
Abstract
The Miocene succession (Allala River Sandstones and Tenes Blue Marls Formation) that crops out in the Tenes area, situated in the northeast of the Lower Chelif Basin in NW-Algeria, contains a low-diversity assemblage of trace fossils. Fifteen (15) ichnogenera were identified: Arenicolites, Beaconites, Cylindrichnus, Diplocraterion, Macaronichnus, Ophiomorpha, Palaeophycus, Parahaentzschelinia, Planolites, Rosselia, Skolithos, Taenidium, Teichichnus, Thalassinoides and Zoophycos. Ethologically, these ichnogenera chiefly display dwelling and feeding activities. The presence of thick, deep-tier, scattered, mainly vertical dwelling burrows attributed to the Skolithos ichnofacies indicates high energy conditions, normal oxygenation and soft substrate. Moreover, elements of the Cruziana ichnofacies show more varied behavioural strategies and higher inchnodiversity with the dominance of horizontal burrows of deposit-feeders. This ichnological study supports the palaeoenvironmental interpretation based on sedimentological analysis of a wave-dominated siliciclastic platform (backshore to offshore), allowing a more precise zonation of the shoreface zone (middle/upper and lower shoreface). In addition, this study allows evaluation of variable degrees of storm influence in response to the contrasting palaeogeomorphology of the coastline.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
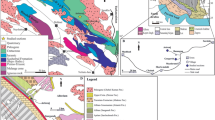
The Tenes area offers outstanding Miocene outcrops, with variable degrees of accessibility, in general attributed to a period of time ranging from the terminal Burdigalian?/Langhian to the Tortonian (Cassan 1968; Lepvrier and Magné 1975; Neurdin-Trescartes 1992; Nemra 2020). These Miocene outcrops belong to the northeastern border of the Lower Chelif Basin, between Dahra mountains in the west and Beni Menacer mountains (also called Bou Maad) in the east (Fig. 1a and c).

Geographical context and geological map of the Tenes area. a large-scale map showing the location of Algeria and a simplified geological map of the central Tellian domain. The red square marks the position of the Tenes area; b geographical location of the studied areas; c geological map of Tenes area (modified from Brives 1913)
Ichnological studies of the Neogene succession of the Lower Chelif Basin are scarce. In the study area, the trace fossils of the Miocene formations have not been investigated in detail. Neurdin-Trescartes (1992) mentioned the presence of burrows in the Allala River Sandstones Formation but without any further taxonomic assignment. Therefore, the systematic description of these trace fossils is one of the aims of this article.
The study of trace fossils is not only restricted to the determination of different ichnotaxa, but also includes the analysis of other related parameters as a valuable tool in sedimentological studies. The most important parameters used in this study are burrow diameter, bioturbation index, relative dominance between vertical in respect to horizontal burrows, ichnodiversity and the inferred behaviour of the producing organism. The studied Miocene succession was attributed to a wave-dominated siliciclastic platform (Nemra 2020); however, a detailed ichnological study allows us to refine the sedimentological interpretation and to assess degrees of storm influence.
Geological setting
The Lower Chelif Basin, located in the external zone of the Tellian Atlas, developed during the Alpine orogeny and constitutes a part of the sublittoral basin of western Algeria (Perrodon 1957; Delteil 1974; Fenet 1975; Belkebir et al. 1996). Geomorphologically, it is a long depression, extending for 350 km in an ENE/WSW trend (Bessedik et al. 2002). In its axial zone, the Miocene formations, stacked essentially above the Cretaceous allochthonous basement, consist of approximately 4000 m of mostly marly deposits (SN Repal 1952; Perrodon 1957). The basement is formed of three distinct geological groups: (1) Triassic formations composed essentially of gypsum, dolomite and iridescent clay; (2) Jurassic limestone of Cape Tenes that forms an integral part of the “Dorsale kabyle” unit (= “chaîne calcaire” of Glangeaud 1932), and (3) Cretaceous-Paleogene sedimentary formations that constitute a major part of the succession (Fig. 1c). At the edges of the basin, an often angular unconformity subdivides the Miocene into two transgressive sedimentary cycles (Perrodon 1957). In the centre of the basin, the lateral equivalent of this discontinuity is a “concordance surface” (Perrodon 1957). These two Miocene cycles are designated by Neurdin-Trescartes (1992) as two transgressive megasequences (MSI and MSII). In the studied area, Miocene rocks are represented by deposits of the first cycle and the lowermost part of the second cycle sensu Perrodon (1957). The first cycle begins with continental deposits evolving to marine deposits. The continental deposits are mainly represented by conglomerates, often reddish in colour. Marine deposits are made up of conglomerates and sandstones containing the remains of marine organisms, bioclastic limestones, sandy marls and generally grey or blue marls (Perrodon 1957; Neurdin-Trescartes 1992; Nemra 2020). The deposits of the lowermost part of the second cycle rest in angular unconformity on first-cycle deposits and they are represented by yellow marls (Neurdin-Trescartes 1992; Nemra 2020).
The Miocene rocks in the Tenes area reach 500 m in thickness (Cassan 1968). The lithostratigraphic study of this Miocene succession has permitted definition of the superposition of two geological formations (Nemra 2020; Fig. 2), the Allala River Sandstones and the Tenes Blue Marls.

The Allala River Sandstones Formation is separated from the basement by an angular unconformity and its top coincides with a lithological change from sandstone to marls. It is essentially constituted by a well-developed succession of light-grey to brown, medium- to very thick-bedded, coarse- to fine-grained sandstone beds that contain sedimentary structures and traces fossils in some levels. These beds are partly separated by sandy marl and gravelly marl interbeds. Locally, in the basal part of this succession, the sandstones pass laterally into conglomerates and biodetritic limestones. Dark-grey, generally metric, sandy marls interrupt these coarse detrital facies locally. The thickness of this formation varies widely, reaching its maximum at Kaïsar outcrop (ca. 250 m) and its minimum at Ounsour Anhas outcrop (31 m).
The Tenes Blue Marls Formation extends from the top of the underlying Allala River Sandstones Formation to a flat discontinuity separating it from the overlying yellow marl unit. It consists essentially of thick, dark grey to blue marls, sometimes crossed by calcite veins. Marls are mostly intercalated by either thin sandstone beds or centimetre levels of indurated silty marls. Synsedimentary slump structures and normal faults have also been observed. It is difficult to estimate the real thickness of this formation. Cassan (1968) reported a total thickness of 250 m, although this thickness is clearly less at the Ounsour Anhas outcrop (ca. 90 m). The upper limit of this formation is only observed at this locality (Neurdin-Trescartes 1992; Nemra 2020). These two formations are attributed to a time interval ranging from the terminal Burdigalian? or early Langhian to the middle Serravallian whereas the yellow marl unit is assigned to the early Tortonian (Cassan 1968; Nemra 2020).
In terms of sequence stratigraphy, the angular unconformity, separating the Miocene succession and the ante-Neogene basement, corresponds to a sequence boundary sensu Vail (1987). This discontinuity is overlain by continental deposits represented by conglomerates and clays, often reddish, belonging mainly to alluvial fan environments (Nemra 2020). These conglomerate-dominated continental deposits correspond to a low system tract (LST sensu Vail 1987). Marine sediments overlie these continental deposits and are represented at Vieux Tenes by stacking of three high-frequency sequences (Nemra 2020), displaying vertical evolution from coarse-grained deposits (conglomerate, sandstone or mixed siliciclatic-carbonate facies) to fine-grained deposits (marls dominated facies). These sequences correspond to a T-cycle sensu Zecchin (2007) and Catuneanu and Zecchin (2013), showing evolution from beachface/shoreface facies to upper/lower offshore facies and they are bounded by ravinement surfaces (Nemra 2020). The stacking sequences show a transgressive trend and represent the transgressive systeme tract (TST sensu Vail 1987).
Material and analytical methods
This study was based on field observations of the Miocene succession in the Tenes area carried out at five different outcrops (Djebel Ksar, Vieux Tenes, Ounsour Anhas, Djebel Ach Zerfet and Kaïsar; Fig. 1b). The upper part of the section at Djebel Ach Zerfet is partially covered; therefore, the description of this part was logged based on cores. The purpose of fieldwork was to collect detailed lithostratigraphic, facies and ichnological data. Lithology, grain size, bed thickness, sedimentary structures (including ichnofabrics), geological contacts and fossil content were logged bed-by-bed at each outcrop. The ichnological features examined at the bed scale were shape, orientation, position within the beds, length, diameter, degree of bioturbation and ichnodiversity. The description of the position of trace fossils within the beds follows the taphonomic classification scheme of Martinsson (1970). The Reineck (1963) and Taylor and Goldring (1993) classifications has been used to evaluate the bioturbation index. Ichnological data gathered from each outcrop are compared to better evaluate the hydrodynamic conditions, degrees of storm influence, oxygenation rate, etc.
Summary of palaeoenvironments
The major lithofacies of the Miocene outcrops in the Tenes area are represented by sandstones, conglomerates, limestones and marls. The detailed analysis of these lithofacies indicates that sedimentation took place on a wave-dominated siliciclastic platform, evolving from a gravel-dominated beachface-shoreface complex to an offshore complex (Fig. 3; Nemra 2020). The detailed sedimentary features of the facies associations are reported in Table 1.

Sedimentological model of the studied deposits (Nemra 2020). The sedimentological interpretations are based on the combination of a gravel beach with microtidal regime model (Bluck 1967; 1999; Massari and Parea 1988; Sanders 2000) and a standard siliciclastic platform model (Reading and Collinson 1996; Buatois and Mángano 2011). Ichnological interpretations are based on field data and the work of MacEachern et al. (2007), Buatois and Mángano (2011), and Pemberton et al. (2012)
The beachface-upper shoreface zone is characterised by conglomerate and sandstone facies with abundant bivalve clasts (Fig. 4f). The conglomerates are either open framework (Fig. 4c, d) or have a sandy matrix (Fig. 4a, b) and may show low-angle cross-stratification. The presence and abundance of coarse cobbles and boulders at Kaïsar outrop indicates close proximity of this site to the sediment source. Furthermore, the lithologic nature of these gravels suggests that they are mainly derived from the erosion of palaeo-cliffs (Cretaceous basement). Sandstone facies (coarse to medium) is represented by massive sandstones and massive pebbly sandstones (Fig. 4b), locally containing scour and fill structures. Episodically, pebbly beds and centimetre- to decimetre-thick conglomerate layers (less than 30 cm) are also intercalated within the sandstones (Fig. 4b).

Photographic illustration of beachface-shoreface facies; Allala River Sandstone at Ounsour Anhas (a, b, d and f), Kaïsar (c), Vieux Tenes (e) and Djebel Ach Zerfet (g-h). a Clast-supported, sandy-matrix conglomerates (Gcs, Gps). Note the imbrication in the Gbs levels and the ravinement basal surface of the Gcs level; b Clast-supported, sandy-matrix conglomerates (Gps) intercalated by massive pebbly sandstones (Sp); c-d Clast-supported, open framework conglomerate (Gco, Gpo) levels with more-or-less imbricated clasts; e bioclastic limestone with Pectinidae fragments; f grain-supported, sandy-matrix conglomerates with Ostrea fragments (yallow arrow); g bioclastic sandy limestone evolving to a calcareous sandstone (positive grading); h detail of sandy bioclastic limestone
The upper to middle shoreface zone includes sandstones, bioclastic limestones (mixed facies; Fig. 4e and g-h), bioclastic sandstones (mixed facies), conglomerates and scarce layers of sandy marl. The dominant facies consists of light-grey to light-brown, very thick to medium bedded, medium- to coarse-grained sandstones with an overall massive appearance and occasional normal grading. Sedimentary structures include current and oscillatory ripples (Fig. 5a and f), planar laminations (Fig. 5a and d), low-angle and high-angle cross-lamination (Fig. 5b). Hummocky cross-stratification sandstones (HCS) are rare, while erosive basal surfaces and amalgamated beds are frequent. Locally, tabular pebbly conglomerates are intercalated within the sandstones (Fig. 5i). The mixed facies (bioclastic limestones, sandy limestones and sandstones with bioclasts) occur at Djebel Ksar, Vieux Tenes and Djebel Ach Zerfet outcrops. They contained a benthic macrofauna characterised by bivalve clasts (often Ostrea), balanids, bryozoans, echinoderm plates, echinoid spines, benthic foraminifera and remnants of red algae. The sedimentary structures (oscillation ripples, planar lamination, low-angle parallel lamination and cross-lamination) preserved in this zone are dominantly wave-generated under both fair-weather and storm conditions. The sediments in this zone were constantly reworked by breaking waves, surf and swash-zone processes. Storm-wave influence is recorded as HCS, erosion surfaces, scour and fill structures (Fig. 5e), normally graded beds and tabular conglomerate beds. In general, the benthic fauna recorded in the mixed facies confirms an agitated shallow-marine environment. The fauna (Ostrea, balanids, bryozoans and echinoderms) suggests hard substrates such as gravels, shell clasts, etc.

Photographic illustration of shoreface/offshore facies; Allala River Sandstone (a-f, i) and Tenes Blue Marls (g-h); a wave-ripple cross-laminated sandstones overlain by horizontal laminated sandstones (Djebel Ksar); b cross-stratified sandstones (Vieux Tenes); c hummocky cross-stratified sandstones “HCS” (Vieux Tenes); d ripple cross-laminations (black arrows) and horizontal laminated sandstones (Vieux Tenes); e scour structure filled with coarse lithic sands and mud chips (Djebel Ksar); f sandstone surface with wave-ripples (Djebel Ksar); g and h sandstone intercalated within marls showing low angle cross laminations (g) and ravinement surface (h) interpreted as tempestites (Vieux Tenes); i conglomerates with a ravinement surface and positive grading intercalated within sandstones (Ounsour Anhas)
The lower shoreface zone consists essentially of thin- to medium-bedded, fine- to medium-grained sandstones interbeded with thin layers of silty/sandy marls. Sandstone facies includes massive and rarely graded sandstones; planar, undulated and rarely oblique-laminated sandstones; as well as HCS (Fig. 5c). Current and oscillatory ripples occur at the top of some massive sandstone beds. The rare amalgamated beds and the presence of thin marl layers indicate a relatively deeper environment of deposition. The marly interbeds are a record of fair-weather conditions above the storm wave base (Pemberton et al. 1992).
The offshore transition to upper offshore zone is dominated by very thick, dark grey to blue marls with intercalations of sandstones (Fig. 5g, h). These sandstones are light grey, erosively based, often very thin to medium bedded (1 to 25 cm), rarely very thick (up to 2 m at Ounsour Anhas outcrop), very fine-grained, containing parallel laminations, low-angle cross-laminations (Fig. 5g), HCS and normal graded beds. Very thin (up to 5 cm), very fine-grained sandy to silty marl lenses occur locally. These facies record environmental changes related to an episodic sedimentation. The marl intervals essentially represent suspended sediment fall-out in a low-energy setting. The sandstone intercalations indicate alternation of lower-energy storm events between fair-weather wave base and storm wave base.
The lower offshore to distal shelf zone is represented by very thick, dark grey to blue marls intercalated locally by sandy to silty marl lenses. Marls of this zone mostly involve suspension fall-out in low-energy setting. The very rare sandy to silty marl lenses are interpreted as distal tempestites.
Systematic description of trace fossils
This part is devoted to the ichnological study of the Miocene of the Tenes region. The inventory shows fifteen (15) ichnogenera. These trace fossils have been analysed on well-exposed bedding planes and vertical sections at the different investigated outcrops (Vieux Tenes, Djebel Ksar, Kaïsar, Djebel Ach Zerfet and Ounsour Anhas). Figures 6, 7, 8, 9, 10a, b, and 11 show the distribution and trace fossils-bearing facies. No trace fossils have been observed in the conglomeratic facies. The preservation of trace fossils is, locally, very poor due to diagenetic factors and weathering/erosion obliteration, thus, their determination in some beds remains tentative.


Lithological log and trace fossils distribution at Ounsour Anhas outcrops

Lithological log and trace fossils distribution at Djebel Ksar outcrops

Lithological log and trace fossils distribution at Vieux Tenes outcrops


Lithological logs and trace fossils distribution at Kaïsar outcrops. a Kaïsar 1 section; b Kaïsar 2 section

Lithological log and trace fossils distribution at Djebel Ach Zerfet outcrops
Arenicolites Salter, 1857
Arenicolites isp.
(Fig. 12a and e)

Photographic illustration of Arenicolites, Diplocraterion, Ophiomorpha (Op); endichnia within Allala River sandstone. a Arenicolites (cross-section; Djebel Ksar); b small Arenicolites or Diplocraterion? (bedding plane; Vieux Tenes); c Diplocraterion parallelum (cross section; Djebel Ach Zerfet); d Ophiomorpha and Diplocraterion (bedding plane; Ounsour Anhas); e small Arenicolites (bedding plane; Ounsour Anhas); f detail of (d) showing Ophiomorpha burrows; g detail of (d) showing Diplocraterion and Ophiomorpha with weakly developed pellets; h Diplocraterion (bedding plane; Ounsour Anhas)
Paired circular openings corresponding to U-shaped burrows without a spreite occur on upper sandstone bedding planes. In the Allala River Sandstone Formation, they are preserved as endichnia. Based on the size of the burrows, two types can be distinguished: (1) large Arenicolites (Djebel Ksar and Djebel Ach Zerfet; Fig. 12a) with a distance of 10 cm between the two circular openings and a tube diameter of 1.5 cm; and (2) small Arenicolites, with a few millimetres wide burrow apertures and a distance of less than 2 mm between the two tubes (Vieux Tenes and Ounsour Anhas; Fig. 12b and e; respectively).
Most Arenicolites correspond to a dwelling trace of suspension-feeding organisms, such as polychaete worms or amphipod crustaceans (Bradshaw 2010; Buatois and Mángano 2011; Hofmann et al. 2012; Knaust 2017). They are common in shallow-marine (Fillion and Pickerill 1984; Gingras et al. 1999; Gerard and Bromley 2008; Bradshaw 2010; Knaust 2017), continental (Ekdale et al. 1984; Ekdale and Bromley 2012; Scott et al. 2012), and deep–marine environments (Crimes et al. 1981; Uchman and Wetzel 2012). Arenicolites is typically associated with high-energy deposits (e.g. storm deposits) or migrating sand bodies (e.g. sand dunes) in an upper to middle shoreface environment (Bradshaw 2010; Pemberton et al. 2012; Knaust 2017).
Beaconites (Vialov, 1962)
Beaconites capronus (Howard and Frey, 1984)
(Fig. 13a)

Photographic illustration of Beaconites and Cylindrichnus (Allala River Sandstone). The scale bar = 5 cm. a Beaconites capronus (Vieux Tenes; bedding plane); the close-up photograph illustrates the meniscus filling; b-d Cylindrichnus concentricus in cross section at Djebel Ksar (b) and Vieux Tenes (c and d)
This ichnospecies corresponds to straight to meandering, thinly lined, cylindrical to sub-cylindrical burrows, parallel to the bedding planes, with a chevron-shaped meniscus fill. It occurs as epichnia within the Allala River Sandstone Formation at Djebel Ksar and Vieux Tenes. The arcuate menisci are heterogeneous, regularly arranged and represented by an alternation of fine- and coarse-grained sediment segments. The diameter of the burrow ranges from 0.6 to 1 cm, and the preserved length can reach 40 cm.
Beaconites differs from other meniscate burrows (Taenidium and Ancorichnus) by a lining (Keighley and Pickerill 1995; Boyd and McIlroy 2017). Beaconites capronus is characterised by chevron-shaped meniscae and a thin lining (Howard and Frey, 1984; Boyd and McIlroy 2017). It is interpreted as a deposit-feeding vermiform organism that processed large amounts of sediment (Boyd and McIlroy 2017) and is common in shallow-marine environments (Howard and Frey, 1984; Boyd and McIlroy 2017). The tracemakers of meniscate burrows, such as Beaconites, progress horizontally through the sediments and deposit backfill sediments behind them as they advance (Frey et al. 1984; Boyd and McIlroy 2017). The direction of movement is indicated by the concave side of the menisci (Frey et al. 1984).
Cylindrichnus Toots in Howard, 1966
Cylindrichnus concentricus
(Fig. 13b-d)
This ichnospecies comprises a cylindrical, arcuate to J-shaped burrow with a funnel-like aperture, concentric lining and a passive fill. The diameter of the burrow decreases from top to bottom (from 1 cm to a few millimetres), the length of the burrow varies between 10 and 20 cm. The massive sandstone fill is identical to the host rock. These burrows are preserved as endichnia within decimetre-thick sandstone beds of the Allala River Sandstone Formation at Djebel Ksar (Fig. 13b) and Vieux Tenes (Fig. 13c, d).
Cylindrichnus is a dwelling trace of detritus-feeding organisms, probably produced by terebellid polychaetes (Goldring 1996; MacEachern and Gingras 2007; Buatois and Mángano 2011; Gingras and MacEachern 2012; Belaústegui and de Gibert 2013; Tonkin 2012; Knaust 2021). It is documented mainly from marine settings (lower shoreface to shelf), as well as marginal-marine, estuarine, delta-front to prodelta environments (Fürsich 1974; Pemberton and Frey 1984; Frey and Howard 1985). In general, Cylindrichnus is recorded in moderate to low-energy facies (Fürsich 1974), although it has also been assigned to high-energy regimes (Pemberton and Frey 1984; Frey and Howard 1985).
Diplocraterion Torell, 1870
Diplocraterion parallelum Torell, 1870
(Fig. 12c, d and g, h)
This trace was recorded as endichnia and in association with frequent Ophiomorpha within the Allala River Sandstone Formation at Ounsour Anhas. On the upper surface of sandstone beds, it appears as paired circular openings connected by a lamellar structure (spreite). The diameter of the apertures is variable (0.4 to 1 cm) and the distance between them ranges from 2 to 12 cm. Vertical U-shaped burrows are composed of two straight and parallel cylindrical tubes connected by a lamellar structure (spreiten). Only one specimen was observed at Djebel Ach Zerfet within Allala River Sandstone Formation. The tube diameter reaches 2 cm, and the burrow depth is about 12 cm.
Diplocraterion differs from Arenicolites by the presence of a spreite and from Rhizocorallium by its vertical orientation (Fleming 1973; Cornish 1986). It is a dwelling burrow of suspension-feeding organisms (Cornish 1986; Hofmann et al. 2012; Zhang et al. 2017). The laminae (spreite) correspond to remnants of the burrow wall reflecting the movement of the tracemaker through the sediment (Cornish 1986). This trace is characteristic of unstable substrates (moving sands) generally reported in high-energy environments from intertidal to proximal subtidal (foreshore to shoreface) (Fürsich 1975; Cornish 1986; Zhang et al. 2017).
Macaronichnus Clifton and Thompson, 1978
Macaronichnus segregatis Clifton and Thompson, 1978
(Fig. 14c-e)

Photographic illustration of Planolites; Macaronichnus; Palaeophycus and Parahaentzschelinia; Allala River Sandstone (a, c-h) and Tenes Blue Marls (b). The scale bar = 5 cm. a Planolites beverleyensis (endichnia and exichnia; cross-section at Vieux Tenes); b Planolites isp. (Plan view in core; endichnia; Djebel Ach Zerfet); c Palaeophycus tubularis and Macaronichnus segregatis (endichnia and exichnia; cross-section at Djebel Ksar); d-e Macaronichnus segregatis in cross-section (d) and bedding plane (e) at Vieux Tenes and Djebel Ksar, respectively; f Palaeophycus tubularis (cross-section; Djebel Ksar); g-h Parahaentzschelinia surlyki (endichnia; vertical section at Ounsour Anhas)
Dense, thin, horizontal to subhorizontal, cylindrical burrows with a circular cross-section, characterised by a distinct dark lining and a relatively clear massive fill. They are slightly sinuous, unbranched, although interpenetrations are common. The size of the burrows is variable and ranges from 0.2 to 0.5 cm in diameter and up to 6 cm in length. They occur as endichnia within the Allala River Sandstone Formation at Djebel Ksar (Fig. 14c and e) and Vieux Tenes (Fig. 14d).
These burrows are common in foreshore and shoreface deposits (Głuszek 1998; Knaust 2004; Seike et al. 2011). They are produced by polychaete worms that feed on bacteria and organic matter on the sand-grain surfaces (Clifton and Thompson 1978; Knaust 2004; Seike et al. 2011).
Ophiomorpha Lundgren, 1891
Ophiomorpha irregulare Frey et al., 1978
(Figs. 13d and f, g and 15a-d and f-j)

Photographic illustration of Ophiomorpha (Op) and Rosselia (Ro) within Allala River Sandstone (vertical section). The scale bar = 5 cm. a Ophiomorpha irregulare (endichnia; Djebel Ksar); b horizontally laminated sandstone bed (Djebel Ksar) with a basal ravinement surface containing endichnial vertical burrows of Ophiomorpha irregular and Rosselia erecta; c detail of (b) showing the walls of Ophiomorpha irregulare with pellets (preserved as a cast); d Ophiomorpha irregulare (endichnia; Djebel Ksar); e detail of (b) showing Rosselia erecta; f Ophiomorpha irregulare with a poorly developed pellets (endichnia; Djebel Ach Zerfet); g Ophiomorpha irregulare (endichnia; Kaïsar); h-i Ophiomorpha irregulare (endichnia; Ounsour Anhas); j horizontal burrows of Ophiomorpha irregulaire (endichnia and exichnia; Vieux Tenes)
Ophiomorpha was observed in the sandstones of the Allala River Sandstone Formation as endichnia at Djebel Ksar (Fig. 15a-d), Ounsour Anhas (Figs. 13d and f, g and 15h, i), Kaïsar (Fig. 15g), Djebel Ach Zerfet (Fig. 15f) and Vieux Tenes (Fig. 15j ). It is vertical to subvertical, cylindrical, occasionally horizontal, with sandstone filling identical to the host rock. This ichnospecies is characterised by walls lined with sparse, irregularly distributed, flame-like, mastoid, conical or ovoid pellets. The diameter of the burrow varies between 0.8 and 2.5 cm and its length is between 15 and 30 cm.
Ophiomorpha irregulare is interpreted to be produced by decapod crustaceans (e.g. shrimps, Callianassidae; Bromley and Ekdale 1998; Buatois and Mángano 2011). It is regarded as a dwelling trace of suspensivorous organisms (Frey et al. 1978), or a feeding trace of a detritivorous tracemaker (Frey and Howard 1990; Bromley and Ekdale 1998). Ophiomorpha is commonly reported from pure, homogeneous, or laminated sandstone deposits (Pollard et al. 1993; Nagy et al. 2016; Knaust 2017), where it has been considered as an important component of high-energy, shallow marine facies (Frey et al. 1978; Pollard et al. 1993; Uchman and Demircan 1999; Bendella et al. 2011; Uchman and Wetzel 2012; Nagy et al. 2016). The pellets of Ophiomorpha originated as a mixture of mucus (matrix) and sand grains (Frey et al. 1978) and were used to support the burrow wall to protect it from collapse (Buatois and Mángano 2011).
Palaeophycus Hall, 1847
Palaeophycus tubularis Hall, 1852
(Fig. 14c and f)
A horizontal, straight to slightly curved, cylindrical to subcylindrical, thinly lined and unornamented burrow. Its length is up to 30 cm and the diameter ranges from 0.8 to 1 cm. It is preserved as endichnia and exichnia within the Allala River Sandstone Formation at Djebel Ksar and as epichnia in silty limestone beds of the Tenes Blue Marls Formation at Vieux Tenes.
Palaeophycus is characterised by a lined wall and a burrow fill similar to the host rock (Pemberton and Frey 1982; Keighley and Pickerill 1995). It is classified as a feeding and dwelling trace (Pemberton and Frey 1982; Lucas and Lerner 2006; Virtasalo et al. 2006). Multiple marine suspension/detritus feeders or predacious organisms can generate this trace, such as annelids, arthropods and polychaetes (Pemberton and Frey 1982; Buatois and Mángano 1993; Knaust 2017), whereas in continental environments, it is probably produced by insects and other arthropods (Ekdale et al. 1984; Buatois and Mángano 1993). In marine environments, P. tubularis is considered as eurybathic (McCann 1988; Uchman 1995, 1998; Frey and Howard 1990; Lucas and Lerner 2006; Gani et al. 2007; Tiwari et al. 2011). In shallow-marine environments, the occurrence of Palaeophycus implies a well-oxygenated environment and abundant nutrients (Tiwari et al. 2011).
Parahaentzschelinia Chamberlain, 1971
Parahaentzschelinia surlyki Dam, 1990
(Fig. 14g, h)
This trace is rare and was oserved only at Ounsour Anhas as endichnia within parallel-laminated sandstone of the Allala River Sandstone. It consists of an overall funnel-shaped, vertical to inclined burrow, with a principal central tube. The upper funnel shaped part is irregularly disturbed by incorporated mud and may show a mud-lined, passively sand-filled tube. The lower part of the main tube appears as ovoid. The trace length is about 10 cm, the diameter of the cylindrical tube can reach 1.5 cm, while the diameter of the preserved funnel-shaped upper part is about 40 cm.
Parahaentzschelinia is mainly regarded as feeding activity of a tellinid bivalve (Fürsich et al. 2006; Knaust 2015). It is recorded in deep-marine deposits (Uchman 1995; Wetzel 2008), however, this trace is common in high-energy shallow- to marginal-marine settings such as foreshore, shoreface, tidal flat and tide-dominated delta (Dam 1990; Bann and Fielding 2004; Mángano and Buatois 2004; Fürsich et al. 2006; McIlroy 2007).
Planolites Nicholson, 1873
?Planolites beverleyensis Billings, 1862
(Fig. 14a)
This trace is rare, observed only at Vieux Tenes, where it is preserved as epichnia as well as exichnia within the Allala River Sandstone Formation. It is a cylindrical, horizontal to subhorizontal, cylindrical burrow. In cross-section, it appears circular to subcircular with a diameter of 0.5 cm and a length up to 30 cm.
Planolites isp.
(Fig. 14b)
This form is rare and was observed only in a core sample of the lowermost part of the Tenes Blue Marls Formation at Djebel Ach Zerfet. The burrows are small, unbranched, horizontal to slightly inclined, cylindrical to subcylindrical, straight, curved, or contorted. It is preserved as endichnia within centimetre- to decimetre-thick, light-grey sandy to gravely marl intercalations with grading. The burrow fill consists of dark-grey marl without any observed internal structures. The appearent length of the burrow is up to 5 cm and the diameter is about 4 mm.
Planolites differs from Palaeophycus by its infill, which is different from the host rock (colour, nature of the sediment) and the lack of lining (Pemberton and Frey 1982; Keighley and Pickerill 1995). It is regarded as a feeding trace of invertebrate deposit-feeders such as worms, insects, crustaceans and molluscs (e.g. bivalves) (Pemberton and Frey 1982; Frey and Howard 1985; Buatois and Mángano 1993; Uchman 1995; Knaust 2017). Planolites is considered as ‘facies-crossing’, occurring in a variety of depositional environments (Pieńkowski 1985; Buatois et al. 2001; Bendella and Ouali Mehadji 2014). It has been reported in continental environments, where it can be produced by insects (Buatois and Mángano 1993; Uchman et al. 2004; Ekdale et al. 2007), and in marine environments of varying depth (McCann 1988; Uchman 1995; 1998; Riahi 2014; Bendella and Ouali Mehadji 2014).
Rosselia Dahmer, 1937
Rosselia erecta (Torell, 1870)

Photographic illustration of Rosselia, Taenidium and Teichichnus within Allala River sandstone. a-b Rosselia erecta (endichnia; transversal section at Vieux Tenes); c-d Taenidium barretti (epichnia; bedding plane at Djebel Ksar); e-f Teichichnus; endichnia; cross-section at Vieux Tenes (e) and Ounsour Anhas (f)
Rosselia erecta was observed as endichnia within the Allala River Sandstone Formation at Djebel Ksar (Fig. 15b and e) and Vieux Tenes (Fig. 16a, b). It consists of a vertical to subvertical, fusiform (bulbous or spindle-like shape), unbranched burrow with a circular to ovoid, funnel-shaped aperture. The cylindrical tube occupies a major part of the funnel. Below the bulbous part, the burrow gradually narrows downward. The overall length of the burrow does not exceed 20 cm, the diameter varies between 1.5 and 3 cm for the bulbous part, whereas the diameter of the internal tube ranges from 0.3 to 1 cm.
Rosselia erecta corresponds to a dwelling trace produced by suspension-feeding organisms attributed to polychaetes and probably Cerianthidae (Anthozoa) (Knaust 2021), inhabiting long tubes within marine substrate (Uchman and Krenmayr 1995). It is often reported from marginal- to shallow-marine environments (e.g. shoreface; Knaust 2021).
Skolithos Haldeman, 1840
?Skolithos annulatus (Howell, 1957)
(Fig. 17c-g)

Photographic illustration of Skolithos; vertical sections at Allala River Sandstone (a, b, e, f, h, i) and Tenes Blue Marls (c, d, g). The scale bar = 5 cm. a, b Skolithos linearis (endichnia; Djebel Ach Zerfet). Note in (a) the abnormally thick lining (mantle); c ?Skolithos annulatus (endichnia; Ounsour Anhas); d ?Skolithos annulatus (exichnia; Ounsour Anhas); e ?Skolithos annulatus (endichnia; Kaïsar). Note the vertical, thick, annulated, indeterminate burrow (IT) showing decrease in diameter downward; f detail of (e) showing an annulated fill of ?Skolithos annulatus; g ?Skolithos annulatus (endichnia; Ounsour Anhas); h Skolithos isp. (endichnia; Ounsour Anhas); i Skolithos linearis (endichnia; Djebel Ksar)
?Skolithos annulatus is characterised by a vertical to inclined, straight, annular burrow. The length of the burrow varies between 10 and 30 cm and the diameter can reach 2 cm. The rings are 0.8 to 2.5 cm in thickness. This ichnospecies was identified as (1) endichnia at Kaïsar (Fig. 17e, f) within the Allala River Sandstones and at Ounsour Anhas (Fig. 17d) within sandstone intercalation of the Tenes Blue Marls Formation; (2) exichnia at Ounsour Anhas (Fig. 17c and g) within the Tenes Blue Marls Formation.
Skolithos linearis Haldeman, 1840
(Fig. 17a, b and i)
This ichnospecies was identified as endichnia at Djebel Ach Zerfet, Djebel Ksar (Allala River Sandstones) and Ounsour Anhas (Tenes Blue Marls Formation). It is distinguished from other Skolithos ichnospecies by a straight to slightly curved burrow with homogeneous fill, and a very high length/diameter ratio (Knaust et al. 2018). The length of the burrow can reach 40 cm and the diameter is generally between 0.2 and 0.6 cm.
In marine sediments, Skolithos corresponds to dwelling traces produced by suspension- or deposit-feeders or organisms that exhibit passive predation strategies (e.g. annelids, phoronids and crustaceans) (Schlirf and Uchman 2005; Buatois and Mángano 2011; Adserà 2018). Skolithos indicates relatively high-energy, shallow-water conditions and occurs generally in various shallow-marine environments (Pieńkowski 1985; Alpert 1974; Fillion and Pickerill 1984; Schlirf and Uchman 2005). However, this trace has also been reported from continental environments where it is probably produced by insects and spiders (Schlirf and Uchman 2005), and in deep-marine environments (Uchman 1995), where it can be produced by aplacophores (mollusks) and holothurians (Dashtgard and Gingras 2012).
Taenidium Heer, 1877
Taenidium barretti (Bradshaw, 1981)
(Fig. 16c, d)
This ichnospecies is composed of straight to meandering, cylindrical to sub-cylindrical burrows, parallel to the bedding planes, with a meniscate fill. It occurs as epichnia within the Allala River Sandstone Formation at Djebel Ksar. The menisci are represented by heterogeneous, short and arcuate segments. The diameter of the burrow is less than 1 cm, and the apparent length can reach 40 cm.
Taenidium corresponds to deposit-feeding trace (Hammersburg et al. 2018). In marine environments, Taenidium may be produced by arthropods or worm-like organisms (Knaust 2017). It is a eurybathic trace that has been reported in both shallow-marine (middle to lower shoreface; Frey and Howard 1990; Sarkar et al. 2009) and deep-marine environments (MacDonald 1982; Crimes 1992), as well as in aeolian (Ekdale et al. 2007) and fluvial environments (Uchman et al. 2004). It is commonly found in marine environments in the Cruziana ichnofacies (Gerard and Bromley 2008; Sarkar et al. 2009; Bendella et al. 2011; Knaust 2017). Taenidium is considered to be produced by an animal which moves horizontally through the sediment and progressively deposits alternating backfill sediments (Bromley et al. 1999; Sarkar et al. 2009).
Teichichnus Seilacher, 1955
Teichichnus ?zigzag Frey and Bromley, 1985
(Fig. 16e)
One specimen was observed as endichnia within the Allala River Sandstone at Vieux Tenes that is associated with Thalassinoides. In cross-section, it appears as patches of concave-up spreiten vertical to the bedding plan. The width of the burrow is about 5 cm and and spreiten become wider downward to reach 2.5 cm in diameter.
Teichichnus isp.
(Fig. 16f)
This ichnospecies is rare and was recorded in association with Ophiomorpha as endichnia within the Allala River Sandstone at Ounsour Anhas. In cross-section, the observed specimen shows patches of vertical concave downwards spreiten. The basal part is the widest (3 cm) and shows a circular cross-section of the passively filled burrow that contrasts with the actively filled spreite. The depth of the observed burrow reaches 12 cm.
Teichichnus is interpreted as produced by worm-like animals (e.g. annelids) and arthropods (e.g. trilobites, crustaceans) (Knaust 2018a, 2018b; Vinn and Toom 2018). Since this trace fossil was found in association with Thalassinoides and Ophiomorpha, its tracemaker is assumed to be a crustacean. Different behaviours of the producing organims can be inferred, including dwelling, deposit- and suspension-feeding, as well as gardening (Knaust 2018a). Teichichnus is commonly reported from marginal-marine to shallow-marine environments (shoreface to offshore, tidal flat) with stressed conditions (i.e. salinity and oxygen fluctuations, sedimentation rate, erosional events). However, it is also mentioned in other environments such as deep-marine settings (Uchman and Wetzel 2012) and outer shelf or distal epeiric basin (Frey and Bromley 1985; Locklair and Savrda 1998). It is considered as equilibrium trace as indicated by stacked laminae that result from the movement of the producing organism in response to different sedimentation and colonization events (Knaust 2017, 2018a).
Thalassinoides Ehrenberg, 1944
Thalassinoides paradoxicus Kennedy, 1967
(Fig. 18b)

Photographic illustration of Thalassinoides; Allala River sandstone. The scale bar = 10 cm. a thick nodular Thalassinoides suevicus (endichnia; vertical section at Kaïsar); b dense three-dimensional Thalassinoides paradoxicus boxworks (endichnia; vertical section at Ounsour Anhas); c-d Thalassinoides suevicus at Kaïsar (c) and Ounsour Anhas (d) (hypichnia; bedding plane); e dense bidimensional boxwork of Thalassinoides suevicus (hypichnia; bedding plane at Kaïsar); f bidimensional boxwork of Thalassinoides suevicus (epichnia; bedding plane at Vieux Tenes); g-h Thalassinoides shaft (endichnia) at Djebel Ksar (g) and Kaïsar (h); i isolated Y-shaped borrow of Thalassinoides suevicus (hypichnia; bedding plane at Kaïsar)
This inchnospecies shows a dense and irregular three-dimensional boxwork. The burrows are preserved as endichnia within the Allala River Sandstone Formation at Ounsour Anhas. They illustrate both "T" and "Y" shaped junctions. The tunnels are curved to straight and oriented at various angles. The diameter of the burrow ranges from 0.5 to 1.5 cm.
Thalassinoides suevicus (Rieth, 1932)
(Fig. 18a and c-i)
These burrows are cylindrical to subcylindrical, horizontally and/or vertically branched with a sandy fill and a smooth surface wall. They often bifurcate into "Y" or "T" shapes with an increase in diameter at the branching points to form irregular bulges and swellings. They are conserved within some sandstone beds of both Allala River Sandstone and Tenes Blue Marls Formations. Three configurations have been observed: (1) isolated burrows conserved as hypichnia at Kaïsar (Fig. 18c and i); (2) two-dimensional networks observed as epichnia, hypichnia or endichnia at Vieux Tenes (Fig. 18f), Kaïsar (Fig. 18e) and Ounsour Anhas (Fig. 18d); (3) three-dimensional boxworks observed as endichnia at Kaïsar (Fig. 18a). The diameter of the burrows is widely variable and ranges from 1 to 2 cm, whereas the length of the burrows varies from 10 to ca. 50 cm. The lowermost part of the Allala River Sandstone Formation contains vertical shafts that can reach 1.5 cm in diameter and 50 cm in length at Kaïsar (Fig. 18h) and Djebel Ksar (Fig. 18g). In the lower part of the Allala River Sandstones Formation (Kaïsar), thick Thalassinoides burrows (Fig. 18a) are preserved as endichnia, which diameter can reach 8 cm at swelling junctions. They are white coloured, calcite cemented with nodular aspect (individual burrows are visible) or completely nodular (individual burrows are hardly visible).
Thalassinoides is commonly recorded in shallow-marine environments (Frey and Howard 1990; El-Sabbagh et al. 2017), although it also occurs in deep-marine environments (Uchman 1995). Many of these dwelling and feeding burrows can be attributed to the activity of decapod crustaceans (Bromley and Frey 1974). However, these trace fossils have been reported in lower Palaeozoic rocks prior to the appearance of thalassinids and callianasids, where other arthropods likely produced similar patterns (Buatois and Mángano 2011). Thalassinoides tends to display a higher proportion of vertical components under high energy conditions (Buatois and Mángano 2011), as it is the case in the sandstones of the Kaïsar or Ounsour Anhas (Allala River Sandstone Formation). Thalassinoides is often considered as an indicator of oxic conditions (El-Sabbagh et al. 2017).
Zoophycos Massalongo, 1855
Two different morphotypes of Zoophycos have been observed.
Zoophycos isp. A
(Fig. 19a-f)

Photographic illustration of Zoophycos (bedding planes; endichnia). The scale bar = 10 cm. a-f Zoophycos isp. A within sandstone intercalations of the Tenes Blue Marls at Ounsour Anhas (a-d) and Allala River sandstone at Djebel Ksar (e-f). Note the bundles of minor laminae (ml) bounded by major laminae (ML); Gf: fine sandstone; Gfa: very fine clayey sandstone; g-h) Zoophycos isp. B (sandstone intercalation of the Tenes Blue marls at Ounsour Anhas)
This morphotype consists of an unbranched spiral of a variable width between 30 and 100 cm and appears in form of a flattened cone (conical skirt-like zone; Bromley and Hanken 2003). It is composed of radiating lobe beams displaying small minor sigmoid lamellae (spreiten) bounded on both sides by major radial lamellae. The beams of minor lamellae emanate from a central cone (or an axial point in the middle of the trace, called "apex") and gradually widen towards the marginal edges of the trace, until reaching 4 cm in width. This trace is found at certain sandstone horizons, parallel to the bedding planes, as endichnia (just a few centimetres below the upper surface) within the Allala River Sandstone (Djebel Ksar; Fig. 19e, f and the Tenes Blue Marls Formations (Ounsour Anhas; Fig. 19a-d). Zoophycos isp. A shows some similarities with Zoophycos rhodensis, but the conical shape is more flattened and the J-shaped marginal tube is absent. Moreover, this trace fossil is also similar to Echinospira pauciradiata, but lacks a U-shaped burrow.
Zoophycos isp. B
(Fig. 19g, h)
This morphotype is distinguished from the previous one by a lack of minor lamellae. It is incompletely preserved as endichnia in a sandstone bed of the Tenes Blue Marls Formation at Ounsour Anhas. The central part of the trace (apex) is not preserved, and the remaining part is about 35 cm wide and shows a lobe composed of parallel to subparallel arching ridges (or lamellae) forming a broad "U" shape. The ridges (or lamellae) are tightly spaced, about 1 to 4 mm apart.
Zoophycos are feeding traces of worm-like animals, they are indicative of the ultimate stage of organic matter exploitation in seafloor sediments (Olivero and Gaillard 1996, 2007; Zhang et al. 2015). However, several behavioural strategies have been proposed, such as: feeding, gardening, resting, excreting, cache and refuse dump (Bromley 1991; Zhang et al. 2015). Ekdale and Lewis (1991) and Uchman and Demircan (1999) suggest for the formation of Echinospira (belonging to the Zoophycos group) a burrow system resulting from the successive lateral movement of a causative "U" shaped burrow. Bromley and Hanken (2003) proposedan active fill and a causative J-shaped burrow, based on the marginal continuous tube around the lobate perimeter of Pliocene Zoophycos rhodensis from Rhodes (Greece). Both forms (J-shaped and U-shaped) were collected in sediment cores sampled in the north-western African continental slope (Wetzel and Werner 1981). Zoophycos is probably produced by sipunculid worms, while exploiting nutrients deeply stored in the sediment (Olivero and Gaillard 1996, 2007; Gaillard et al. 1999; Olivero 2003). The representative palaeoenvironment of Zoophycos has shifted from shelf to deep sea over geological time (Bottjer et al. 1988; Olivero and Gaillard 1996; Zhang et al. 2015; Uchman et al. 2016).
Discussion
Ethology and depositional conditions
The nature of the substrate and environmental conditions (agitation, oxygenation, salinity, etc.) control the distribution, density, diversity and behaviours of trace fossil-producing organisms. This makes trace fossils very useful in sedimentological and palaeoenvironmental analyses (Pemberton et al. 2001, 2012; MacEachern and Bann 2008). Based on the sedimentologic analyses (Table 1), the palaeoenvironment of the Miocene succession at Tenes area has been interpreted as a wave-dominated siliciclastic platform that shows evolution from the gravel-dominated beachface-shoreface complex to offshore complex (Nemra 2020). The different ichnotaxa described above are consistent with this interpretation and can be grouped into two different ichnofacies: the Skolithos ichnofacies and the Cruziana ichnofacies. In addition, the comparison between the trace fossils of the five considered outcrops provides information on the depositional environment and the degree of influence of storm waves.
The facies related to the shoreface zone (Table 1) are characterised by the dominance of vertical burrows, the scarcity of trace fossils and a bioturbation index generally between 0 and 1. However, bioturbation index can reach 2 to 3 at (1) the base of Kaïsar sections in some sandstone intervalsbearing robust three-dimensional networks of Thalassinoides burrows; (2) Ounsour Anhas in some sandstone levels with Diplocraterion (Fig. 12d and g, h), Ophiomorpha (Fig. 12d) and other indeterminate traces. Occasionally, at Ounsour Anhas, some sandstone intervals show a strong bioturbation (BI: 5 to 6) with dense three-dimensional Thalassinoides boxwork (Fig. 18b). The ichnodiversity is also very low throughout the entire section.
In the Kaïsar area, three ichnogenera are identified: Thalassinoides (Fig. 18a and h, i) and, more rarely, Skolithos (Fig. 17e, f) and Ophiomorpha (Fig. 15g). Thalassinoides burrows are characterised by a large diameter and often occur in three-dimensional systems with vertical shafts, except for those (two-dimensional networks) observed at the base of the channelized beds intercalated within the sandy marls at the top of the section (offshore-related facies). In the Ounsour Anhas area, Ophiomorpha (Fig. 15h, i), Diplocraterion (Fig. 12g-h), Teichichnus (Fig. 16f) Skolithos, Parahaentzschelinia (Fig. 14g, h) and Arenicolites (Fig. 12e) have been observed. Diplocraterion and Teichichnus are produced by organisms that tolerate a decrease in salinity (Knaust 2017, 2018b). At Djebel Ach Zerfet, the bioturbation is represented by deep vertical burrows of Skolithos (Fig. 17a, b) and more rarely Ophiomorpha (Fig. 15f) and Diplocraterion (Fig. 12c). Horizontal burrows are rarely found and are represented by Thalassinoides and Palaeophycus. The upper to middle shoreface sandstones at Djebel Ksar contain Thalassinoides (including shafts; Fig. 18g), Skolithos (Fig. 17i), Ophiomorpha (Fig. 15a-d), Diplocraterion and Arenicolites (Fig. 12a).
The scarcity of traces and the dominance of vertical dwelling burrows of suspension-feeding organisms in these areas (Kaïsar, Ounsour Anhas and Djebel Ach Zerfet), as well as the low ichnodiversity there are characteristics of an agitated environment with frequent high-energy storm events, where oscillating and unidirectional currents maintain nutrient particles in suspension, and thus promote a strategy of suspension feeding behaviour (Pollard et al. 1993; Bendella et al. 2011; Bendella 2012; Rajkonwar et al. 2014; Zhang et al. 2017). Furthermore, the occurrence of equilibrium traces (Diplocraterion and Teichichnus) in sandstones of the Ounsour Anhas area, that might be produced by organisms that tolerate a decrease in salinity (Knaust 2017, 2018b), indicates the recurrence of erosive events and periods of sedimentation (Goldring 1964). Moreover, the large diameters of Thalassinoides burrows (up to 4 cm in diameter at Kaïsar) and the high proportion of vertical components (Thalassinoides shafts) also provide more evidence of an oxygenated environment and high-energy currents (Buatois and Mángano 2011; El-Sabbagh et al. 2017).
In the upper part of the upper sandstone member at Djebel Ksar, the bioturbation remains quite rare, with a moderate ichnodiversity containing Ophiomorpha (Fig. 15a-d), Cylindrichnus (Fig. 13b), Rosselia (Fig. 15b and e), Skolithos and Taenidium. The occurrence of rare elements of the Cruziana ichnofacies (Rosselia and Taenidium; Knaust 2017), in association with elements of the Skolithos ichnofacies (Ophiomorpha, Cylindrichnus, Skolithos), is indicative of the distal Skolithos ichnofacies, corresponding to the middle shoreface zone (Buatois and Mángano 2011).
The lower shoreface facies in the Djebel Ksar and Vieux Tenes outcrops show an inconsistency in the degree of bioturbation. The surfaces of some sandstone beds and some sandy marl interbeds are moderately to strongly bioturbated (BI: 3 to 6), while the interfaces of the sandstone beds are, in general, weakly bioturbated (BI: 0 to 1). The ichnodiversity is relatively high (eleven identified ichnogenera) compared to the upper to middle shoreface sandstones. The trace fossils are characterised by (1) the abundance of horizontal burrows such as Beaconites (Fig. 13a), Thalassinoides (two-dimensional networks; Fig. 18f), Taenidium (Fig. 16c, d), Planolites (Fig. 14a) and Ophiomorpha (horizontal components only; Fig. 15j); (2) the infrequent presence of vertical and inclined burrows such as Cylindrichnus (Fig. 13b-d), small-sized Arenicolites (Fig. 12b), Rosselia (Fig. 16a, b), Ophiomorpha that always occur in sandstone beds; (3) the dominance of traces generated by detritus-feeding organisms (Beaconites, Taenidium, Planolites, Thalassinoides); (4) a variety in the behaviour of organisms (feeding: Thalassinoides, Taenidium, Planolites, Zoophycos and dwelling: Ophiomorpha, Arenicolites). These combined features allow us to attribute this trace association to a relatively deeper and quieter environment of the Cruziana ichnofacies (Pemberton et al. 1992; MacEachern et al. 1999; Carmona et al. 2008; Frey and Dashtgard 2011). The vertical burrows mentioned above are related to opportunistic organisms that exploit nutrients during post-storm quiescent periods (Mángano et al. 2005; Buatois and Mángano 2011). Also, the presence of Zoophycos (Fig. 19e, f) in the last sandstone levels confirms a relatively deep environment and fair-weather conditions.
The highest sandstone levels (Allala River Sandstone Formation) at Ounsour Anhas, are also assigned to the lower shoreface. These deposits are heavely bioturbated (BI: 5 to 6), they yielded mainly three-dimensional Thalassinoides boxwork and rare Ophiomorpha burrows. These caracteristics suggest conditions of high oxygenation and abundance of nutrient matter (probably by fluvial supply).
In general, the offshore mudstone facies are characterised by pervasive burrowing (BI: 4–5) of high diversity. Distribution of ichnogenera is consistent with fully marine, unstressed settings (Pemberton et al. 2012; Dashtgard et al. 2012). In contrast to these data, the outcrops of the offshore dark, silty marls deposits of the Djebel Ksar, Vieux Tenes, Kaïsar, Ounsour Anhas and Djebel Ach Zerfet are weakly or unbioturbated. The scarcity or absence of trace fossils in the lower and upper offshore facies is probably related to oxygen-depleted conditions (Głuszek 1998; Mángano et al. 2005). The anaerobic conditions have been inferred by (1) the dark colour; (2) the presence of microbially induced carbonate concretions; (3) the occurrence of framboidal pyrite, clotted microfabric and the faint undulating lamination inside carbonate concretions; (4) pyritized and small foraminifera; (5) the preservation of plant debris (Nemra et al. 2019; Nemra 2020). These oxygen-depleted conditions were probably caused by sea-water column stratification.
Assessment of storm signature on sedimentation
Wave-dominated, generally non-tidal shorefaces may be weakly, moderately to intensely influenced by storms (MacEachern and Pemberton 1992; MacEachern and Bann 2008; Dashtgard et al. 2021). Based on this, three types of wave-dominated shorefaces have been distinguished: storm-affected, storm-influenced, and storm-dominated (Dashtgard et al. 2012). The storm-affected shoreface is characterised by intensely bioturbated muddy to silty sandstone beds of lower to middle shoreface related to fair-weather conditions. These deposits may be intercalated by thin storm beds that are commonly bioturbated with limited preservation of primary sedimentary structures (Buatois and Mángano 2011; Dashtgard et al. 2012). The storm-influenced shoreface is recognised by intensely bioturbated fair-weather muddy sandstone beds, alternating with unbioturbated to slightly bioturbated sandstone beds that reflect storm events. These latter might be bioturbated by vertical dwelling structures representing initial opportunistic colonisation (storm-related suite) and, at the top, by deposit-feeders, which in turn is followed by the re-establishment of fair-weather conditions (Mángano et al. 2005; Buatois and Mángano 2011; Dashtgard et al. 2012). The high energy conditions, causing repeated erosion in the storm-dominated shoreface, commonly preclude the preservation of biogenic structures (Mángano et al. 2005; Buatois and Mángano 2011). Only the remnant of deepest components of the storm-related suite might be preserved. In fact, the successions of the storm-dominated shoreface are represented by slightly to unbioturbated, low-angle undulatory and parallel-laminated sandstone beds (Buatois and Mángano 2011; Dashtgard et al. 2012). In the storm-dominated offshore, the bioturbation may be less and characterised by a small and dispersed vertical storm-related suite, most commonly Skolithos, Arenicolites, and Ophiomorpha (Mángano et al. 2005; Buatois and Mángano 2011).
In the Kaïsar, Ounsour Anhas and Djebel Ach Zerfet areas, the distinction between the lower and middle shoreface is difficult to determine due to the scarcity of sedimentary structures and the lack of trace fossils characteristic of fair-weather conditions. In these areas, the dominance of thick vertical dwelling burrows of suspension feeders, the rarity of traces and the low ichnodiversity in the shoreface facies confirm stressful conditions related the dominance of storm-wave activity. Furthermore, the offshore transition/upper offshore facies in Ounsour Anhas provide additional evidence of storm domination and high-energy conditions. The fining-upward sandstone intercalations with erosive or channelized basal surfaces within the sandy-silty marl are more frequent, thicker and host low-density Skolithos burrows (fig. 17d) and Thalassinoides shafts. At the top of the section, the presence of Zoophycos (Fig. 19a-d) in the uppermost sandstone intercalations is worth noting. The occurrence of these Zoophycos in offshore as well as in shoreface facies (Djebel Ksar) will be discussed later. The disseminated Skolithos burrows (elements of the Skolithos ichnofacies, storm–related Skolithos) are produced by post-depositional opportunistic organisms that exploit suspended nutrient matter in the water column after storm events (Pemberton et al. 1992; Mángano et al. 2005; Buatois and Mángano 2011).
At Vieux Tenes and Djebel Ksar, it is easier to distinguish the facies of the middle shoreface from those of the lower shoreface, according to the appearance of traces that indicate fair-weather conditions (horizontal burrows of the Cruziana ichnofacies). The trace-fossil association at these areas shows an alternation of fair-weather and agitated periods (storm-wave activity). The agitated periods are represented by non-bioturbed or weakly bioturbed sandstone beds with vertical dwelling burrows of suspension feeders (Cylindrichnus, Arenicolites, Rosselia, Ophiomorpha). In contrast, the facies related to fair-weather periods (sandstone and sandy marl) are characterised by a moderately to heavily bioturbated facies with horizontal burrows of detritus-feeding organisms (Beaconites, Taenidium, Planolites, Thalassinoides). This implies alternating agitated and calmer periods, thus, the shoreface in these localities was rather under the influence of storm-wave activity.
The lateral distance between the different outcrops is approximately one kilometre (Fig. 1b, c). In other words, the deposits of this same siliciclastic platform seem to be dependent on the localities that are either dominated or influenced by the storm-wave activity. This lateral variation in the intensity of storm waves might be controlled by a contrasting palaeogeomorphology of the coastline (narrow coastal reliefs and gravel beach adjacent to wider sandy bays). Therefore, the gravel beach is more exposed to wave activity, whearas the sandy bays are relatively less affected by wave action. This configuration, which results from differential erosion of the coast, can also be observed in present-day environments (Hemdane et al. 2016). Indeed, the facies at the base of Kaïsar (open framework conglomerates, sandstone, cemented conglomerates with Ostrea clasts, pebbly sandstones) have been assigned to a gravel-beach environment (beachface/shoreface complex) located at the base of the cliffs (Nemra 2020). In addition, the rapid lateral variations in thickness and facies have been documented (Nemra 2020); for example, at Vieux Tenes and Djebel Ksar, the conglomerates appear as a single bed (in each section) with a thickness of less than 1 m, whereas they occur as a set of decimetric to metric levels at Kaïsar.
Zoophycos of Tenes Miocene succession: palaeoenvironment and ethology
The different data regarding Zoophycos show that the palaeoenvironment shifted over geologic time (Bottjer et al. 1988; Olivero and Gaillard 1996). Indeed, the presence of Zoophycos in Palaeozoic successions has been reported from shallow-marine to deep-sea palaeoenvironments (Bottjer et al. 1988). Consequently, their occurence is not considered evidence of a palaeoenvironment deeper than shelf (Uchman et al. 2016). During the Jurassic, the Zoophycos-producing organism had migrated to the deep-sea environment (Olivero 2003) such as slopes, deep-sea fans and basin floors (Wetzel and Werner 1981; Uchman and Demircan 1999; Löwemark et al. 2006). Contrary to these observations, Zoophycos (Fig. 19) of the Tenes region (Middle Miocene) have been assigned to the Cruziana ichnofacies. They occur in sandstone facies of shallow-marine environments ranging from the lower shoreface (Zoophycos of Djebel Ksar area; Fig. 19e, f) to the upper offshore (Zoophycos of Ounsour Anhas area; Fig. 19a-d). Similarly, some Jurassic (Pemberton et al. 2001) and other Miocene (Pervesler and Uchman 2004) Zoophycos have also been reported in similar marine settings (lower shoreface to upper offshore). In conclusion, we support the idea of Pervesler and Uchman (2004) that though Mesozoic and Cenozoic Zoophycos have often been reported from deep-marine environments (slope edge to basin), it still occurs in the offshore to shoreface zone.
The studied Zoophycos (Fig. 19a-d, g, h) were observed as endichnia in the upper part of fining-upward sandstone beds, interpreted as the result of storm-wave action. Opportunistic organisms producing Zoophycos exploited the uppermost part of the nutrient-rich sandy-clay sediment. Zoophycos are often associated with low-oxygen environments (Ekdale 1992) and the large trace size (30-100 cm) is not necessarily related to the oxygenation of the environment (Wetzel and Werner 1981; Giannetti 2010), but rather to the search for and storage of nutrients, beneath the redox level, out of reach of other bioturbating organisms (Bromley, 1991). Their abundance and large size indicate that the sediment was nutrient-rich and the sedimentation rate was low, at least during the period of exploitation (Löwemark et al. 2006; Nasiri et al. 2018). The Zoophycos tracemaker colonised the surface that separates the Tenes Blue Marls Formation (Middle Miocene) from the overlying formation, represented by yellowish marls of Tortonian age (upper Miocene). This surface is regarded as a discontinuity surface (sedimentation breakup surface or omission surface sensu Ekdale and Lewis 1991) and may correspond to the maximum flooding surface of the first Miocene transgressive cycle sensu Perrodon (1957).
Zoophycos of the last sandstone beds of the Tenes Blue Marls were developed in the upper offshore zone (Nemra 2020) on a relatively narrow and steep sea floor. This is documented by (1) the occurrence of synsedimentary deformation structures like slumps and faults (Perrodon 1957; Nemra et al. 2019); (2) the co-existence of proximal material (plant debris and benthic foraminifera) and pelagic material (high abundance of planktic foraminifera), which can be explained by the narrow shelves common in tectonically active basins such as the Mediterranean basin (Nemra et al. 2019).
Conclusions
The trace fossils of the Middle Miocene outcrops in the Tenes area have been studied for the first time in five localities (Vieux Tenes, Kaïsar, Djebel Ksar, Djebel Ach Zerfet and Ounsour Anhas). Fifteen ichnogenera have been identified in the Allala River sandstone and Tenes Blue Marl formations, reflecting multiple responses of the benthic fauna to environmental factors and, thus, they provide valuable information on the depositional environments.
The trace-fossil assemblages show two ichnofacies: (1) Skolithos ichnofacies related to an upper/middle shoreface environment, and (2) Cruziana ichnofacies that reflects lower shoreface to transition offshore environment. Skolithos ichnofacies is characterised by low ichnodiversity and the presence of scattered vertical burrows (Skolithos, Diplocraterion, Arenicolites, Ophiomorpha) of suspension-feeding organisms, which suggests stressful conditions associated with frequent high-energy events (storms) and continuous sedimentation and remobilization of detrital deposits. Locally, Thalassinoides burrows are well developed (the lower part of Allala River sandstone at Kaïsar, Ounsour Anhas and Djebel Ksar) and show a higher proportion of vertical components, thick burrows and generally dense networks and boxworks (Ounsour Anhas and Kaïsar). All these characteristics suggest an oxygenated environment and high energy conditions. In some cases, distal expression of Skolithos ichnofacies might be distinguished by the occurrence of rare elements from the Cruziana ichnofacies (Rosselia and Taenidium) and allow us to assign the facies related to the middle shoreface zone.
The Cruziana ichnofacies is characterised by a relatively high bioturbation and dominance of horizontal burrows of deposit-feeding organisms, such as Thalassinoides (horizontal components), Taenidium, Planolites, Beaconites and Ophiomorpha (horizontal components). These trace fossils represent a wide variety in behaviours (feeding, locomotion, gardening, resting and dwelling). In this ichnofacies, rare vertical burrows might occur, such as Arenicolites (small size), Rosselia and Skolithos, which could be related to storm facies and suggest an opportunistic behaviour.
The comparison of trace fossils between the studied localities allows us to estimate different degrees of storm-wave influence. Kaïsar, Ounsour Anhas and Djebel Ach Zerfet outcrops are dominated by thick, deep, vertical dwelling burrows of suspension feeders and show low ichnodiversity that confirm storm-wave dominated conditions. In these localities the distinction between the zonations of shoreface is difficult because of the lack of trace fossils related to fair-weather conditions. On the other hand, at Vieux Tenes and Djebel Ksar localities, the alternation of fair-weather-related trace fossils (Thalassinoides networks, Taenidium, Beaconites, Planolites) and the storm-related trace fossils (Cylindrichnus, Arenicolites, Rosselia, Skolithos) in the lower shoreface zone indicate storm-wave influence. In these areas, the occurrence of fair-weather-related trace fossils allows as to easily identify the lower shoreface zone.
Contrary to what is globally documented in siliciclastic platforms, the offshore related facies in the Tenes area are weakly bioturbated to unbioturbated. This is interpreted as being the result of oxygen-depleted conditions, which suggests sea-water column stratification.
Finally, the Zoophycos recorded at Ounsour Anhas and Djebel Ksar outcrops are related to offshore transition and lower shoreface facies, respectively. Although Cenozoic Zoophycos are common in deep-marine deposits, these observations confirm that the occurrence of Cenozoic Zoophycos extends to the shoreface. In addition, the presence of these Zoophycos at the top of the storm-related sandstones beds attests to an opportunistic behaviour. The large size of these traces at Ounsour Anhas is probably related to oxygen-depleted conditions and low sedimentation rate or omission.
Data availability
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
References
Adserà, P. (2018). Distinctive morphological features Skolithos linearis from the Eocene of the Aínsa-Jaca Basin (South-Central Pyrenees). Batalleria, 26, 4-10.
Alpert, S. P. (1974). Systematic Review of the Genus Skolithos. Journal of Paleontology, 48, 661-669.
Bann, K. L., & Fielding, C. R. (2004). An integrated ichnological and sedimentological comparison of non-deltaic shoreface and subaqueous delta deposits in Permian reservoir units of Australia, In D. McIlroy (Ed.), The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis. Geological Society London Special Publication, 228, 273-310.
Belaústegui, Z., & de Gibert, J. M. (2013). Bow-shaped, concentrically laminated polychaete burrows: A Cylindrichnus concentricus ichnofabric from the Miocene of Tarragona, NE Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 381-382, 119-127.
Belkebir, L., Bessedik, M., Ameur-Chehbeur, A., & Anglada, R. (1996). Le Miocène des bassins nord-occidentaux d’Algérie: biostratigraphie et eustatisme. (pp. 553-561). In Géologie de l’Afrique et de l’Atlantique Sud: Actes Colloques Angers 1994. Pau: Edition Elf Aquitaine 16.
Bendella M. (2012). Evolution des traces fossiles dans l’espace (Tell, Atlas, Sahara) et dans le temps en Algérie occidentale: inventaire et mise en évidence des évènements anoxiques (pp. 1-161). PhD thesis. University Oran, Algérie.
Bendella, M., & Ouali Mehadji, A. (2014). Depositional environment and ichnology (Nereites ichnofacies) of the Late Devonian Sahara region (SW Algeria). Arabian Journal of Geosciences, 8, 5303-5316.
Bendella, M., Benyoucef, M., Cherif, A., & Benhamou, M. (2011). Ichnology and sedimentology of the Argiles de Saïda formation (Callovo-Oxfordian) of the Djebel Brame (Tiaret, Algeria). Bulletin de la Société géologique de France, 5, 417-425.
Bessedik, M., Belkebir, L., & Mansour, B. (2002). Le Miocène supérieur du bassin du Bas Chélif: attribution biostratigraphique à partir des foraminifères planctoniques. Mémoire du Service Géologique de l’Algérie, 11, 187-194.
Bluck, B. J. (1967). Sedimentation of beach gravels: examples from South Wales. Journal of Sedimentary Petrology, 37, 128-156.
Bluck, B. J. (1999). Clast assembling, bed-forms and structure in gravel beaches. Transactions of the Royal Society of Edinburgh: Earth Sciences, 89, 291–332.
Bottjer, D. J., Droser, M. L., & Jablonski, D. (1988). Palaeoenvironmental trends in the history of trace fossils. Nature, 333 (6170), 252–255.
Boyd, C., & McIlroy, D. (2017). Three-dimensional morphology of Beaconites capronus from Northeast. England. Ichnos, 24(4), 250–258.
Bradshaw, M. A. (2010). Devonian trace fossils of the Horlick Formation, Ohio Range, Antarctica: systematic description and palaeoenvironmental interpretation. Ichnos, 17, 58-114.
Brives, A. (1913). Carte géologique au 1/50 000 de Ténès - Cap Tenes. Service Géolgique de l’Algérie.
Bromley, R. G. (1991). Zoophycos: strip mine, refuse dump, cache or sewage farm? Lethaia, 24, 460-462.
Bromley, R. G., & Ekdale, A. A. (1998). Ophiomorpha irregulaire (trace fossil): redescription from the Cretaceous of the Book Cliffs and Wasatch Plateau, Utah. Journal of Paleontology, 72, 773-778
Bromley, R. G., & Frey, R. W. (1974). Redescription of the trace fossil Gyrolithes and taxonomic evaluation of Thalassinoides, Ophiomorpha and Spongeliomorpha. Bulletin of the Geological Society of Denmark, 23, 311-335.
Bromley, R. G., & Hanken, N. M. (2003). Structure and function of large, lobed Zoophycos, Pliocene of Rhodes, Greece. Palaeogeography, Palaeoclimatology, Palaeoecology, 192, 79-100.
Bromley, R. G., Ekdale, A. A., & Richter, B. (1999). New Taenidium (trace fossil) in the Upper Cretaceous chalk of northwestern Europe. Bulletin of the Geological Society of Denmark, 46, 47-51.
Buatois, L. A., & Mángano, M. G. (1993). Trace fossils from a Carboniferous turbidic lake: implications for the recognition of an additional nonmarine ichnofacies. Ichnos, 2, 217-224.
Buatois, L. A., & Mángano, M. G. (2011). Ichnology: Organism-Substrate Interactions in Space and Time (pp. 1-358). Cambridge: Cambridge University Press,
Buatois, L. A., Mángano, M. G., & Sylvester, Z. (2001). A diverse deep-marine Ichnofauna from the Eocene Tarcau sandstone of the Eastern Carpathians, Romania. Ichnos, 8, 23-62.
Carmona, N. B., Buatois, L. A., Mángano, M. G., & Bromley, R. G. (2008). Ichnology of the Lower Miocene Chenque Formation, Patagonia, Argentina: animal-substrate interactions and the Modern Evolutionary Fauna. Ameghiniana, 45, 93-122.
Cassan, G. (1968). Foraminifères planctoniques du stratotype du Cartennien (Tenes) Algérie. Committee Mediterranean Neogene Stratigraphy. Proc. IV Session, Bologne, 1967, Giornale di geologia 2, XXXV, fasc. II, 377-386.
Catuneanu, O., & Zecchin, M. (2013). High-resolution sequence stratigraphy of clastic shelves II: controls on sequence development. Marine and Petroleum Geology, 39, 26-38
Chamberlain, C. K. (1971). Morphology and ethology of trace fossils from the Ouachita Mountains, southeast Oklahoma. Journal of Paleontology, 45, 212-246.
Clifton, H. E., & Thompson, J. K. (1978). Macaronichnus segregatis: a feeding structure of shallow marine polychaetes. Journal of Sedimentary Petrology, 48(4), 1293-1302
Cornish, F. G. (1986). The Trace-Fossil Diplocraterion: Evidence of Animal-Sediment Interactions in Cambrian Tidal Deposits. Palaios, 1, 478-491.
Crimes, T. P. (1992). Changes in the trace fossil biota across the Proterozoic-Phanerozoic boundary. Journal of the Geological Society, 149, 637-646.
Crimes, T. P., Goldring, R., Homewood, P., Stuijvenberg, J. V., & Winkler, W. (1981). Trace fossil assemblages of deep-sea fan deposits, Gurnigel and Schlieren flysch (Cretaceous-Eocene), Switzerland. Eclogae Geologicae Helvetiae, 74, 953-995.
Dam, G. (1990). Taxonomy of trace fossils from the shallow marine Lower Jurassic Neill Klinter Formation, East Greenland. Bulletin of the Geological Society of Denmark, 38, 119-144.
Dashtgard, S. E., & Gingras, M. K. (2012). Marine invertebrate neoichnology. In D. Knaust, & R. G. Bromley (Eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology 64, 273-295.
Dashtgard, S. E., Maceachern, J. A., Frey, S. E., & Gingras, M. K. (2012). Tidal effects on the shoreface: Towards a conceptual framework. Sedimentary Geology, 279, 4-61.
Dashtgard, S. E., Vaucher, R., Yang, B., & Dalrymple, R.W. (2021) Hutchison medallist 1. Wave-dominated to tide-dominated coastal systems: a unifying model for tidal shorefaces and re-finement of the coastal-environments classification scheme. Geoscience Canada, 48, 5– 22.
Delteil, J. (1974). Tectonique de la chaîne alpine en Algérie d’après l’étude du Tell oriental (Mont de la Mina, Beni Chougrane, Dahra) (pp. 1-294). PhD thesis. University of Nice, France.
Ekdale, A. A. (1992). Muckraking and mudslinging: The joys of deposit-feeding. Short Courses in Paleontology, 5, 145–171.
Ekdale, A. A., & Bromley, R. G. (2012). Eolian environments. In D. Knaust, & R. G. Bromley (Eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology 64, 419-437.
Ekdale, A. A., & Lewis, D. W. (1991). The New Zealand Zoophycos revisited: Morphology, ethology, and paleoecology. Ichnos, 1, 183-194.
Ekdale, A. A., Bromley, R. G., & Pemberton, S. G. (1984). Ichnology: Trace fossils in sedimentology and stratigraphy. Society of Economic Paleontologists and Mineralogists 15, 1-317, Tulsa, Oklahoma.
Ekdale, A. A., Bromley, R. G., & Loope, D. B. (2007). Ichnofacies of an ancient erg: A climatically influenced trace fossil association in the Jurassic Navajo Sandstone, Southern Utah, USA. In W. Miller III (Ed.), Trace fossils: Concepts, problems, prospects (pp. 562-564), Amsterdam: Elsevier.
El-Sabbagh, A., El-Hedeny, M., & Al Farraj, S. (2017). Thalassinoides in the middle Miocene succession at Siwa oasis, northwestern Egypt. Proceedings of the Geologists' Association, 128, 222-233.
Fenêt, B. (1975). Recherches sur l’alpinisation de la bordure septentrionale du Bouclier africain à partir de l’étude d’un élément de l’orogène nord-maghrébin: les monts du Djebel Tessala et les massifs du littoral de l’Oranais (pp. 1-301). PhD thesis. University of Nice, France.
Fillion, D., & Pickerill, R. K. (1984). Systematic ichnology of the Middle Ordovician Trenton Group, St. Lawrence lowland, eastern Canada. Maritime Sediments and Atlantic Geology, 20, 1-41.
Fleming, C. A. (1973). "Fossil cuff-links": a new Miocene tract: fossil of the genus Diplocraterion from New Zealand. Tohoku University Science Reports, Special Volume 6 (Hatai Memorial Volume), 415-418.
Frey, R. W., & Bromley. R. G. (1985). Ichnology of American chalks: the Selma Group (Upper Cretaceous), western Alabama. Canadian Journal of Earth Sciences, 22, 801-828.
Frey, R. W., Curran, H. A., & Pemberton, S. G. (1984). Tracemaking activities of crabs and their environmental significance: the ichnogenus Psilonichnus. Journal of Paleontology, 58, 333-350.
Frey, S. E., & Dashtgard, S. E. (2011). Sedimentology, ichnology and hydrodynamics of strait-margin, sand and gravel beaches and shorefaces: Juan de Fuca Strait, British Columbia, Canada. Sedimentology, 58(6), 1326-1346.
Frey, R. W., & Howard, J. D. (1985). Trace fossils from the Panther Member, Star Point Formation (Upper Cretaceous), Coal Creek Canyon, Utah. Journal of Paleontology, 59, 370-404.
Frey, R. W., & Howard, J. D. (1990). Trace fossils and depositional sequences in a clastic shelf setting, Upper Cretaceous of Utah. Journal of Paleontology, 64, 803-820.
Frey, R. W., Howard, J. D., & Pryor, W. A. (1978). Ophiomorpha: its morphologic, taxonomic, and environmental significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 23, 199-229.
Fürsich, F. T. (1974). On Diplocraterion Torell 1870 and the significance of morphological features in vertical, spreiten-bearing, U-shaped trace fossils. Journal of Paleontology, 48, 952-962.
Fürsich, F. T. (1975). Trace fossils as environmental indicators in the Corallian of England and Normandy. Lethaia, 8, 151-172.
Fürsich, F.T., Wilmsen, M., & Seyed-Emami, K. (2006). Ichnology of Lower Jurassic beach deposits in the Shemshak Formation at Shahmirzad, southeastern Alborz Mountains, Iran. Facies, 52, 599-610.
Gaillard, C., Hennebert, M., & Olivero, D. (1999). Lower Carboniferous Zoophycos from the Tournai area (Belgium): environmental and ethologic significance. Geobios, 32, 513-524.
Gani, M. R., Bhattacharya, J. P., & Maceachern, J. A. (2007). Using ichnology to determine relative influence of waves, storms, tides, and rivers in deltaic deposits: examples from Cretaceous Western Interior Seaway, U.S.A. In J. A MacEachern, K. L Bann, M. K Gingras, et al (Eds.), Applied ichnology. Society of Economic Paleontologists and Mineralogists, 52, 209-225.
Gerard, J. R. F., & Bromley, R. G. (2008). Ichnofabrics in Clastic Sediments: Applications to Sedimentological Core Studies (pp. 1-97). Gerard J. R. F. (Ed.).
Giannetti, A. (2010). Influence of climate, sea-level changes and tectonics on ichnoassemblages distribution in a carbonate dominated, deep-marine environment (Upper Paleocene, Zumaya section). Palaeogeography, Palaeoclimatology, Palaeoecology, 285, 104-118.
Gingras M. K., & MacEachern J. A. (2012). Tidal ichnology of shallow-water clastic settings, In R.A Davis Jr., & R. W. Dalrymple (Eds.), Principles of Tidal Sedimentology (pp. 57-77). Dordrecht, Heidelberg: Springer.
Gingras, M. K., Pemberton, S. G., Saunders, T. D. A., & Clifton, H. E. (1999). The ichnology of modern and Pleistocene brackish-water deposits at Willapa Bay, Washington: variability in estuarine settings. Palaios, 14, 352-374.
Glangeaud, L. (1932). Etude géologique de la région littorale de la province d'Alger (pp. 1-617). Bulletin du Service de la Carte géolgique de l’Algérie.
Głuszek, A. (1998). Trace fossils from Late Carboniferous storm deposits, Upper Silesia Coal Basin, Poland. Acta Palaeontologica Polonica, 43, 517-546.
Goldring, R. (1964). Trace fossils and the sedimentary surface in shallow water marine sediments. Developments in Sedimentology, 1, 136-143.
Goldring, R. (1996). The sedimentological significance of concentrically laminated burrows from Lower Cretaceous Ca-bentonites, Oxfordshire. Journal of the Geological Society, 153, 255–263.
Hammersburg, S. R., Hasiotis, S. T., & Robison, R. A. (2018). Ichnotaxonomy of the Cambrian Spence Shale Member of the Langston Formation, Wellsville Mountains, Northern Utah, USA. Paleonotological Contributions, 20, 1-66.
Hofmann, R., Mángano, M. G., Elicki, O., & Shinaq, R. (2012). Paleoecologic and biostratigraphic significance of trace fossils from shallow-marginal-marine environment from the middle Cambrian (Stage 5) of Jordan. Journal of Paleontology, 86, 831- 955.
Howard, J. D., & Frey, R. W. (1984). Characteristic trace fossils in nearshore to offshore sequences, Upper Cretaceous of east-central Utah. Canadian Journal of Earth Sciences, 21, 200-219.
Keighley, D. G., & Pickerill, R. K. (1995). The ichnotaxa Palaeophycus and Planolites: historical perspectives and recommendations. Ichnos, 3, 301-309.
Knaust, D. (2004). Cambro-Ordovician trace fossils from the SW-Norwegian Caledonides. Geological Journal, 39, 1-24.
Knaust, D. (2015). Siphonichnidae (new ichnofamily) attributed to the burrowing activity of bivalves: ichnotaxonomy, behaviour and palaeoenvironmental implications. Earth-Science Reviews, 150, 497-519.
Knaust, D. (2017). Atlas of trace fossils in well core: Appearance, taxonomy and interpretation (pp. 1-209). Cham: Springer, Switzerland.
Knaust, D. (2018a). Teichichnus zigzag Frey and Bromley, 1985: a probable echiuran or holothurian burrow from the Jurassic offshore Norway. Paläontologische Zeitschrift, 92, 617-632. https://doi.org/https://doi.org/10.1007/s12542-018-0413-9.
Knaust, D. (2018b). The ichnogenus Teichichnus Seilacher, 1955. Earth-Science Reviews, 177, 386-403. https://doi.org/https://doi.org/10.1016/j.earscirev.2017.11.023.
Knaust, D. (2021). Rosselichnidae ifam. nov.: burrows with concentric, spiral or eccentric lamination. Papers in Palaeontology, 7, 1847-1875.
Knaust, D., Thomas, R. D. K., & Curvan, H. A. (2018). Skolithos linearis Haldeman, 1840 at its early Cambrian type locality, Chickies Rock, Pennsylvania: Analysis and designation of a neotype. Earth-Science Reviews, 185, 15-3.
Lepvrier, C., & Magné, J. (1975). Le Néogène «postnappes» du Tell septentrional à l’Ouest d’Alger. Bulletin du la Société Géologique de France, 7, 612-619.
Locklair, R. E., & Savrda, C. E. (1998). Ichnology of rhythmically bedded Demopolis Chalk (Upper Cretaceous, Alabama): Implications for paleoenvironment, depositional cycle origins, and tracemaker behavior. Palaios, 13, 423-438.
Löwemark, L., Lin, H-L., & Sarnthein, M. (2006). Temporal variations of the trace fossil Zoophycos in a 425 ka long sediment record from the South China Sea: Implications for the ethology of the Zoophycos-producer. Geological Magazine, 143, 105-114.
Lucas, S. G., & Lerner, A. J. (2006). Invertebrate ichnofossil assemblages of the Upper Triassic Redonda Formation at Mesa Redonda, east-central New Mexico. New Mexico Museum of Natural History and Science Bulletin 37, 122-127.
MacDonald, D. I. M. (1982). Palaeontology and ichnology of the Cumberland Bay Formation, South Georgia. British Antarctic Survey Bulletin, 57, 1-14.
MacEachern, J. A., & Bann, K. L. (2008). The role of ichnology in refining shallow marine facies models. In G. J. Hampson, R. J. Steel, P. B. Burgess, & R. W. Dalrymple (Eds.), Recent advances in models of siliciclastic shallow-marine stratigraphy. Tulsa, USA: Society for Sedimentary Geology (SEPM) 90, 73-116.
MacEachern, J. A., & Gingras, M. K. (2007). Recognition of brackish-water trace fossil suites in the Cretaceous Western Interior Seaway of Alberta, Canada. In R.G. Bromley, L. A. Buatois, M. G. Mángano, J. F. Genise, & R. N. Melchor (Eds.), Sediment-Organism Interactions: a Multifaceted Ichnology. SEPM Special Publication, 89, 149-194.
MacEachern, J. A., & Pemberton, S. G. (1992). Ichnological aspects of Cretaceous shoreface successions and shoreface variability in the Western Interior Seaway of North America. In S. G. Pemberton (Ed.), Applications of Ichnology to Petroleum Exploration. SEPM, Core Workshop, 17,. 57-84.
MacEachern, J. A., Zaitlin, B. A., & Pemberton, S. G. (1999). A sharp-based sandstone of the Viking Formation, Joffre Field, Alberta, Canada, criteria for recognition of transgressively incised shoreface complexes. Journal of Sedimentary Research, 69, 876-892.
Mángano, M. G., & Buatois, L. A. (2004). Ichnology of Carboniferous tide-influenced environments and tidal flat variability in the North American Midcontinent. In D. McIlroy (Ed.), The application of ichnology to palaeoenvironmental and stratigraphic analysis. Geological Society Special Publication, 228, 157-178.
Mángano, M. G., Buatois, L. A., & Muñiz Guinea, F. (2005). Ichnology of the Alfarcito Member (Santa Rosita Formation) of northwest Argentina: animalsubstrate interactions in a lower Paleozoic wave-dominated shallow sea. Ameghiniana, 42, 641-668.
Martinsson, A. (1970). Taphonomy of trace fossils. In T. P. Crimes, & J. C. Harper (Eds.), Trace fossils (pp. 323-330). Liverpool: Seel House Press.
Massari, F., & Parea, G. C. (1988). Progradation of gravel beach sequences in a moderate- to high-energy, microtidal marine environment. Sedimentology, 35, 881-913.
McCann, T., & Pickerill, R. K. (1988). Flysch trace fossils from the Cretaceous Kodiak Formation of Alaska. Journal of Paleontology, 62, 330-348.
McIlroy, D. (2007). Lateral variability in shallow marine ichnofabrics; implications for the ichnofabric analysis method. Journal of the Geological Society of London, 164, 359-369.
Nagy, J., Tovar, F. J. R., & Reolid, M. (2016). Environmental significance of Ophiomorpha in a transgressive-regressive sequence of the Spitsbergen Paleocene. Polar Research, 35, 24192.
Nasiri, Y., Reza, M.-H., Mahboubi, A., Olivero, D., & Mosaddegh, H. (2018). Zoophycos ichnogenus distribution and paleoenvironmental analysis: examples from the Mississippian Mobarak Formation (Alborz Basin, Iran). Historical Biology, 32, 848-867.
Nemra, A. (2020). Lithostratigraphie, environnements sédimentaires et aspect génétique des concrétions calcaires des terrains miocènes de la région de Ténès (bordure nord orientale du bassin du Bas Chélif, Algérie nord-ouest) (pp. 1-223). PhD thesis. University of Oran 2, Algeria.
Nemra, A., Ouali Mehadji, A., Munnecke, A., Belkhedim, S., & Belkbir, L. (2019). Carbonate concretions in Miocene mudrocks in NW Algeria: Types, geochemistry, and origins. Facies, 65 (17).
Neurdin-Trescartes, J. (1992). Le remplissage sédimentaire du basin néogène du Chélif, modèle de référence de basins intramontagneux. PhD thesis. Université Pau et pays de l’Adour, France.
Olivero, D. (2003). Early Jurassic to Late Cretaceous evolution of Zoophycos in the French Subalpine Basin (southeastern France). Palaeogeography, Palaeoclimatology, Palaeoecology, 192, 59-78.
Olivero, D., & Gaillard, C. (1996). Palaeoecology of Jurassic Zoophycos from south-eastern France. Ichnos, 4, 249-260.
Olivero, D., & Gaillard, C. (2007). A constructional model for Zoophycos. In W. Miller III Elsevier (Ed.), Trace fossils: concepts, problems, prospects (pp. 466-477). Amsterdam: Elsevier.
Pemberton, S. G., & Frey, R. W. (1982). Trace fossil nomenclature and the Planolites Palaeophycus dilemma. Journal of Paleontology, 56, 843-881.
Pemberton, S. G., & Frey, R. W. (1984). Ichnology of storm-influenced shallow marine sequence: Cardium Formation (Upper Cretaceous) at Seebe, Alberta. In D. F. Scott, & D. J. Glass (Eds.), The Mesozoic of Middle North America. Canadian Society of Petroleum Geologists, Memoir 9, 281-304.
Pemberton, S. G., Van Wagoner, J. C., & Wach, G. D. (1992). Ichnofacies of a wave dominated shoreline. In S. G. Pemberton (Ed.), Application of ichnology to petroleum exploration. Society of Economic Paleontologists and Mineralogists Core Workshop, 17, 339-382.
Pemberton, S. G., Spila, M. V., Pulham, A. J., Saunders, T., Maceachern, J. A., Robbins, D., & Sinclair, I. (2001). Ichnology and sedimentology of shallow and marginal marine systems: Ben Nevis and Avalon Reservoirs, Jeanne D’Arc Basin. Geological Association of Canada, 15, 1-353.
Pemberton, S. G., MacEachern, J. A., Dashtgard, S. E., Bann, K. L., Gingras, M. K., & Zonneveld, J. P. (2012). Shorefaces. In D. Knaust & R. G. Bromley (Eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology, 64, 563-604.
Perrodon, A. (1957). Etude géologique des basins néogènes sublittoraux de l’Algérie occidentale. Bulletin du Service de la Carte géolgique de l’Algérie, 12, 1-328.
Pervesler, P., & Uchman, A. (2004). Ichnofossils from the type area of the Grund formation (Miocene, Lower Badenian) in Northern Lower Austria (Molasse Basin). Geologica Carpathica, 55, 103-110.
Pieńkowski, G. (1985). Early Liassic trace fossil assemblages from the Holy Cross Mountains, Poland: Their distribution and marginal marine environments. In H. A Curran (Ed.), Biogenic structures: Their use in interpreting depositional Environments. Society of Economic Paleontologists and Mineralogists, Special Publication, 35, 37-51.
Pollard, J. E., Goldring, R., & Buck, S. G. (1993). Ichnofabrics containing Ophiomorpha: significance in shallow-water facies interpretation. Journal of the Geological Society of London, 150, 149-164.
Rajkonwar, C., Ralte, V. Z., Lianthangpuii, P. C., Tiwari, R. P., & Patel, S. J. (2014). Miocene ichnofossils from upper Bhuban Succession, Bhuban Formation (Surma Group), Mizoram, India. Journal of the Palaeontological Society of India, Special Publication, 5, 247-255.
Reading, H. G., & Collinson, J. D. (1996). Clastic coasts. In H. G. Reading (Ed.), Sedimentary Environments: Processes, Facies and Stratigraphy (pp. 154–231). Oxford: Blackwell Science.
Reineck, H.-E. (1963). Sedimentgefüge im Bereich der sudlichen Nordsee. Abhandlungen der Senckenbergische Naturforschende Gesellschaft, 505, 1-138.
Riahi, S., Uchman, A., Stow, D., Soussi, M., & Lattrache, K. B. I. (2014). Deep-sea trace fossils of the Oligocene–Miocene Numidian Formation, northern Tunisia. Palaeogeography, Palaeoclimatology, Palaeoecology, 414, 155-177.
Sanders, D. (2000). Rocky shore-gravelly beach transition, and storm/post-storm changes of a Holocene gravelly beach (Kos Island, Aegean Sea): stratigraphic significance. Facies, 42, 227-244.
Sarkar, S., Ghosh, S. K., & Chakraborty, C. (2009). Ichnology of a Late Palaeozoic ice-marginal shallow marine succession: Talchir Formation, Satpura Gondwana basin, central India. Palaeogeography, Palaeoclimatology, Palaeoecology, 283, 28-45.
Schlirf, M., & Uchman, A. (2005). Revision of the ichnogenus Sabellarifex Richter, 1921 and its relationship to Skolithos Haldeman, 1840 and Polykladichnus Fürsich, 1981. Journal of Systematic Palaeontology, 3, 115-131.
Scott, J. J., Buatois, L. A., & Mángano, M. G. (2012). Lacustrine environments. In D. Knaust, & R. G. Bromley (Eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology 64, 379-419.
Seike, K., Yanagishima, S., Nara, M., & Sasaki, T. (2011). Large Macaronichnus in modern shoreface sediments: Identification of the producer, the mode of formation, and paleoenvironmental implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 311, 224-229.
SN REPAL (1952) Le basin néogène du Chélif. Publication du XIXe Congrès géologique international d’Alger, Alger 16 (1èresérie), 1-56.
Taylor, A. M., & Goldring, R. (1993). Description and analysis of bioturbation and ichnofabric. Journal of the Geological Society, 150(1), 141–148.
Tiwari, R. P., Rajkonwar, C., Lalchawimawii, Malsawma, P. L. J., Ralte, V. Z., & Patel, S. J. (2011). Trace fossils from Bhuban Formation, Surma Group (Lower to Middle Miocene) of Mizoram India and their palaeoenvironmental significance. Journal of Earth System Science, 120, 1127-1143.
Tonkin, N. S. (2012) Deltas. In D. Knaust, R. G. Bromley (Eds.) Trace fossils as indicators of sedimentary environments, 64, 507 – 528.
Uchman, A. (1995). Taxonomy and palaeoecology of flysch trace fossils: the Marnosoarenacea Formation and associated facies (Miocene, Northern Apennines, Italy). Beringeria, 15, 3-115.
Uchman, A. (1998). Taxonomy and ethology of flysch trace fossils: revision of the Marian Ksiazkiewicz collection and studies of complementary material. Annales Societatis Geologorum Poloniae, 68, 105-218.
Uchman, A., & Demircan, H. (1999). A Zoophycos group trace fossil from Miocene flysch in southern Turkey: evidence for a U-shaped causative burrow. Ichnos, 6, 251-259.
Uchman, A., & Krenmayr, H. G. (1995). Trace fossils from the lower Miocene (Ottnangian) molasse deposits of Upper Austria. Paläontologische Zeitschrift, 69, 503-524.
Uchman, A., & Wetzel, A. (2012). Deep-sea fans. In R. G. Bromley, & D. Knaust, (Eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology, 64, 643-671.
Uchman, A., Pika-Biolzi, M., & Hochuli, P. A. (2004). Oligocene trace fossils from temporary flood plain ponds: an example from the freshwater molasses of Switzerland. Eclogae Geologicae Helvetiae, 97, 133-148.
Uchman, A., Hanken, N-M., Nielson, J. K., Grundvag, S-A., & Piasecki, S. (2016). Depositional environment, ichnological features and oxygenation of Permian to earliest Triassic marine sediments in central Spitsbergen, Svalbard, Polar Research, 35, 24782.
Vail, P. R. (1987). Seismic stratigraphy interpretation procedure. In A. W. Bally (Ed.) Atlas of Seismic Stratigraphy. American Association of Petroleum Geologists Studies in Geology 27, 1–10.
Vinn, O., & Toom, U. (2018). First description of rare Teichichnus burrows from carbonate rocks of the Lower Paleozoic of Estonia. Carnets de Géologie, 18, 305-312.
Virtasalo, J. J., Kotilainen, A. T., & Gingras, M. K. (2006). Trace fossils as indicators of environmental change in Holocene sediments of the Archipelago Sea, northern Baltic Sea. Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 453-467.
Wetzel, A. (2008). Recent bioturbation in the deep South China Sea: a uniformitarian ichnologic approach. Palaios, 23, 601-615.
Wetzel, A., & Werner, F. (1981). Morphology and ecological signifcance of Zoophycos in deep-sea sediments of NW Africa. Palaeogeography, Palaeoclimatology, Palaeoecology, 32, 185-212.
Zecchin, M. (2007). The architectural variability of small-scale cycles in shelf and ramp clastic systems: the controlling factors. Earth-Science Reviews, 84, 21-55.
Zhang, L.-J., Buatois, L. A., Gabriela Mángano, M., Qi, Y.-A., Zhang, X., Sun, S., & Tai, C. (2017). Early Triassic estuarine depauperate Cruziana Ichnofacies from the Sichuan area of South China and its implications for the biotic recovery in brackish-water settings after the end-Permian mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 485, 351-360.
Zhang, L., Fan, R., & Gong, Y. (2015). Zoophycos macro-evolution since 541 Ma. Scientific Reports, 5, 14954.
Acknowledgements
Our grateful thanks go to the Direction Générale de la Recherche Scientifique et du Développement Technologique (DGRSDT) for funding this work. The authors wish to thank our colleagues for their help during the field work. We are also grateful to Sören Jensen (University of Extremadura, Spain) and an anonymous reviewer for their very constructive comments that helped us to improve the manuscript. We are also grateful to the Editor-in-Chief Dieter Uhl for his constructive remarks. Alan Lord (Senckenberg Research Institute and Natural History Museum Frankfurt) is very much acknowledged for improving the English.
Funding
Salim Belkhedim acknowledges financial support from the Deutscher Akademischer Austauschdienst (DAAD, funding n° 57588362).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nemra, A., Tellal, J., Knaust, D. et al. Middle Miocene trace fossils from the Tenes area (NW Algeria) and their palaeoenvironmental implications. Palaeobio Palaeoenv 104, 327–362 (2024). https://doi.org/10.1007/s12549-023-00594-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-023-00594-y