi (Geinitz, 1849) from the Oberhäslich Formation (Saxony), Wendischcarsdorf near Dippoldiswalde, SaK 15489. a Overview showing the dense packing of D. ottoi preserved as negative epireliefs. Details of individual D. ottoi showing the palmate (b) to nearly circular (e) shapes of the rosettes as well as the trunks of the vertical shafts (c, d). Scale in mm

Dactyloidites ottoi (Geinitz, 1849) from the Regensburg Formation (Bavaria), stairs of the Befreiungshalle near Kelheim, bedding-plane views. w = width of photograph. a Set of three rosettes; w = 8 cm. b Two rosettes cut in different levels; w = 10 cm. c Densely crowded arrangement of rosettes; w = 16 cm. d Overview of bedding plane with several rosettes, in part cross-cut by Ophiomorpha isp. (arrow); w = 30 cm. e Detail of rosette in left centre of d showing the different sediment fabric in radial elements (fine-grained quartz sandstone, light) and glauconitic host rock (dark); w = 5 cm
* | 1849 | Spongia ottoi Geinitz: 264, pl. 12, figs. 6, 7. |
1854 | Spongia ottoi Geinitz. Otto: 12, pl. 4, fig. 7. | |
* | 1871 | Epitheles furcata Goldf. sp. Geinitz: 34, pl. 8, fig. 8. |
1930 | Spongia ottoi Geinitz, 1849. Häntzschel: 261, figs. 1–3. | |
1937 | Gyrophyllites cf. kwassizensis Glocker 1841. Lehner: 190, pl. 17, fig. 37. | |
1960 | Spongia shikokuensis Katto, new species. Katto: 324, pl. 34, figs. 10, 13. | |
1975 | Haentzschelinia ottoi (Geinitz, 1849). Häntzschel: W 65, fig. 42, 3a, b. | |
1985 | Dactyloidites ottoi (Geinitz 1849). Fürsich and Bromley: 199, figs. 1–9, 10 (right figure). | |
1993 | Dactyloidites ottoi (Geinitz, 1849). Pickerill et al.: 1073, figs. 2, 3. | |
1995 | Dactyloidites ottoi (Geinitz, 1849). Gibert et al.: 770, text-fig. 2, pl. 91, figs. 2–4 [see for additional synonymy]. | |
1997 | Häntzschelinia ottoi. Seilacher: 48, fig. 28. | |
1999 | Dactyloidites ottoi (Geinitz, 1849). López-Martínez and Ardèvol: 131, fig. 3. | |
Non | 2002 | ?Dactyloidites cf. ottoi (Geinitz 1849). Kappel: 85, pl. 3, figs. 2, 3 [Lorenzinia isp.?]. |
2004 | Dactyloidites ottoi (Geinitz, 1849). Agirrezabala and Gibert: 276, figs. 5a–c, 6a–f. | |
2004 | Dactyloidites ottoi (Geinitz, 1849). Blisset and Pickerill: 348, pl. 3, fig. A. | |
2007 | Dactyloidites ottoi (Geinitz). Gibert et al.: 3, fig. 5b, c. | |
2008 | Dactyloidites ottoi (Geinitz, 1849). Lazo et al.: 628, figs. 2, 3. | |
2010 | Dactyloidites ottoi (Geinitz, 1849). Srivastava et al.: 172, pl. 1, figs. 1–3. | |
2011 | Dactyloidites ottoi (Geinitz). Göhler: 59, figs. 2, 3. |
Types:
Holotype is a single sandstone slab (SaK 478) from the Upper Cenomanian Oberhäslich Formation of Wendischcarsdorf, Saxony. This slab displays four rosettes of D. ottoi, two of which were originally illustrated by Geinitz in separate figures (1849: pl. 12, figs. 6, 7; see Fig. 3).
Emended diagnosis:
Fan-shaped, rarely palmate spreiten structure originating from a central, vertical to oblique shaft leading downwards into the sediment. The radial elements are subhorizontal protrusive vertical spreiten (probes), mostly forming incomplete circular rosettes of 200°–270°. Number of radial elements up to 20 due to branching (bi- and trifurcation) of 6–9 primary probes. Diameter of rosettes between 30 and 75 mm, width of radial elements between 4 and 6 mm.
Material:
The Saxonian type material of D. ottoi (Geinitz, 1849) is from the Oberhäslich Formation at Wendischcarsdorf near Dippoldiswalde and comprises two specimens in the lower left corner of a ca. 160 × 150 mm2 sandstone slab, accompanied by two additional, poorly preserved specimens (SaK 478, see Figs. 3a, 4). Furthermore, about 20 D. ottoi are preserved on a large sandstone slab (SaK 15489, height 595 mm; see Fig. 5) from the same locality and bed, first illustrated by Häntzschel (1930: fig. 2); ca. 10 additional specimens are visible on another large slab (SaK 481, height ca. 500 mm). It has to be noted that Häntzschel (1930) reported three large sandstone slabs with D. ottoi from Wendischcarsdorf which had been in the collection of the MMG at that time, the largest of which he illustrated in his fig. 2. This latter one is only fragmentarily preserved (Fig. 5a), together with another fragment (SaK 481, not illustrated). The third large sandstone slab is completely missing. Based on the relatively new appearance of the fracture surfaces in SaK 15489 and SaK 481, it can be concluded that this loss is related to the destruction of the Dresden museum towards the end of World War 2.
The Bavarian material consists of several tens of specimens from the (upper) freestone bed of the Saal Member of the Regensburg Formation at Ihrlerstein, north of Kelheim. D. ottoi is preserved as faintly positive reliefs on the stairs of the Befreiungshalle in Kelheim (Fig. 6), which have been in place since the grand opening 150 years ago (B. Schels in Haala and Manstorfer 2012). Further records are from the flood masonry walls and the bridge arches of the Steinerne Brücke of Regensburg (Meyer and Schmidt-Kaler 1995: fig. B 6 right). Rutte (1982, 6) also mentioned the abundant “rosetten- bis sternchenförmigen Figuren von maximal 5 Zentimetern Durchmesser” from the stairs of the Befreiungshalle that, according to him, had not yet been recorded in the literature.
Description:
Dactyloidites ottoi is preserved as positive hyporelief on a bedding surface of a sandstone bed in case of the holotype (SaK 478). It consists of 7–9 primary radial elements 4–5 mm in diameter originating directly from a central depression (representing the position of the central shaft). The radial elements curve into the central depression, suggesting that it represents a lower bed surface. Most primary branches bifurcate, producing the 15–20 radial elements present in the outermost parts of the structure. The diameter of the radial elements is nearly constant (Figs. 3a, 4a–c). Also the diameter of the four rosettes is nearly constant, ranging between 55 and 60 mm. Full circles of radial elements are not developed (although the specimen of Fig. 3c comes close), often ranging between 180° and 270°. The host rock is a light-grey, fine-grained quartz sandstone with mica flakes while the radial elements consist of mica-free, fine-grained quartz sandstone (Fig. 4a–c). Mica flakes are enriched between the radial elements (Fig. 4c). Spreiten structures are difficult to recognize due to the high maturity and homogeneous fabric of the host rock. However, the observation that the radial elements weather as ridges (rather than simple tubes) separated by less resistant layers of wall material (enrichment of mica flakes in the grooves) suggests that they are in fact vertical spreite structures (cf. Fürsich and Bromley 1985). Support for this comes from weakly developed vertical spreiten-like structures in cross-section (Fig. 4d, e, the latter showing protrusive spreiten as proposed by Fürsich and Bromley 1985).
On the large sandstone slab SaK 15489, D. ottoi occurs with around 20 specimens in negative epireliefs (Fig. 5a). Rosettes are subhorizontally oriented, close to fully circular (Fig. 5e) or fan-shaped arranged (Fig. 5b), with diameters varying between 59 and 75 mm. The radial elements are 4–6 mm wide, originating from a central area where in some cases relicts of the vertical shaft are preserved (e.g. Fig. 5c, d). In the periphery, up to 20 bluntly ending radial elements can be counted (Fig. 5e), resulting from common branching. The host rock is a fine- to medium-grained, feldspar-bearing sandstone of brownish colour. The fill of the radial elements is not preserved, and spreiten structures have not been observed. The upwards orientation of the radial elements towards the central, elevated shaft suggests that SaK 15489 represents an upper bedding surface.
The specimens from the Regensburg Formation consist of up to 16 radial elements that fan out up to 270° (Fig. 6a–e). Branching of the radial elements is common. In comparison with the surrounding sediment (a calcareous, fine- to medium-grained, glauconitic quartz sandstone), the radial elements weather as weak ridges that lack glauconite and consist of light-coloured, well-cemented, fine-grained quartz sandstone (Fig. 6). The lithology corresponds to the upper freestone bed of the Ihrlerstein Quarry (Fig. 2). The radius of the structures varies between 30 and 50 mm, while the diameter of the radial elements is rather constant, ranging between 4 and 5 mm. Due to the only two-dimensional, bedding-parallel exposure of D. ottoi at the Befreiungshalle, it cannot be ascertained whether or not vertical spreiten are present. The rosettes of D. ottoi are cross-cut by later Ophiomorpha isp. and Laminites isp. (Fig. 6d), and bioturbation can be pervasive, reaching a ichnofabric index of five (Fig. 7a–c; cf. Droser and Bottjer 1986). Coarse-grained, bioclastic beds with hummocky cross-stratification and scoured bases are intercalated (Fig. 7c, d).

Other trace fossils and sedimentary structures from the Regensburg Formation (Bavaria), stairs of the Befreiungshalle near Kelheim. w = width of photograph. a Laminites isp. (L) and Ophiomorpha isp. (O), bedding-plane view, w = 10 cm. b Actively backfilled burrows of Ophiomorpha isp., bedding-plane view, w = 16 cm. c Strongly bioturbated, fine- to medium-grained, glauconitic sandstone with Ophiomorpha isp. (O) and Laminites isp. (L), cut by erosional scour at the base of a tempestite (see also d); bedding-plane view, w = 95 cm. d Erosional, hummocky cross-stratified, coarse-grained bioclastic tempestite cutting into fine- to medium-grained, glauconitic sandstone rich in traces of Ophiomorpha isp. (arrows), section perpendicular to bedding, w = 35 cm
Occurrence:
The present material comes from the Lower–Middle Cenomanian of the Regensburg Formation (Bavaria) and the Upper Cenomanian of the Oberhäslich Formation (Saxony). Furthermore, D. ottoi has been described from the Middle Jurassic of India (Srivastava et al. 2010), the Upper Jurassic of Argentina (Lazo et al. 2008), the Lower (Agirrezabala and de Gibert 2004) and Upper Cretaceous of Spain (López-Martínez and Ardèvol 1999), the Upper Cretaceous of Greenland (Fürsich and Bromley 1985), the Coniacian of Bavaria (Lehner 1937), the Eocene and Miocene of Spain (de Gibert et al. 1995, 2007), the Oligocene and Miocene of Japan (Katto 1960; Seilacher 1997) and the Oligocene–Neogene of Jamaica (Pickerill et al. 1993; Blissett and Pickerill 2004). If Haentzschelinia Vyalov, 1964 is placed in the synonymy of Dactyloidites (see below), the range of D. ottoi extends back to the Triassic.
Discussion
Ichnotaxonomic issues
A trace fossil similar to D. ottoi has been described by Glocker (1841, 320–324) from fine-grained, marine glauconitic sandstones of Cenomanian age at the Capellenberg near Kwassitz (Kvasice), Moravia, Czech Republic. These sandstones are rich in trace fossils, regarded as plant fossils at that time, and Glocker figured, described and named a rosetted form, Gyrophyllites kwassizensis. The trace consists of a shaft with horizontally radiating, leaf-like elements, resulting in a flower-like form (his “zehenblättriger Verticill”). The rosette has a radius of 30–40 mm, and the radial elements are 3–5 mm in diameter, being somewhat club-shaped. Eight years later, Geinitz (1849) erected S. ottoi based on the occurrence in Cenomanian sandstones of Saxony, described above. In 1854, Otto described inferred floral remains from the Cretaceous of Saxony and also discussed S. ottoi, which he regarded as remains of amorphozoans (= sponges), following Geinitz. He also stated that he and H.B. Geinitz regarded G. kwassizensis and S. ottoi as identical and that Glocker’s species has priority (Otto 1854, 13). In 1871, Geinitz re-described his S. ottoi as Epitheles furcata (Goldfuss, 1826), a siliceous sponge known from the Cenomanian Essen Greensand Formation of the Münsterland Cretaceous Basin, and repeated his statement that he regarded it as synonymous to G. kwassizensis Glocker, 1841. It should be noted in this respect that D. ottoi has also been reported as Gyrophyllites cf. kwassizensis by Lehner (1937) from the Lower to Middle Coniacian Hartmannshofer Sandstein (Jeding Formation of Niebuhr et al. 2009) of the Hubmersberg, east of Nürnberg (Bavaria).
Gyrophyllites Glocker is now regarded as a feeding trace with an oblique to vertical shaft from which 5–20 (average 10) club- or leaf-shaped feeding tunnels radiate at different superimposed levels in rosetted or helical arrangement, essentially without spreiten structure (cf. Häntzschel 1975; Fürsich and Kennedy 1975). However, in the original description by Glocker (1841), neither a helical arrangement with several levels of rosettes nor spreiten structures are mentioned. Unfortunately, Glocker’s type is lost (Otto 1854), and it cannot be proved that G. kwassizensis and S. ottoi are in fact identical (S. ottoi would then be a subjective junior synonym of G. kwassizensis). However, considering the lost type and the fact that the radial elements in the illustration by Glocker (1841) are club-shaped and unbranched (in contrast to S. ottoi), Geinitz’s species is retained as valid. The absence of vertical spreiten structures and branching as well as the club-shaped form of the radial elements with several levels of rosettes may serve as the diagnostic key features to separate Gyrophyllites from Dactyloidites.
Another ichnotaxonomic issue is the “Dactyloidites–Haentzschelinia problem”. Vyalov (1964) erected the ichnogenus Häntzschelinia [sic] (type ichnospecies S. ottoi Geinitz) and described two new species from Triassic sandstones of Sibiria, Häntzschelinia kolymensis and H. pygmaea. Judging from the photographs (Vyalov 1964, figs. 2, 3), they are very similar to D. ottoi, especially H. pygmaea. Fürsich and Bromley (1985) argued that the basic behaviour pattern in Haentzschelinia Vyalov, 1964 (rosetted vertical spreiten with central shaft) is the same as that of Dactyloidites Hall, 1886 and Brooksella Walcott, 1896. Therefore, Dactyloidites Hall should have priority, and both H. kolymensis Vyalov and H. pygmaea Vyalov were tentatively placed in the synonymy of D. ottoi (Geinitz) (Fürsich and Bromley 1985, 207). Most authors followed their view (e.g. Pickerill et al. 1993; de Gibert et al. 1995, 2007, among others), although others expressed scepticism (e.g. Schweigert 1998) or stressed the need for a general revision of rosetted trace fossils (Uchman and Pervesler 2007). However, most of the criticism is unfounded from a purely ichnotaxonomic viewpoint (see Bertling et al. 2006 for details).
Vyalov (1989) argued against Fürsich and Bromley (1985) mainly because of the fact that the radial elements in Haentzschelinia are numerous and of constant width compared with Dactyloidites Hall, which is also much larger. However, in ichnotaxonomy, fossil behaviour is classified, being basically the same in both ichnogenera, and the number of radial elements (which is mainly dependent on branching) and the total size may be used for ichnospecies separation (note that the width of the radial elements is also constant in other Dactyloidites species; cf. Fürsich and Bromley 1985; Uchman and Pervesler 2007). The high age (Cambrian) and the differing depositional setting (deep marine) of the type material of Dactyloidites Hall are also not a valid ichnotaxonomic argument. The same holds true for the observation that the radial elements in Haentzschelinia Vyalov rarely describe fully circular rosettes but are mainly fan-shaped (Schweigert 1998). This has been explained by Fürsich and Bromley (1985) by maximum resource utilization and is also related to an inclination of the central shaft.
Seilacher (2007) recognized both ichnogenera and placed them in his probers group (burrow systems of actively backfilled, blind-ending branches), also commonly called “fucoids”. The Cambrian Dactyloidites is displayed as a star-shaped trace fossil (“medusiform gyrophyllitid”) with a central shaft coming from above, and the radial, unbranched tunnels are characterized by radial protrusive spreiten. In Haentzschelinia, one of Seilacher’s (2007) “various fucoids”, the branching tunnels are spread in palmate pattern from an oblique shaft and are backfilled with sand. No further ichnotaxonomic information is available from this work.
In conclusion, most (if not all) of the counter-arguments against a synonymization of Haentzschelinia with Dactyloidites are unfounded from an ichnotaxonomic viewpoint. Size, age and depositional environment as well as shape of rosettes and number of radial elements are not valid arguments for ichnogeneric separation of the traces under discussion (cf. Bertling et al. 2006; Buatois and Mángano 2011). Thus, in the absence of an urgently necessary revision of rosetted trace fossils, the inclusion of S. ottoi in Dactyloidites is the most reasonable classification for the time being.
The potential tracemaker
The potential tracemaker of D. ottoi has been discussed by several authors. The first one to have ever mentioned the possibility of an animal trace was in fact Otto (1852, 26), but he discarded that idea because his friend, the influential palaeontologist H.B. Geinitz, regarded it as a sponge (Geinitz 1849). Surprisingly, some time later Geinitz (1871) placed his S. ottoi in the synonymy of E. furcata, a lithistid sponge erected by Goldfuss (1826). After that, the species fell into obliteration again, but Morin (1907) compared S. ottoi with fiddler crab feeding traces. Häntzschel (1930) finally documented that S. ottoi is in fact a “Lebensspur”, presumably formed by decapod or amphipod crustaceans, although also bivalves can produce star-shaped surface trails (Häntzschel 1934). A short-lived “step back” was performed by Katto (1960, 324, pl. 34, figs. 10, 13), who erected S. shikokuensis based on numerous specimens in clean, medium-grained nearshore sandstones of the Misaki Formation of Shikoku (Oligocene, Japan), clearly representing D. ottoi. Fürsich and Bromley (1985) revived the discussion of the enigmatic trace and suggested an infaunal formation by a worm-like organism, possibly possessing a proboscis for sediment reworking. Seilacher (1997, 48–49) discussed several possibilities (undermat mining, sediment feeding, sanitation, fluid pumping) for the formation of D. ottoi (termed Häntzschelinia [sic] ottoi by him). He stated that the radial probes most likely became actively backstuffed by their maker, suggesting a feeding trace, but also did not discount sanitation. However, he obviously did not recognize any spreiten structures.
The view of Dactyloidites as an infaunal feeding structure potentially produced by a worm-like animal (cf. Fürsich and Bromley 1985) is supported by studies on the feeding behaviour of the modern lugworm Arenicola marina by Rijken (1979; see Fig. 8). A. marina usually sits at the bottom of a J- or U-shaped tube in the sediment. Through the open end of the tube, oxygen-rich water enters and flows downward by peristaltic movements of the worm. At the closed end of the tube, the water is pressed through the sediment upwards, creating a column of a semi-fluid mixture of sand and water. The lugworm swallows the sediment at the lower part of this “quick-sand column” which gradually sinks downward so that a funnel develops at the sediment–water interface, acting as a trap for organic material. However, under specific circumstances, the lugworm switches from this “normal” feeding strategy to an alternative one: when nutrient-rich sediments occur at living depth, it directly feeds on the surrounding sediment by generating radial tunnels originating from the central shaft. By this feeding strategy, composite rosetted structures very similar to D. ottoi may form (Fig. 8), and vertical shift of the burrows may enhance sediment exploitation and result in spreiten structures (cf. Fürsich and Bromley 1985).

Generation of Dactyloidites ottoi-like structures by the Recent lugworm Arenicola marina. a Normal feeding behaviour within U-shaped burrow containing quick-sand column and funnel. b Facultative feeding behaviour when organic-rich sediments occur at living depth. Modified after Rijken (1979). See text for further explanations
Almost all occurrences of D. ottoi are from shallow-water, nearshore to deltaic, potentially nutrient-rich environments that are compatible with the occurrences of worms with very similar ecological requirements and behaviour as the Recent lugworm A. marina. The absolute majority of the known occurrences are from siliciclastic facies (see compilation in Agirrezabala and de Gibert 2004), but a few occurrences are also from carbonates (Blissett and Pickerill 2004; Lazo et al. 2008). D. ottoi was recently also recorded from medium-grained, calcareous sandstone beds of the Khadir Formation (Upper Bathonian–Lower Callovian) of Kachchh, India (Srivastava et al. 2010). However, deltas and siliciclastic coasts with ample terrestrial nutrient input and water depths of a few metres only seem to support the formation of D. ottoi (see also summary in Agirrezabala and de Gibert 2004), the producer of which most probably required particulate organic matter directly ingested from nutrient-rich sediments some distance below the sediment–water interface. Evidence for reduced salinities (Agirrezabala and de Gibert 2004) is not supported by the studied occurrences from Saxony and Bavaria as the co-occurring faunas are fully marine.
Dactyloidites ottoi is a relatively rare trace fossil. Both the type material and the large sandstone slabs from Saxony are all from the uppermost bed (of the Oberhäslich Formation) of a single locality (the “Wolf’scher Quaderbruch” in Wendischcarsdorf), where they obviously occurred in great abundance (Otto 1852, 26). Another record from the same formation, ca. 18 km westnorthwest of Dippoldiswalde, was recently reported by Göhler (2011). Towards the top of the Oberhäslich Formation, shallowing is recorded, shifting the environment into the nearshore zone with rare records of Skolithos isp. (Wilmsen and Richardt 2012). Thus, in this example D. ottoi is preserved at the top of a shallowing-upward cycle characterized by high sedimentation rates (late highstand of depositional sequence Cenomanian 5; Richardt and Wilmsen 2013), and the transition from the Cruziana into the lower Skolithos ichnofacies (cf. Seilacher 1967). High sedimentation rates have also been proposed for the records of D. ottoi in the Aptian–Albian Otoio Formation of the Basque–Cantabrian Basin, Spain (Agirrezabala and de Gibert 2004). However, D. ottoi from the Regensburg Formation of the Ihrlerstein Quarry stems from a fining-upward cycle (i.e. transgressive systems tract of depositional sequence Cenomanian 4; see Wilmsen et al. 2010 for sequence stratigraphic details). For the Ihrlerstein occurrence, it can be shown that D. ottoi is cross-cut by Ophiomorpha isp. and Laminites isp., which may occur in super-abundance (up to ichnofabric index 5 of Droser and Bottjer 1986; see Fig. 6d). Thus, originally more abundant D. ottoi may have been destroyed by subsequent bioturbation (Laminites isp. is a trace produced by burrowing echinoids; Plaziat and Mahmoudi 1988). This is related to low accumulation rates during deepening of the depositional environment and bioturbation by later-tier traces of the Cruziana ichnofacies. Support for this interpretation comes from intercalated hummocky cross-stratified (HCS) tempestites within the Ihrlersteiner Grünsandstein: their rapid deposition may have occasionally “frozen” an ichnofabric in different developmental stages, preventing complete homogenization of the sediment, which is characteristic of other occurrences of the Saal Member of the Regensburg Formation (where tempestites are unknown). The presence of HCS also demonstrates that D. ottoi from the Regensburg Formation formed below the fair weather and above the storm wave base (cf. Myrow and Southard 1996), thus belonging to the (upper) Cruziana ichnofacies of Seilacher (1967).
Another factor explaining the relative rarity of D. ottoi may be related to the variable feeding behaviour of the worm-like tracemaker, which only in special cases (nutrient-rich sediment at living depth) will result in the formation of rosetted trace fossils. The distribution of such sediments in nearshore settings may be very local. Thus, both formation and preservation of D. ottoi are related to a narrow palaeoenvironmental window and specific taphonomic conditions, very well explaining the scarcity of the trace in the stratigraphic record.
Conclusions
The rosetted trace fossil D. ottoi (Geinitz, 1849) is discussed based on new occurrences from Cenomanian glauconitic sandstones of the Regensburg Formation (Bavaria, southern Germany) as well as a careful re-description of the type material and other specimens from the Cenomanian Oberhäslich Formation (shallow-marine quartz-rich sandstones) of Saxony (eastern Germany), housed in the Senckenberg Naturhistorische Sammlungen Dresden.
Dactyloidites ottoi is a feeding trace consisting of a fan-shaped to palmate spreiten structure originating from a central, vertical to oblique shaft leading downwards into the sediment. The numerous branching radial elements (up to 20, 4–6-mm-thick tubes of equal width) are subhorizontal protrusive vertical spreiten, mostly forming incompletely circular rosettes with radii of 200°–270° and diameters varying between 30 and 75 mm.
The ichnotaxonomic classification of the trace has been discussed controversially. G. kwassizensis Glocker, 1841 is a trace fossil similar to D. ottoi. It has been described from contemporaneous glauconitic nearshore sandstones in the Bohemian Cretaceous Basin and consists of a shaft with horizontally radiating, leaf-like elements, resulting in a flower-like form. Although Otto (1854) and Geinitz (1871) stress the concordance of both structures, Geinitz’s species is considered valid based on the fact that the holotype of G. kwassizensis (which is the type ichnospecies of Gyrophyllites) is lost and that the radial elements in G. kwassizensis are club-shaped and unbranched and do not show any spreiten.
The validity and necessity of the ichnogenus Haentzschelinia Vyalov, 1964 are questioned because the essential behaviour expressed by its form (rosetted vertical spreiten with central shaft) is basically the same as in Dactyloidites Hall, 1886, which then has priority (Fürsich and Bromley 1985). The counter-arguments against a synonymization of Haentzschelinia with Dactyloidites are unfounded from a strict ichnotaxonomic viewpoint because size, age, depositional environment, shape of rosettes (circular vs. fan-shaped) and number of radial elements are not valid arguments for an ichnogeneric separation of the traces. Thus, the classification of S. ottoi in Dactyloidites is reasonable. However, rosetted trace fossils are in urgent need of a careful general revision.
The potential tracemaker of D. ottoi has been widely discussed in the literature. The view of Fürsich and Bromley (1985) of a formation by a worm-like organism systematically reworking the sediment for organic food particles is supported by studies on the feeding behaviour of the Recent lugworm A. marina. Under specific circumstances (nutrient-rich sediments at living depth), the lugworm switches from its normal feeding strategy (U-shaped burrow with funnel trapping surficial food particles) to direct ingestion of the surrounding sediment by generating radial tunnels originating from the central shaft. By this feeding strategy, composite rosetted structures very similar to D. ottoi may form, and vertical shifts result in spreiten structures.
Dactyloidites ottoi has a proven range from the Jurassic to the Neogene and predominantly occurs in shallow-water, nearshore to deltaic, potentially nutrient-rich siliciclastic settings. Deltas and siliciclastic coasts with ample terrestrial nutrient input seem to support the formation of D. ottoi in the lower Skolithos and upper Cruziana ichnofacies, and its worm-like producer most probably fed on particulate organic matter ingested directly from nutrient-rich sediments some distance below the sediment–water interface. The formation of the trace is thus dependent on relatively shallow-marine conditions and the presence of organic-rich sediments at living depth of the producer. This narrow palaeoenvironmental window is furthermore closed by preservational constraints: high sedimentation rates towards the top of shallowing-upward cycles may preserve local nearshore occurrences of D. ottoi, while low accumulation rates during transgressive conditions enhance the chances of their destruction by subsequent bioturbation. We thus conclude that both formation and preservation of D. ottoi are related to a narrow palaeoenvironmental window and specific taphonomic conditions, readily explaining the rarity of the ichnotaxon in the stratigraphic record.
References
Agirrezabala, L.M., and J.M. de Gibert. 2004. Paleodepth and paleoenvironment of Dactyloidites ottoi (Geinitz, 1849) from Lower Cretaceous deltaic deposits (Basque-Cantabrian Basin, west Pyrenees). Palaios 19: 276–291.
Bertling, M., S.J. Braddy, R.G. Bromley, G.D. Demathieu, J. Genise, R. Mikuláš, J.K. Nielsen, K.S.S. Nielsen, A.K. Rindsberg, M. Schlirf, and A. Uchman. 2006. Names for trace fossils: A uniform approach. Lethaia 39: 265–286.
Blissett, D.J., and R.K. Pickerill. 2004. Soft-sediment ichnotaxa from the Cenozoic White Limestone Group, Jamaica, West Indies. Scripta Geologica 127: 341–378.
Buatois, L.A., and M.G. Mángano. 2011. Ichnology—organism–substrate interactions in space and time. i–xii. Cambridge: University Press,.
de Gibert, J.M., J. Martinell, and R. Domènech. 1995. The rosetted feeding trace fossil Dactyloidites ottoi (Geinitz) from the Miocene of Catalonia. Geobios 28: 769–776.
de Gibert, J.M., M. López-Blanco, and E. Ramos. 2007. Presencia de la icnoespecie Dactyloidites ottoi en el complejo de abanico costero de Sant Llorenç del Munt (Eoceno, Cuenca del Ebro, NE de España). Geogaceta 41: 3–6.
Droser, M., and D.J. Bottjer. 1986. A semiquantitative field classification of ichnofabric. Journal of Sedimentary Petrology 56: 558–559.
Endlicher, G. 1984. Petrographisch-mineralogische Untersuchungen der Bausteine und Verwitterungskrusten des Regensburger Domes. Acta Albertina Ratisbonensis 42: 53–80.
Fitch, A. 1850. A historical, topographical and agricultural survey of the County of Washington. Part 5. New York Agricultural Society, Transactions 9(1849): 753–944.
Fürsich, F.T., and W.J. Kennedy. 1975. Kirklandia texana Caster – Cretaceous hydrozoan medusoid or trace fossil chimaera? Palaeontology 18: 665–679.
Fürsich, F.T., and R.G. Bromley. 1985. Behavioural interpretation of a rosetted spreite trace fossil Dactyloidites ottoi (Geinitz). Lethaia 18: 199–207.
Geinitz, H.B. 1849. Das Quadersandsteingebirge oder Kreidegebirge in Deutschland. 1–292. Freiberg: Craz and Gerlach.
Geinitz, H.B. 1871–1875. Das Elbthalgebirge in Sachsen. Teil I – Der untere Quader. Palaeontographica 20(1): 1–276.
Glocker, E.-F. 1841. Über die kalkführende Sandsteinformation auf beiden Seiten der mittleren March, in der Gegend zwischen Kwassitz und Kremsier. Nova Acta Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum 19 (Suppl. 2): 309–334, pls 1–4.
Göhler, T. 2011. Betrachtungen einiger Grabgangstücke [u.a. Ophiomorpha nodosa Lundgren und Thalassinoides saxonicus (Geinitz)] aus obercenomanen Sandsteinen des Tharandter Waldes. BGSK (Beiträge zur Geologie der Sächsischen Kreide)-Info 2011: 59–64.
Goldfuss, G.A. 1826–1844. Petrefacta Germaniae. 198 pls. Düsseldorf: Arnz & Comp.
Haala, H., and S. Manstorfer. 2012. Einzigartige Funde auf der Befreiungshalle. Mittelbayerische Zeitung 05.12.2012: 25.
Häntzschel, W. 1930. Spongia ottoi Geinitz, ein sternförmiges Problematikum aus dem sächsischen Cenoman. Senckenbergiana 12: 261–275.
Häntzschel, W. 1934. Sternspuren erzeugt von einer Muschel: Scrobicularia plana (da Costa). Senckenbergiana 16: 325–330.
Häntzschel, W. 1975. Trace fossils and Problematica. Treatise on Invertebrate Palaeontology, Part W Miscellanea, Supplement 1: i–xxi, W1–W269.
Hafner, E. 1998. Geschichte von Ihrlerstein. Kelheim: Leik.
Hall, J. 1886. Note on some obscure organisms in the roofing slate of Washington County, New York. Trustees New York State Museum of Natural History, Annual Report 39: 160, pl. 11.
Kappel, J. 2002. Ichnofossilien im Campanium des südöstlichen Münsterlandes. Dissertation, Westfälische Wilhelms-Universität Münster, 1–236, 13 pls. [unpubl.].
Katto, J. 1960. Some problematica from the so-called unknown Mesozoic strata of the southern part of Shikoku, Japan. Science Reports Tohoku University, 2nd Series (Geology), Special Volume 4: 323–334.
Lazo, D.G., R.M. Palma, and R.D. Piethé. 2008. La traza Dactyloidites ottoi (Geinitz) en la Formación La Manga, Oxfordiano de Mendoza. Ameghiniana 45: 627–632.
Lehner, L. 1937. Fauna und Flora der Fränkischen Albüberdeckenden Kreide. II. Fauna 2. Teil und Flora – Herausgegeben von R. Dehm. Palaeontographica 87. Abteilung A 3–6: 158–230.
López-Martínez, N., and L. Ardèvol. 1999. Hallazgo de Dactyloidites ottoi (Geinitz, 1849) en la Arenisca de Arén (Cretácico Superior Surpirenaico). Revista Española de Paleontología, numero extraordinario: 129–133.
Meyer, R.K.F., and H. Schmidt-Kaler. 1995. Rund um Regensburg. Wanderungen in der Erdgeschichte 7: 1–128.
Morin, H. 1907. Ein Rätsel weniger. Verhandlungen der Kaiserlich-Königlichen zoologisch-botanischen Gesellschaft in Wien: 267–270.
Myrow, P.M., and J.B. Southard. 1996. Tempestite deposition. Journal of Sedimentary Research 66: 875–887.
Niebuhr, B., M. Hiss, U. Kaplan, K.-A. Tröger, S. Voigt, T. Voigt, F. Wiese, and M. Wilmsen. 2007. Lithostratigraphie der norddeutschen Oberkreide. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 55: 1–136.
Niebuhr, B., T. Pürner, and M. Wilmsen. 2009. Lithostratigraphie der außeralpinen Kreide Bayerns. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 65: 7–58.
Philip, J., and M. Floquet. 2000. Late Cenomanian (94.7–93.5). In Atlas Peri-Tethys palaeogeographical maps, ed. J. Dercourt, M. Gaetani, B. Vrielynck, E. Barrier, B. Biju-Duval, M.F. Brunet, J.P. Cadet, S. Crasquin, and M. Sandulescu, 129–136. Paris: CCGM/CGMW.
Pickerill, R.K., S.K. Donovan, and H.L. Dixon. 1993. The trace fossil Dactyloidites ottoi (Geinitz, 1849) from the Neogene August Town Formation of South-Central Jamaica. Journal of Paleontology 67: 1070–1074.
Plaziat, J.-C., and M. Mahmoudi. 1988. Trace fossils attributed to burrowing echinoids: A revision including new ichnogenus and ichnospecies. Geobios 21: 209–233.
Richardt, N., and M. Wilmsen. 2013. Sequence stratigraphy of the lower Upper Cretaceous Elbtal Group (Cenomanian–Turonian of Saxony, Germany). Zeitschrift der Deutschen Gesellschaft für Geowissenschaften (submitted).
Richardt, N., M. Wilmsen, and B. Niebuhr. 2013. Late Cenomanian–Early Turonian facies development and sea-level changes in the Bodenwöhrer Senke (Danubian Cretaceous Group, Bavaria, Germany). Facies 59. doi:10.1007/s10347-012-0337-x.
Rieger, G., and H. Rieger.1954. Kelheimer Heimatbuch für die Stadt und den Landkreis Kelheim, 2nd ed. Kelheim: Leik.
Rijken, M. 1979. Food and food uptake in Arenicola marina. Netherlands Journal of Sea Research 13: 406–421.
Rutte, E. 1982. Natursteine der Befreiungshalle. Weltenburger Akademie, Schriftenreihe 4 (4): 1–7.
Schweigert, G. 1998. Die Spurenfauna des Nusplinger Plattenkalks (Oberjura, Schwäbische Alb). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie) 262: 1–47.
Seilacher, A. 1967. Bathymetry of trace fossils. Marine Geology 5: 413–428.
Seilacher, A. 1997. Fossil art. An exhibition of the Geologisches Institut Tübingen University. Drumheller, Alberta: The Royal Tyrrell Museum of Palaeontology.
Seilacher, A. 2007. Trace fossil analysis. New York: Springer.
Srivastava, D.K., M.P. Singh, and A.K. Kulshrestha. 2010. The trace fossil Dactyloidites Hall, 1886 from the Middle Jurassic Khadir Formation of Bela Island, Kachchh, India and its palaeoenvironmental significance. Journal of the Palaeontological Society of India 55: 171–176.
Tröger, K.-A. 2003. The Cretaceous of the Elbe valley in Saxony—a review. Carnets de Géologie A03: 1–14.
Uchman, A., and P. Pervesler. 2007. Palaeobiological and palaeoenvironmental significance of the Pliocene trace fossil Dactyloidites peniculus. Acta Palaeontologica Polonica 52: 799–808.
von Otto, E. 1852. Addidamente zur Flora des Quadergebirges in der Gegend um Dresden und Dippoldiswalde, enthaltend meist noch nicht oder wenig bekannte fossile Pflanzen. 1–30, 7 pls. Dippoldiswalde: C. Sehne.
von Otto, E. 1854. Addidamente zur Flora des Quadergebirges in Sachsen. 2. Heft. 1–54, 9 pls.; Leipzig: G. Mayer.
Vyalov, O.S. 1964. Zvezdchatye ieroglify iz Triasa severovostoka Sibiri (Star-shaped hieroglyphs from the Triassic of northeastern Sibiria). Geologija i Geofizika 5: 112–115. (in Russian, with English summary).
Vyalov, O.S. 1989. Paleoichnological studies. Paleontologičeskij Sbornik 26: 72–78. (in Russian, with English summary).
Walcott, C.D. 1896. Fossil jelly fishes from the Middle Cambrian terrane. United States National Museum, Proceedings 18: 611–614, pls 31, 32.
Wilmsen, M., and E. Nagm. 2013. Upper Cenomanian–Lower Turonian ammonoids from the Saxonian Cretaceous (lower Elbtal Group, Saxony, Germany). Bulletin of Geosciences (in press).
Wilmsen, M., and B. Niebuhr. 2010. On the age of the Upper Cretaceous transgression between Regensburg and Neuburg an der Donau (Bavaria, southern Germany). Neues Jahrbuch für Geologie und Paläontologie 256: 267–278.
Wilmsen, M., and N. Richardt. 2012. Biofacies, depositional environment and stratigraphy of the Oberhäslich Formation (lower Upper Cenomanian, Saxonian Cretaceous Basin, Germany). Terra Nostra 2012 (3): 199–200.
Wilmsen, M., B. Niebuhr, P. Chellouche, T. Pürner, and M. Kling. 2010. Facies pattern and sea-level dynamics of the early Late Cretaceous transgression: A case study from the lower Danubian Cretaceous Group (Bavaria, southern Germany). Facies 56: 483–507.
Acknowledgments
We thank F.T. Fürsich (Erlangen) for a constructive review and discussions on ichnotaxonomy. R. Winkler (MMG, Dresden) photographed the Saxonian specimens of D. ottoi. This paper is a contribution to DFG project WI 1743/6-1 granted to M.W.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wilmsen, M., Niebuhr, B. The rosetted trace fossil Dactyloidites ottoi (Geinitz, 1849) from the Cenomanian (Upper Cretaceous) of Saxony and Bavaria (Germany): ichnotaxonomic remarks and palaeoenvironmental implications. Paläontol Z 88, 123–138 (2014). https://doi.org/10.1007/s12542-013-0185-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-013-0185-1