
Abstract
Over the last years, the knowledge of the children’s diet is a topic of growing interest in dental anthropology. Our aim seeks to establish patterns of interpopulation and intrapopulation variability in dietary microwear among children from four Iberian sites dated to the Neolithic through Bronze Age. Buccal and occlusal surfaces are compared to assess whether their differential rates of microwear turnover correspond with dietary differences linked to social and biological maturation (e.g., weaning and shifts to adult-like diets). This study is based on the analysis of 46 deciduous molars (Udm1, Udm2, and Ldm2). Occlusal and buccal surfaces were observed using an environmental scanning electron microscope (ESEM) following standard microwear methodologies. The results show that from an interpopulation perspective, there are differences in the number of buccal scratches between Valdavara and the other sites. From an intrapopulation perspective, there was a greater number of buccal striations in the older age category from Cova de la Guineu and more occlusal pitting in the older age category from Cova dels Galls Carboners. This study shows the utility of the combined approach to buccal and occlusal microwear analysis as a means of understanding child dietary maturation in prehistory, showing that feeding practices and/or food choice can explain differences between specific age categories of children in addition to differences between archeological sites.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The origins of food production and its intensification contributed to widespread cultural, social, and economic transformations in prehistory which also had extensive implications for human biology (Pinhasi and Stock 2011). The development of food production in the Neolithic introduced new strategies to prepare plant and animal foods, such as cereal gruels and dairy products that reduced the duration of breastfeeding and permitted early weaning (Buikstra et al. 1986; Armelagos et al. 1991; Bocquet-Appel 2002; Howcroft 2013). The nutritional practices and stress experienced during childhood are reflected in later life health and influence demographic patterns, thus making children a sensitive indicator of overall population health (Dhavale et al. 2017; King et al. 2018). The transition between breastfeeding and weaning, when breast milk is gradually replaced by other kinds of foods, is a particularly significant cultural process with important biological ramifications (Mays 2016; Ventresca Miller et al. 2017). This transitional period is critical for child health status (Waterman et al. 2014; Fernández-Crespo et al. 2018; King et al. 2018), because a decrease in, or cessation of, breastfeeding can lower the flow of antibodies from mother to child which can increase the likelihood of contracting infectious diseases (Pearson et al. 2010; Meehan and Roulette 2013).
Child feeding practices and the timing of the weaning process depend on culturally mediated factors such as maternal investment and childcare practices, as well as environmental factors like seasonality (Pearson et al. 2010; Howcroft 2013; Scott and Halcrow 2017). Weaning practices reported in ethnographic studies of agricultural, pastoral, and forager-horticulturalist populations document the inclusion of soft complementary foods during the weaning process that include cereal gruels, porridges, milk, meat, soups, and blood (Sellen and Smay 2001; Pearson et al. 2010; Howcroft 2013; Han et al. 2016; Ventresca Miller et al. 2017; Scharlotta et al. 2018). Another practice involves the reduction of food hardness, toughness, or consistency through pre-mastication—a potentially beneficial cultural practice that may promote the enzymatic process of digestion with adult saliva (Pelto et al. 2010; Waters-Rist et al. 2011; Scott and Halcrow 2017).
Dental microwear is a powerful and non-destructive tool for dietary reconstruction that provides insights into the physical properties, consistency, and degree of cultural processing (e.g., cooking, grinding, drying, and exposure to exogenous abrasives) that foodstuffs undergo prior to consumption (Teaford and Lytle 1996; Lalueza et al. 1996; Polo-Cerdá et al. 2007; El-Zaatari 2010; Sołtysiak 2011; Romero et al. 2013; Schmidt et al. 2015; Mahoney et al. 2016; Estalrrich et al. 2017; Scott and Halcrow 2017; Schmidt et al. 2019). Variation in dental microwear has proven useful for distinguishing between human groups with ethnographically known or archeologically inferred socioeconomic and dietary strategies (Pérez-Pérez et al. 1994, 2003; Lalueza et al. 1996; Romero et al. 2004, 2013; Polo-Cerdá et al. 2007; El-Zaatari 2010; Schmidt et al. 2015; Estalrrich et al. 2017; Schmidt et al. 2019). While most microwear analyses have focused on the reconstruction of adult diets, the ability to distinguish human groups based on the properties of the foods consumed is also relevant for understanding dietary maturation from childhood to adulthood corresponding to practices like infant weaning and other social conventions (Halcrow and Tayles 2008; Knudson and Stojanowski 2008; Mahoney et al. 2016; Mays 2016; Scott and Halcrow 2017; Ventresca Miller et al. 2017; Mays et al. 2017).
Dietary microwear accumulates on both the buccal and occlusal surfaces, but with important distinctions related to chewing cycle. During the mastication, two phases of movement occur: phase I, puncture crushing, in which the upper and lower molars compress the food between opposing occlusal surfaces, and phase II or chewing cycle, in which the molars separate and no pressure is exerted on the bolus (Kay and Hiiemae 1974; Tausch et al. 2015). For that reason, dietary microwear requires different modes of analysis and interpretation for each surface. Dental microwear features on the buccal surface appear as scratches of various lengths, frequency, and orientation corresponding to the properties of the food masticated and the endogenous (e.g., phytoliths) and exogenous abrasives (e.g., grit and dust) it contains (Pérez-Pérez et al. 1994; Romero et al. 2004). In contrast, occlusal microwear consists of characteristic scratches and pits created by enamel contact with masticated food and tooth-to-tooth contact (Ryan 1979; Gordon 1988). Thus, the width, length, density, and distribution of scratches and pits on the occlusal surface vary widely among human groups depending on the properties of the foods (e.g., hardness and toughness), phytolith content, environmental abrasives, and food preparation techniques (Molleson et al. 1993; Teaford and Lytle 1996; Schmidt 2001; Teaford et al. 2001; El-Zaatari 2010; Schmidt et al. 2015) in addition to the compressive forces produced during mastication (Gordon 1982; King et al. 1999; Gügel et al. 2001; Mahoney 2006, 2007). Microwear turnover occurs quickly on occlusal surfaces (Teaford 1994)—the so-called the last supper effect—thus, documenting diet during the last days or weeks before an individual’s death (Grine 1986; Teaford and Tylenda 1991; Ungar 2011; Teaford et al. 2017). In contrast, the microwear pattern on the buccal surface has a more gradual turnover than the occlusal surface and stabilizes at maturity (Pérez-Pérez et al. 1994; Romero and De Juan 2007; Romero et al. 2012a). The differential turnover rates between occlusal and buccal surfaces offer a unique opportunity to assess dietary variability at short-(occlusal) and longer-term (buccal) intervals by individual and across groups.
Researchers have turned to deciduous dental microwear studies to assess their potential for distinguishing differences in foodstuffs consumed between different child age categories (Scott and Halcrow 2017; Fernández-Crespo et al. 2018; García-González et al. 2019). The above insights into differential microwear turnover rates are particularly interesting with respect to dental microwear analysis of infants and children. The majority of microwear studies focus on adult teeth and diets, despite the potential for non-adult microwear to provide novel insights into weaning and child feeding practices in the past (Scott and Halcrow 2017). Specific cultural practices whereby adults provision children with specific solid foods or with objects for “teething” could influence dental microwear. Within populations, access to different types of foods or the influences of specific cultural practices could be linked to important factors co-varying with biological age, different social age categories, or sex. All of these social and cultural determinants often correlate with aspects of biological and social maturation during childhood (Scott and Halcrow 2017). The aim of this paper is to delve into this poorly understood topic with an approach that combines inferences from buccal and occlusal microwear on deciduous molars among food-producing groups from the Neolithic to the Bronze Age of the Iberian Peninsula. The second aim is to assess intragroup variation in dental microwear related to dietary differences between children of different biologically defined age categories.
While the analysis of both the occlusal and buccal surfaces in the same groups is not a new approach (García-González et al. 2015; Pérez-Pérez et al. 2018), our analysis is the first to use both buccal and occlusal surfaces of the deciduous molars to better understand temporal and age-related differences in the diets of prehistoric children with the same microscope. Importantly, these analyses provide a non-destructive analytical framework to investigate aspects of feeding practices, food choice, and differential access to foods corresponding to specific age groups (Halcrow and Tayles 2008; Knudson and Stojanowski 2008; Mahoney et al. 2016; Mays 2016; Scott and Halcrow 2017; Ventresca Miller et al. 2017).
Materials and methods
Material
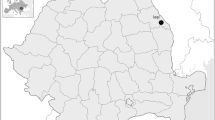
This study is based on the analysis of microwear from 44 deciduous molars belonging to four populations from the Iberian Peninsula (Late Neolithic: Cova de la Guineu, Chalcolithic: El Mirador, and Middle Bronze Age: Cova dels Galls Carboners and Valdavara (Fig. 1, Supplementary Table 1). Despite studying all relevant deciduous molars from each of these sites, final sample size is modest, 28 individuals for the buccal surface and 27 for the occlusal surface (Table 1). However, there are many factors contributing to the underrepresentation of child remains in the archeological record such as differential conservation, cultural treatment, and practices concerning deceased children, and/or excavation biases (Gibaja et al. 2010). The paucity of child remains from other Holocene sites in the Iberian Peninsula has been documented elsewhere (Daura et al. 2015, 2017; Fontanals-Coll et al. 2017) and is also reflected in the present study. Nevertheless, it should be noted that small sample size is typical and largely unavoidable, reality of research aimed at understanding child dietary patterns (e.g., Lalueza-Fox and Pérez-Pérez 1993; García-González et al. 2019; King et al. 2018; Eerkens et al. 2019) and dental microwear analyses of adult or fossil simple in general (Galbany and Pérez-Pérez 2004; Estebaranz et al. 2009).

Location of the four archeological populations within the Iberian Peninsula
Abbreviations are used to designate tooth types by jaw (upper or lower: U or L), side (left or right: L or R), and position in tooth row (first or second: 1 or 2) for each deciduous molars (dm).
The four sites included in this study are the following:
-
Cova de la Guineu: The collective burial site of Cova de la Guineu (Font-rubí, Barcelona, NE Iberian Peninsula) is dated to the late Neolithic as indicated by radiocarbon dates on human remains that yielded dates of 4110 ± 38 (4820–4454 cal. BP) and 4385 ± 32 (5040–4865 cal. BP) (Oms et al. 2016). The material culture is represented by few funerary furnishing (arrowheads, beads, and bell-shaped ceramics) given the number of individuals associated with the late Neolithic phase of the site (Morales et al. 2013; Oms et al. 2016). The dental remains from the sites provide a minimum number of 81 individuals, 24 of whom are children. Postdepositional damage limited analyses to 19 Ldm2 (Online supplementary Table 1).
-
El Mirador Cave: The cave of El Mirador is located on the southern slope of the Sierra de Atapuerca (Burgos, North Iberian Peninsula) at an altitude of 1.033 m a.s.l. El Mirador Cave has a collective inhumation with at least 23 individuals of both sexes and various ages. Two human bones yielded dates of 4000 ± 30 BP (4550–4390 cal. BP) and 4120 ± 30 BP (4880–4480 cal. BP) indicating that the burial episode occurred during the Chalcolithic (Reimer et al. 2013; Vergès et al. 2016). The dating and associated material culture indicate that the El Mirador population engaged in developed agriculture and stockbreeding (Cabanes et al. 2009; Vergès et al. 2016). Two individuals from El Mirador had well-preserved deciduous molars: an ULdm2 from a maxillary fragment belonging to a child around 6 years old at death and LRdm2 from a mandibular fragment belonging to a child of ~ 4 years of age at death (Online supplementary Table 1).
-
Valdavara: The cave of Valdavara is located in Becerreá (Lugo, NW Iberian Peninsula), about 600 m s.n.m. Valdavara is part of the karstic formations surrounding the municipality of Becerreá. Level 3 of the location known as Valdavara 2 yielded 55 human remains, including two infant individuals (a mandible and a hemi-maxilla) that were between 3 and 5 years old at death (Vaquero et al. 2009). There was no material culture in association with the human remains. Two of human bones were dated to 3250 ± 40 BP (3600–3360 cal. BP) and 3270 ± 40 years BP (3600–3400 cal. BP) corresponding to the Middle Bronze Age (Vaquero et al. 2009). Both individuals (URdm1 and LRdm2) were analyzed (Online supplementary Table 1).
-
The Galls Carboners Cave: The Galls Carboners Cave (Mont-Ral, Tarragona, NE Iberian Peninsula) opens onto a cliff on the north-eastern slope of the Motllats mountains, at 965 m a.s.l., on the western margin of the Brugent river valley (Vergès et al. 2017). An assemblage up to 1600 human remains, mainly postcranial bones, were recovered from the cave. A set of 349 human permanent and deciduous dental remains was analyzed and some deciduous teeth were chosen for dental microwear analysis. Specifically, 15 of the 18 Ldm2’s were free of postdepositional damage and included in analyses (Online supplementary Table 1). Apart from the human remains, ornamentation was found consistent with the character of burial site (Vergès et al. 2017). Radiocarbon dating on a human phalange dates this samples in the Middle Bronze Age 3310 ± 30 BP (3620–3460 cal. BP) (Vergès et al. 2017).
Age groupings are based on biological age following Halcrow and Tayles (2008): individuals aged from 0 to 4 years old correspond to the breastfeeding to terminal weaning stages, individuals aged from 5 to 7 years old are children that are still largely dependent on adults for feeding and protection, and individuals aged from 8 to 11 years old belong to the prepubertal period. Individual age-at-death was estimated using the timing of dental formation and the sequence of eruption of the teeth in each maxilla or mandible (Ferembach et al. 1980). In the case of isolated teeth, occlusal dental wear and resorption stages of the root were the basis for age determination (Schour and Massler 1941; Ferembach et al. 1980; AlQahtani et al. 2010).
Methods: Microwear analysis
The dental remains were observed directly using an environmental scanning electron microscope (ESEM: FEI Quanta 600) to analyze buccal and occlusal surfaces. Tooth orientation was standardized by keeping the buccal side at the top of both the ESEM monitor and in micrographs (Molleson et al. 1993). Both buccal and occlusal surfaces show dental microwear. However, there are some methodological differences related to the acquisition of the images depending on the surface studied.
For buccal microwear: Micrographs were taken at × 100 (1024 × 832 pixels) on the medial third of buccal surfaces. ESEM parameters were fixed at 20 kV acceleration voltage and low vacuum mode with a spot size of 4.5 and 0° of tilt angle. The working distance (WD) ranged between 15 and 25 mm (Pérez-Pérez et al. 1994; Galbany et al. 2004). Micrographs were enhanced with GIMP (https://www.gimp.org/) to increase contrast to facilitate wear feature characterization and measurement (Galbany et al. 2004). For each micrograph, an area of 0.56 mm2 was sharpened for buccal microwear feature analysis. All measured scratches start and end inside the field of view defined by the borders of the cropped image. A scratch was defined as a linear mark on the enamel surface, at least four times longer than its width (4:1), with a minimum length of 15 μm (Gordon 1982; Grine 1986). The total number of scratches (BTN) and their average length (BXT) in micrometers (μm) were computed, measured, and classified into four orientation categories from 0° to 180° (in 45° intervals) as horizontal (H), vertical (V), mesiodistal (MD), or distomesial (DM) (Pérez-Pérez et al. 1994) using Image J (Schneider et al. 2012). Two orientation indices were calculated: total horizontal/total number (TH/TN) and total vertical/total number (TV/TN) since they track broad differences in dietary patterns across groups (Lalueza et al. 1996).
For occlusal microwear: Micrographs were captured on facet 9—a crushing and grinding surface that experiences both compression and shearing during chewing (Kay and Hiiemae 1974)—which is located on the distobuccal cusp of the first and second molar (Maier and Schneck 1982; Mahoney 2006). Magnification was set to × 500 (1024 × 832 pixels). Images were enhanced and modified in the same way as the buccal surface micrographs (Galbany et al. 2004). Images were cropped at 700 × 500 pixels, which represents approximately 0.03 mm2 of the occlusal molar surface. Two kinds of features are present on the occlusal surface: scratches and pits (Ryan 1979; Gordon 1988). Scratches were defined and measured in the same manner as for buccal microwear, whereas pits are defined as features with a length to width ratio ≤ 4:1. As with buccal microwear, all measured features start and end inside the field of view defined by the borders of the cropped image. The total number of scratches (OTN) and their average length (OXT) in micrometers (μm) were computed and measured with ImageJ (Schneider et al. 2012).
Figure 2 shows a series of surfaces that illustrate buccal and occlusal microwear measurements and examples of surfaces that are excluded from the study because of the taphonomic or antemortem factors that obscure microwear.

A series of surfaces with different characteristics: a Well-preserved enamel on the buccal surface (× 100), striations in red; b Dietary wear in the occlusal surface (× 500), striations in red and pits in blue; c Surface affected by post-mortem damage and d surface with perikymata
Observer error: One author (RH) was responsible for the measurement of all microwear. In order to test intraobserver consistency in the analysis (Grine 1986; Galbany et al. 2005), five micrographs were analyzed three times each. Low values of standard errors and variance evidence the limited intraobserver variation in the analysis (Online Supplementary Table 4).
Small sample size required the use of non-parametric statistics, and Kruskal-Wallis tests were chosen for the intrapopulation comparisons with R commander statistical package (Fox 2005). As in other child microwear studies (García-González et al. 2019), we relied on overarching trends and data visualization rather than any statistical treatment for interpopulation comparisons.
Results
Microwear preservation and age determination
Microwear was present on 38 of 44 deciduous molars derived from the four study sites. Of the well-preserved teeth, microwear is preserved on 28 (63.6%) buccal and 27 (61.3%) occlusal surfaces (Table 1). Only 6 teeth (13.6%) preserved neither buccal nor occlusal microwear as a result of taphonomic modification or advanced occlusal wear (Table 1). A complete list of details (age categorization, estimated age at death, and surfaces preserving microwear) for all teeth screened for microwear is presented in Online Supplementary Table 1. Importantly, the sample is limited to a single tooth type (dm2) from the maxilla or mandible for a given individual, except for a single URdm1 from Valdavara (Online Supplementary Tables 1 and 2). The timing of upper and lower dm2 eruption is very similar (AlQahtani et al. 2010), thus variation in microwear (especially buccal) due to differences in eruption sequence across tooth types is negligible in the present study. Any differences between the URdm1 of Valdavara 145 and the dm2’s from the rest of the individuals will be discussed below.
Buccal microwear analysis
Descriptive statistics for buccal microwear by site are listed in Table 2 and complete datasets are provided in the Online Supplementary Table 2.
For the statistical analysis, Valdavara and El Mirador were not included. However, as the boxplot shows (Fig. 3), Valdavara has a high number of striations (BTN) compared with the other groups. Due to the high number of striations found on the buccal surfaces of Valdavara individuals, micrographs of the lingual surface were also explored. The lingual surfaces show a lack of striations which is to be expected since lingual surfaces are not usually affected by dietary wear (Fig. 4). Therefore, the high BTN for Valdavara is unlikely to be a taphonomic artifact. For the other two groups, Cova de la Guineu and Galls Carboners, no significant differences were observed for any variable (Table 3). In Fig. 5, the vertical and the horizontal index are plotted showing a high index of vertical striation rather horizontals.

Box and dot plots of total number (BTN) of buccal scratches by age category and archeological site. Box lines correspond to 25, 50 (median), and 75 percentiles of data. (Red Circles = 0-4 years; Blue Circles = 5-7 years; Green Circles = 8-11 years)

Micrographs of a buccal and b lingual surfaces of LRdm2 Valdavara individual. The lingual surfaces show a lack of striations which is to be expected since lingual surfaces are not usually affected by dietary wear

Plot of NH/TN index vs NV/TN index on the buccal surface for the four studied populations
We assessed whether intragroup differences exist among the different age categories for the larger samples from Cova dels Galls Carboners and Cova de la Guineu since El Mirador Cave and Valdavara samples are too small for similar comparisons. All three age categories (0–4, 5–7, and 8–11) were examined for Cova de la Guineu, but sample size (N = 9) for Cova dels Galls Carboners required condensing individuals into two age categories: 0–6 years and from 7 to 11 years (Table 4). No significant differences between the two Cova dels Galls Carboners age categories for BTN or BXT (Fig. 3, Table 5). In the case of Cova de la Guineu, significant differences between the three different age categories for BTN (F = 10.563; p value: 0.005) (Fig. 3, Table 5). A post hoc test shows that the 8–11-year-old children significantly differ from the rest of the children (0–4 and 5–7-year-old) from the Cova de la Guineu for BTN showing a high number of striations in 8–11-year-old children (Fig. 3, Tables 4 and 5). No significant differences between age categories for BXT by age category for either site (Table 5).
Occlusal microwear analysis
The descriptive statistics for the occlusal surface analysis by site are summarized in Table 6 and the complete datasets are available in Online Supplementary Table 3. No significant differences were between OTN and the site (Table 7). Likewise, no statistically significant differences were found between sites for OXT, mean number of pits, pit length, or pit width (Table 7).
As with the buccal surface, we assessed differences in occlusal microwear variation between age categories for Cova de la Guineu (0–4, 5–7, and 8–11) and Cova dels Galls Carboners (0–4 and 5–7) (Table 8). There are no significant differences between the two age categories at Cova dels Galls Carboners for OTN or OXT (Table 9). Similarly, no significant differences between occlusal OTN or OXT, respectively, for the three age categories represented at Cova de la Guineu (Table 9). For occlusal pitting variables, only the total number of pits are significantly different between the two age categories from Cova dels Galls Carboners (t = 4.048, p value 0.044) (Table 9, Fig. 6). There are fewer pits among the 0–4-year-old than the 5–7-year-old children. No other significant differences for pit length or width were shown for either site (Table 9).

Box and dot plots of total number of pits on occlusal surface by age category and archeological site. Box lines correspond to 25, 50 (median), and 75 percentiles of data. (Red Circles = 0-4 years; Blue Circles = 5-7 years; Green Circles = 8-11 years)
Discussion
The Valdavara case
Buccal microwear provides information about diet and food habits over a long period, due to the accumulation of microwear during life in conjunction with a rate slow turnover, of an individual lifespan in contrast to the occlusal pattern (Pérez-Pérez et al. 1994; Romero and De Juan 2007; Romero et al. 2012a). The individuals from Valdavara have substantially more buccal scratches than any other group (Fig. 3). However, the Valdavara sample is composed of only two individuals; therefore, we cannot extrapolate these results to a population level. Even so, the high number of scratches at Valdavara compared to the rest of the groups studied is interesting, since these two individuals belong to the youngest age category (0–4 years old)—a period characterized by breastfeeding and the eventual introduction of complementary foods until the end of the weaning (Halcrow and Tayles 2008; Meehan and Roulette 2013; Mays 2016; Scott and Halcrow 2017; Ventresca Miller et al. 2017; Mays et al. 2017). The high TN value corresponds to the abrasiveness of the diet (Romero et al. 2012b) related to the presence of exogenous particles like dust, sand, and ash (Pérez-Pérez et al. 1994; Polo-Cerdá et al. 2007; Romero et al. 2013) or the consumption of vegetables that contain phytoliths (Gügel et al. 2001). Given the difference in eruption sequence between dm1 and dm2, it is interesting to note that there are more buccal striations on the URdm1 of Valdavara 145 than the LRdm2 of Valdavara 146. This difference may be due to the longer time since eruption of the URdm1 compared to LRdm2 (AlQahtani et al. 2010).
We propose two potential explanations for the high buccal BTN result for Valdavara. First, the high abrasiveness of a diet could relate to a type and/or the degree of food processing at Valdavara that differs from the other sites. During the preparation of the food, the inclusion of the extrinsic particles from grindstones create extensive enamel microwear because the extrinsic particles are harder than enamel (Pérez-Pérez et al. 1994; Teaford and Lytle 1996; Gügel et al. 2001; Polo-Cerdá et al. 2007; Romero and De Juan 2007; Jarosová 2008; Galbany et al. 2008; Sołtysiak 2011; Schmidt et al. 2015; Schmidt et al. 2019). Thus, the use of grindstones may have been responsible for the high level of abrasiveness in the diet of Valdavara children. Although no archeological evidence of grindstones has been found at Valdavara (Vaquero et al. 2009), it is well known that Bronze Age populations from Northwestern Iberia used grindstones for crushing and milling cereals to obtain flour (Teira Brión and Amado 2014). The age of the two children from Valdavara suggests that they were near the end of their weaning period (Halcrow and Tayles 2008; Fulminante 2015) and probably consumed soft complementary food like porridge, gruels, or mashed food stuff (Sellen and Smay 2001; Meehan and Roulette 2013; Ventresca Miller et al. 2017). Although porridge is a soft and semiliquid food, many studies of ground tools made from sandstone, limestone, and basalt yield foods that retain mineral particles from milling (Teaford and Lytle 1996; Gügel et al. 2001; Polo-Cerdá et al. 2007; Mahoney et al. 2016; Mays 2016; Scott and Halcrow 2017; Ventresca Miller et al. 2017), which aligns well with our hypothesis about the dental microwear patterns documented at Valdavara.
Alternatively, the Valdavara children may have been consuming a harder diet, or more adult-like diet, compared to individuals from the other sites. Such diets would require more chewing time, therefore, generating a higher density of buccal scratches. In this case, other variables such as buccal BXT—which depend on many factors such as the pressure from the oral muscles or hardness of the food (Pérez-Pérez et al. 1994)—as well as the occlusal surface microwear need to be taken in to account. While the trend is not as clear as that for the buccal surface, there is a higher number of scratches and pits on the occlusal surface of the Valdavara teeth compared with the other groups. All of this evidence supports the assertion that the Valdavara children consumed foods with high-abrasive loads related to the consumption of milled foods with extrinsic abrasives or an adult-like diet requiring more mastication.
Although there are no adults from Valdavara that can be compared to the child sample, Aspe, a group of adults from the East of Iberian Peninsula (Alicante, Spain) from the same chronology as Valdavara (Bronze Age), has similar mean number and length of buccal scratches (BTN = 135; BXT = 126.83 μm) (Romero et al. 2004). A sample of Medieval children from La Olmeda (Palencia, Spain) have a microwear pattern with fewer buccal striations (BTN = 56.6; BXT = 151.231 μm) (Pérez-Pérez et al. 1994). Whereas, children from Alto de la Huesera (Late Neolithic, North-Cental Iberia) have striation densities that are similar to Valdavara (BNT = 108; BXT = 84.31 μm) (García-González et al. 2019). As a whole, this suggests that the prehistoric Iberian samples had more abrasive diets than the Medieval one (García-González et al. 2019).
The number of horizontal striations on the buccal surface is lower than the vertical ones in the child populations studied (Fig. 5). That is a common trend when it was compared with the prehistoric samples of Alto de la Huesera (NV/TN = 0.490; NH/TN = 0.037) and El Portalón (Sierra de Atapuerca, North-Cental Iberia) (NV/TN = 0.495; NH/TN = 0.028), (García-González et al. 2019) or Medieval samples (La Olmeda), 3–5-year-old children (NV/TN = 0.496; NH/TN = 0.197) and 6–9-year-old children (NV/TN = 0.469; NH/TN = 0.142) (Pérez-Pérez et al. 1994).
Ethnographic data also support the latter hypothesis since Ngandu farmers from the Central African Republic have a complete cessation of breastfeeding at 2 years old, despite this population tending to breastfeed for a longer time than pastoral and agricultural groups (Meehan and Roulette 2013). Sellen and Smay (2001) also suggest that preindustrial populations include solid complementary foods at an earlier age. Here we should bear in mind that, although deciduous enamel has different mechanical properties compared permanent enamel (De Menezes Oliveira et al. 2010), there is currently no consensus concerning differences in microwear formation between deciduous and permanent teeth (Krueger 2016; Mahoney et al. 2016; Kelly et al. 2020).
In summary, the combination of buccal and occlusal microwear patterning across the groups in this study suggests that the Valdavara group processed food for infants in a different way than other populations (Polo-Cerdá et al. 2007; Romero and De Juan 2007) or they were supplied with an adult-like diet. Such dietary patterns may be influenced by a wide variety of factors ranging from the death of a mother, cultural and social structure, or habitat constraints.
Intrapopulation variation by age category
There were no differences between the age categories for the Cova dels Galls Carboners group, but we did find greater buccal BTN in the older age categories at Cova de la Guineu. It is especially interesting that 8–11-year-old children have significant differences in the total number of buccal scratches compared to the rest of children at Cova de la Guineu. We suggest two possible explanations for these results. First, if we observe just the buccal surface, there is a significant difference between the 8–11-year-old children in comparison to the other two age groups, which may be attributed to eating similar foods to adults. Like in other dental microwear studies of child populations (Mahoney et al. 2016), the higher number of scratches among older children is possibly related to a more adult-like diet. Future analyses of adults from the Cova de la Guineu site will shed light on this hypothesis. Nonetheless, the total number of scratches of the 8–11-year-old group (97.25) is similar to the adult Neolithic populations from Vedrovice (85.94) in the Czech Republic (Jarosová 2008) and other adult populations from the from Bronze Age of the eastern Iberian Peninsula: Cova dels Blaus (114.66) and Cabezo Redondo (110.06) (Romero et al. 2004; Polo-Cerdá et al. 2007; Romero and De Juan 2007). The previously mentioned isotope studies from Alto de la Huesera also support this assertion (Fernández-Crespo et al. 2018).
However, the lack of differences in the occlusal facet microwear variables may be due to more rapid occlusal microwear turnover than on the buccal facet, the so-called the last supper effect, which reflects the diet of the last weeks of the individual (Grine 1986; Teaford 1994; Ungar 2011). Therefore, the greater differences in buccal microwear versus occlusal striations across age groups are probably due to a longer-term accumulation of scratches on the buccal surface which stabilizes at maturity (Pérez-Pérez et al. 1994; Romero and De Juan 2007; Romero et al. 2012a). This highlights a level of temporal resolution in dietary change across the lifespan of children that is difficult to elucidate through anything but destructive dietary analyses (e.g., incremental isotopic sampling). All of this supports our assertion, and that of others (García-González et al. 2015), that a more holistic picture of childhood dietary variability across age groups can be gained from analyses that integrate microwear from both surfaces.
With regard to occlusal pits (Fig. 6), the 0–4 age category from Cova dels Galls Carboners has a lower number of pits than that of the 5–7 age category. This could be explained by cultural differences in Cova dels Galls Carboners with the other groups in terms of dietary intake. The different food intake, like adults, among 5–7-year-old children of Cova dels Galls Carboners probably yielded a greater number of pits in comparison with the younger children, which could have consumed more processed foods. The higher, albeit not significantly different, number of occlusal scratches in the older age group also supports this finding.
Conclusion
Microwear analyses can provide remarkable insights into children’s feeding practices in the past that can be interpreted with respect to biological maturation and the social and cultural determinants of food choice and child feeding practices.
We found that buccal and occlusal microwear analyses provide complementary data on prehistoric diets since each surface provides different inferences about diet. We were able to detect differences through buccal surface analyses (e.g, the Valdavara case), as well as intrapopulation differences at Cova de la Guineu between the youngest and oldest children. Lastly, the intrapopulation differences among the Cova de la Guineu individuals show the utility of understanding differential turnover rates between buccal and occlusal surfaces where the long-term accumulation of buccal striations is reflected in older individuals at Cova de Galls Carboners versus the high density of pits associated with cultural differences.
References
AlQahtani S, Hector M, Liversidge H (2010) Brief communication: the London atlas of human tooth development and eruption. Am J Phys Anthropol 142:481–490. https://doi.org/10.1002/ajpa.21258
Armelagos GJ, Goodman AH, Jacobs KH (1991) The origins of agriculture: population growth during a period of declining health. Popul Environ 13:9–22
Bocquet-Appel J (2002) Paleoanthropological traces of a Neolithic demographic transition. Curr Anthropol 43:637–650. https://doi.org/10.1086/342429
Buikstra JE, Konigsberg LW, Bullington J (1986) Fertility and the development of agriculture in the prehistoric Midwest. Am Antiq 51:528–546
Cabanes D, Burjachs F, Expósito I et al (2009) Formation processes through archaeobotanical remains: the case of the bronze age levels in El Mirador cave, Sierra de Atapuerca, Spain. Quat Int 193:160–173. https://doi.org/10.1016/j.quaint.2007.08.002
Daura J, Sanz Borràs M, Arias O et al (2015) La Cova de l’Avi (Vallirana, Barcelona) y el inicio del Neolítico final en el Nordeste de la Península Ibérica. Inhumaciones colectivas y nuevas redes de intercambio. Trab Prehist 72:327–341
Daura J, Sanz M, Soriano I et al (2017) Objetos de oro y epicampaniforme en la Cova del Gegant. Relaciones en la costa mediterránea de la Península Ibérica durante la Edad del Bronce. Trab Prehist 74:149–167
De Menezes Oliveira MAH, Torres CP, Gomes-Silva JM et al (2010) Microstructure and mineral composition of dental enamel of permanent and deciduous teeth. Microsc Res Tech 73:572–577
Dhavale N, Halcrow SE, Buckley HR et al (2017) Linear and appositional growth in infants and children from the prehistoric settlement of Ban Non Wat, Northeast Thailand: evaluating biological responses to agricultural intensification in Southeast Asia. J Archaeol Sci Rep 11:435–446
Eerkens JW, Bartelink EJ, Bartel J, Johnson PR (2019) Isotopic insights into dietary life history, social status, and food sharing in American Samoa. Am Antiq 84(2):336–352.
El-Zaatari S (2010) Occlusal microwear texture analysis and the diets of historical/prehistoric hunter-gatherers. Int J Osteoarchaeol 20:67–87. https://doi.org/10.1002/oa.1027
Estalrrich A, El Zaatari S, Rosas A (2017) Dietary reconstruction of the El Sidrón Neandertal familial group (Spain) in the context of other Neandertal and modern hunter-gatherer groups. A molar microwear texture analysis. J Hum Evol 104:13–22. https://doi.org/10.1016/j.jhevol.2016.12.003
Estebaranz F, Martínez LM, Galbany J, Turbón D, Pérez-Pérez A (2009) Testing hypotheses of dietary reconstruction from buccal dental microwear in Australopithecus afarensis. J Hum Evol 57(6):739–750
Ferembach D, Schwindezky I, Stoukal M (1980) Recommendation for age and sex diagnoses of skeletons. J Hum Evol 9:517–549
Fernández-Crespo T, Czermak A, Lee-Thorp JA, Schulting RJ (2018) Infant and childhood diet at the passage tomb of Alto de la Huesera (north-central Iberia) from bone collagen and sequential dentine isotope composition. Int J Osteoarchaeol 28:542–551
Fontanals-Coll M, Eulàlia Subirà M, Díaz-Zorita Bonilla M, Gibaja JF (2017) First insight into the Neolithic subsistence economy in the north-east Iberian Peninsula: paleodietary reconstruction through stable isotopes. Am J Phys Anthropol 162:36–50
Fox J (2005) Getting started with the R commander: a basic-statistics graphical user interface to R. J Stat Softw 14:1–42
Fulminante F (2015) Infant feeding practices in Europe and the Mediterranean from prehistory to the middle ages: a comparison between the historical sources and bioarchaeology AU - Fulminante, Francesca. Child Past 8:24–47. https://doi.org/10.1179/1758571615Z.00000000026
Galbany J, Martínez Martínez L, Pérez-Pérez A (2004) Tooth replication techniques, SEM imaging and microwear analysis in primates: methodological obstacles. Anthropologie 42(1):5
Galbany J, Pérez-Pérez A (2004) Buccal enamel microwear variability in Cercopithecoidea primates as a reflection of dietary habits in forested and open savanna environments. Anthropologie 42(1):13–20
Galbany J, Martínez LM, López-Amor HM, Espurz V, Hiraldo O, Romero A, de Juan J, Pérez-Pérez A (2005) Error rates in buccal-dental microwear quantification using scanning electron microscopy. Scanning 27(1):23–29
Galbany J, Garriga N, Majoral-Salichs M et al (2008) Microdesgaste y patología dental en la población de la Edad de Bronce de “Mar i Muntanya” (Alella, Barcelona). Rev Esp Antrop Fís 28:25–36
García-González R, Carretero JM, Richards MP et al (2015) Dietary inferences through dental microwear and isotope analyses of the Lower Magdalenian individual from El Mirón Cave (Cantabria, Spain). J Archaeol Sci 60:28–38. https://doi.org/10.1016/j.jas.2015.03.020
García-González R, Sánchez-Puente Z, Arsuaga JL, Carretero JM (2019) Dietary inferences from dental microwear patterns in Chalcolithic populations from the Iberian Peninsula: the case of El Portalón de Cueva Mayor (Sierra de Atapuerca, Burgos, Spain) and El Alto de la Huesera (Álava, Spain). Archaeol Anthropol Sci 11(8):3811–3823. https://doi.org/10.1007/s12520-018-0711-x
Gibaja JF, Majó T, Chambon P, et al (2010) Prácticas funerarias durante el Neolítico. Los enterramientos infantiles en el noreste de la Península Ibérica
Gordon KD (1988) A review of methodology and quantification in dental microwear analysis. Scanning Microsc 2:1139–1147
Gordon KD (1982) A study of microwear on chimpanzee molars: implications for dental microwear analysis. Am J Phys Anthropol 59:195–215
Grine FE (1986) Dental evidence for dietary differences in Australopithecus and Paranthropus: a quantitative analysis of permanent molar microwear. J Hum Evol. https://doi.org/10.1016/S0047-2484(86)80010-0
Gügel IL, Grupe G, Kunzelmann K-H (2001) Simulation of dental microwear: characteristic traces by opal phytoliths give clues to ancient human dietary behavior. Am J Phys Anthr 114:124–138. https://doi.org/10.1002/1096-8644(200102)114:2<124::AID-AJPA1012>3.0.CO;2-S
Halcrow SE, Tayles N (2008) The bioarchaeological investigation of childhood and social age: problems and prospects. J Archaeol Method Theory 15:190–215. https://doi.org/10.1007/s10816-008-9052-x
Han CS, Martin MA, Dichosa AEK et al (2016) Salivary microbiomes of indigenous Tsimane mothers and infants are distinct despite frequent premastication. PeerJ 4:e2660
Howcroft R (2013) Weaned upon a time : studies of the infant diet in prehistory. Doctoral dissertation, Department of Archaeology and Classical Studies, Stockholm University.
Jarosová I (2008) Dietary inferences using buccal microwear analysis on the LBK population from Vedrovice, Czech Republic. Anthropologie 46:175
Kay RF, Hiiemae KM (1974) Jaw movement and tooth use in recent and fossil primates. Am J Phys Anthropol 40:227–256. https://doi.org/10.1002/ajpa.1330400210
Kelly CD, Schmidt CW, D´Anastasio R (2020) Dental microwear texture analysis in deciduous teeth. In: Dental Wear in Evolutionary and Biocultural Contexts (pp. 169-186). Academic Press.
King C, Snoddy AM, Millard AR et al (2018) A multifaceted approach towards interpreting early life experience and infant feeding practices in the ancient Atacama Desert, Northern Chile. Int J Osteoarchaeol 28(5):599–612. https://doi.org/10.1002/oa.2671
King T, Aiello LC, Andrews P (1999) Dental microwear of Griphopithecus alpani. Acad Press J Hum Evol 36:3–31. https://doi.org/10.1006/jhev.1998.0258
Knudson K, Stojanowski C (2008) New directions in bioarchaeology: recent contributions to the study of human social identities. J Archaeol Res 16(4):397–432. https://doi.org/10.1007/s10814-008-9024-4
Krueger KL (2016) Dental microwear texture differences between permanent and deciduous enamel. In: the 85th annual meeting of American Association of Physical Anthropologists
Lalueza-Fox C, Pérez-Pérez (1993) The diet of the Neanderthal Child Gibraltar 2 (Devil’s Tower) through the study of the vestibular striation pattern. J Hum Evol 24(1):29–41
Lalueza C, Péréz-Perez A, Turbón D (1996) Dietary inferences through buccal microwear analysis of Middle and Upper Pleistocene human fossils. Am J Phys Anthropol 100:367–387. https://doi.org/10.1002/(SICI)1096-8644(199607)100:3<367::AID-AJPA5>3.0.CO;2-R
Mahoney P (2006) Dental microwear from Natufian hunter-gatherers and early neolithic farmers: comparisons within and between samples. Am J Phys Anthropol. https://doi.org/10.1002/ajpa.20311
Mahoney P (2007) Human dental microwear from Ohalo II (22,500-23,500 cal BP), southern Levant. Am J Phys Anthropol. https://doi.org/10.1002/ajpa.20548
Mahoney P, Schmidt C, Deter C et al (2016) Deciduous enamel 3D microwear texture analysis as an indicator of childhood diet in medieval Canterbury, England. J Archaeol Sci 66:128–136. https://doi.org/10.1016/j.jas.2016.01.007
Maier W, Schneck G (1982) Functional morphology of hominoid dentitions. J Hum Evol 11:693–696. https://doi.org/10.1016/S0047-2484(82)80057-2
Mays S (2016) A study of the potential of deciduous incisor wear as an indicator of weaning using a human skeletal population. Int J Osteoarchaeol 26:725–731. https://doi.org/10.1002/oa.2464
Mays S, Gowland R, Halcrow S, Murphy E (2017) Child bioarchaeology: perspectives on the past 10 years. Child Past:1–19. https://doi.org/10.1080/17585716.2017.1301066
Meehan CL, Roulette JW (2013) Early supplementary feeding among central African foragers and farmers: a biocultural approach. Soc Sci Med 96:112–120. https://doi.org/10.1016/j.socscimed.2013.07.029
Molleson T, Jones K, Jones S (1993) Dietary change and the effects of food preparation on microwear patterns in the Late Neolithic of Abu Hureyra, northern Syria. J Hum Evol 24:455–468
Morales JI, Cebrià A, Mestres J, et al (2013) La Cova de la Guineu. 12.000 anys de presència humana a les capçaleres del Foix. III Monogr del Foix 172–183
Oms FX, Cebrià A, Mestres J, et al (2016) Campaniforme i metal· lúrgia en un espai sepulcral del III mil· lenni cal. BC: la Cova de la Guineu (Font-rubí, Alt Penedès). Jornades d’Arqueologia del Penedès 109–116
Pearson JA, Hedges REM, Molleson TI, Özbek M (2010) Exploring the relationship between weaning and infant mortality: an isotope case study from Aşıklı Höyük and Çayönü Tepesi. Am J Phys Anthropol 143:448–457
Pelto GH, Zhang Y, Habicht J (2010) Premastication: the second arm of infant and young child feeding for health and survival? Matern Child Nutr 6:4–18
Pérez-Pérez A, Espurz V, Bermúdez de Castro JM et al (2003) Non-occlusal dental microwear variability in a sample of Middle and Late Pleistocene human populations from Europe and the Near East. J Hum Evol 44:497–513. https://doi.org/10.1016/S0047-2484(03)00030-7
Pérez-Pérez A, Lalueza C, Turbón D (1994) Intraindividual and intragroup variability of buccal tooth striation pattern. Am J Phys Anthropol 94:175–187. https://doi.org/10.1002/ajpa.1330940203
Pérez-Pérez A, Martínez LM, Gómez M et al (2018) Correlations among dietary proxies in African fossil hominins: dental buccal microwear, occlusal textures and 13C stable isotope. J Archaeol Sci Rep. https://doi.org/10.1016/j.jasrep.2018.03.013
Pinhasi R, Stock JT (2011) Human bioarchaeology of the transition to agriculture. Eds: John Wiley & Sons.
Polo-Cerdá M, Romero A, Casabó J, De Juan J (2007) The bronze age burials from Cova Dels Blaus (Vall d’Uixó, Castelló, Spain): an approach to palaeodietary reconstruction through dental pathology, occlusal wear and buccal microwear patterns. HOMO - J Comp Hum Biol 58:297–307. https://doi.org/10.1016/j.jchb.2006.10.005
Reimer PJ, Bard E, Bayliss A et al (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55:1869–1887
Romero A, De Juan J (2007) Intra- and interpopulation human buccal tooth surface microwear analysis: inferences about diet and formation processes. Anthropologie 45:61–70
Romero A, Galbany J, De Juan J, Pérez-Pérez A (2012a) Brief communication: short- and long-term in vivo human buccal-dental microwear turnover. Am J Phys Anthropol 148:467–472. https://doi.org/10.1002/ajpa.22054
Romero A, Galbany J, Juan D, Pe A (2012b) Brief communication : short- and long-term in vivo human buccal – dental microwear turnover 000:0–5. https://doi.org/10.1002/ajpa.22054
Romero A, Martínez-Ruiz N, De Juan J (2004) Non-occlusal dental microwear in a Bronze-Age human sample from East Spain. Anthropologie 42:65–70
Romero A, Ramírez-Rozzi FV, De Juan J, Pérez-Pérez A (2013) Diet-related buccal dental microwear patterns in central African pygmy foragers and bantu-speaking farmer and pastoralist populations. PLoS One. https://doi.org/10.1371/journal.pone.0084804
Ryan AS (1979) Wear striation direction on primate teeth: a scanning electron microscope examination. Am J Phys Anthropol 50:155–167. https://doi.org/10.1002/ajpa.1330500204
Scharlotta I, Goude G, Herrscher E et al (2018) Shifting weaning practices in Early Neolithic Cis-Baikal. Siberia 28:579–598. https://doi.org/10.1002/oa.2708
Schmidt CW (2001) Dental microwear evidence for a dietary shift between two nonmaize-reliant prehistoric human populations from Indiana. Am J Phys Anthropol. https://doi.org/10.1002/1096-8644(200102)114:2<139::AID-AJPA1013>3.0.CO;2-9
Schmidt CW, Beach JJ, McKinley JI, Eng JT (2015) Distinguishing dietary indicators of pastoralists and agriculturists via dental microwear texture analysis. Surf Topogr Metrol Prop 4:14008
Schmidt CW, Remy A, Van Sessen R et al (2019) Dental microwear texture analysis of Homo sapiens sapiens: foragers, farmers, and pastoralists. Am J Phys Anthropol:1–20. https://doi.org/10.1002/ajpa.23815
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9:671
Schour I, Massler M (1941) The development of the human dentition. J Am Dent Assoc 28:1153–1160
Scott RM, Halcrow SE (2017) Investigating weaning using dental microwear analysis: a review. J Archaeol Sci Rep 11:1–11. https://doi.org/10.1016/j.jasrep.2016.11.026
Sellen DW, Smay DB (2001) Relationship between subsistence and age at weaning in “preindustrial” societies. Hum Nat 12:47–87
Sołtysiak A (2011) Cereal grinding technology in ancient Mesopotamia: evidence from dental microwear. J Archaeol Sci 38:2805–2810. https://doi.org/10.1016/j.jas.2011.06.025
Tausch J, Kullmer O, Bromage TG (2015) A new method for determining the 3D spatial orientation of molar microwear. Scanning 37(6):446–457
Teaford MF (1994) Dental microwear and dental function. Evol Anthropol Issues News Rev 3:17–30. https://doi.org/10.1002/evan.1360030107
Teaford MF, Larsen CS, Pastor RF, Noble VE (2001) Pits and scratches: microscopic evidence of tooth use and masticatory behavior in La Florida. Bioarchaeol Span Florida Impact Colon Univ Press Florida, Gainesv 82–112
Teaford MF, Lytle JD (1996) Brief communication: diet-induced changes in rates of human tooth microwear: a case study involving stone-ground maize. Am J Phys Anthropol 100:143–147
Teaford MF, Ungar PS, Taylor AB et al (2017) In vivo rates of dental microwear formation in laboratory primates fed different food items. Biosurf Biotribol 3:166
Teaford MF, Tylenda CA (1991) A new approach to the study of tooth wear. J Dent Res 70(3):204–207
Teira Brión A, Amado E (2014) Molinos fuera de lugar. Fronteras y contextos de la molienda en la arqueología de la Edad del Hierro del noroeste ibérico Rev d’Arqueologia Ponent 24:271–287
Ungar PS (2011) Dental evidence for the diets of Plio-Pleistocene hominins. Am J Phys Anthropol 146:47–62. https://doi.org/10.1002/ajpa.21610
Vaquero M, Alonso Fernández S, Alonso Fernández C et al (2009) New radiometric dates for the prehistory of northwestern Iberia: Valdavara cave (Becerreá, Lugo). Trab Prehist 66:99–113
Ventresca Miller A, Hanks BK, Judd M, Epimakhov A, Razhev D (2017) Weaning practices among pastoralists: new evidence of infant feeding patterns from Bronze Age Eurasia. Am J Phys Anthropol 162:409–422. https://doi.org/10.1002/ajpa.23126
Vergès JM, Allué E, Fontanals M et al (2016) El Mirador cave (Sierra de Atapuerca, Burgos, Spain): a whole perspective. Quat Int 414:236–243
Vergès JM, Munoz L, Pedro M et al (2017) La cova dels Galls Carboners (Mont-ral, Alt Camp), una cavitat dínhumació col·lectiva durant l’edat del Bronze. Butlletí Arqueol V 38-39:17–43
Waterman AJ, Silva AM, Tykot RH (2014) Stable isotopic indicators of diet from two late prehistoric burial sites in Portugal: an investigation of dietary evidence of social differentiation. Open J Archaeom 2:1. https://doi.org/10.4081/arc.2014.5258
Waters-Rist AL, Bazaliiskii VI, Weber AW, Katzenberg MA (2011) Infant and child diet in Neolithic hunter-fisher-gatherers from cis-baikal, Siberia: intra-long bone stable nitrogen and carbon isotope ratios. Am J Phys Anthropol 146:225–241
Acknowledgments
Special thanks to the three anonymous reviewers and associate editor for comments and suggestions that considerably improved this manuscript. We would like to thank Andrés Teira Brión for his useful comments on the Galician Bronze Age and also to José Ramón Rabuñal for his helpful suggestions. Thanks also to the El Mirador Cave, Cova de la Guineu, Galls Carboners, and Valdavara excavation teams. ESEM analyses were conducted at the Scientific and Technical Resources Service of the University Rovira i Virgili, Tarragona, Spain.
Funding
This work has been funded by the Dirección General de Investigación of M.E.C, project numbers PGC2018-093925-B-C32, CGL2015-65387-C03-1-P (MINECO/FEDER), the Junta de Castilla y León and CERCA Programme/Generalitat de Catalunya, AGAUR 2017 SGR 1040 and AGAUR 2017- SGR 00011. MINECO 2017-HAR 86509. The Galls Carboners and Cova de la Guineu excavations are funded by the 2014/100574 and the CLT009/18/00024 projects of the Culture Department of the Generalitat de Catalunya. The Valdavara excavation was funded by the Concello de Becerreá. JCW was funded by a Marie Skłodowska-Curie Actions (H2020-MSCA-IF-2016 No. 749188) and JIM by Juan de la Cierva – Incorporación (IJCI-2017-31445) R.H is beneficiary of PhD research fellowship Martí i Franquès (2019PMF-PIPF-59).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 133 kb)
Rights and permissions
About this article

Cite this article
Hernando, R., Willman, J.C., Vergès, J.M. et al. Inferring childhood dietary maturation using buccal and occlusal deciduous molar microwear: a case study from the recent prehistory of the Iberian Peninsula. Archaeol Anthropol Sci 12, 30 (2020). https://doi.org/10.1007/s12520-019-00997-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-019-00997-z