
Abstract
Coastal mangrove–freshwater marsh ecotones of the Everglades represent transitions between marine salt-tolerant halophytic and freshwater salt-intolerant glycophytic communities. It is hypothesized here that a self-reinforcing feedback, termed a “vegetation switch,” between vegetation and soil salinity, helps maintain the sharp mangrove–marsh ecotone. A general theoretical implication of the switch mechanism is that the ecotone will be stable to small disturbances but vulnerable to rapid regime shifts from large disturbances, such as storm surges, which could cause large spatial displacements of the ecotone. We develop a simulation model to describe the vegetation switch mechanism. The model couples vegetation dynamics and hydrologic processes. The key factors in the model are the amount of salt-water intrusion into the freshwater wetland and the passive transport of mangrove (e.g., Rhizophora mangle) viviparous seeds or propagules. Results from the model simulations indicate that a regime shift from freshwater marsh to mangroves is sensitive to the duration of soil salinization through storm surge overwash and to the density of mangrove propagules or seedlings transported into the marsh. We parameterized our model with empirical hydrologic data collected from the period 2000–2010 at one mangrove–marsh ecotone location in southwestern Florida to forecast possible long-term effects of Hurricane Wilma (24 October 2005). The model indicated that the effects of that storm surge were too weak to trigger a regime shift at the sites we studied, 50 km south of the Hurricane Wilma eyewall, but simulations with more severe artificial disturbances were capable of causing substantial regime shifts.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In wave-protected shorelines of southern Florida, mangroves are the dominant wetland ecosystem (Lugo and Snedaker 1974; Bertness 2007; Zhang et al. 2012). These mangrove communities usually form landward transition zones (ecotones) with freshwater marshes. The mangroves of southern coastal Florida have been observed to be expanding inland at the expense of both salt marsh and freshwater marsh over several decades (Ball 1980; Ross et al. 2000; Doyle et al. 2003; Gaiser et al. 2006; Krauss et al. 2011; Raabe et al. 2012). Sea level rise of more than 20 cm since 1930 (Maul and Martin 1993) is thought to be the primary driver of the changes (Krauss et al. 2011; Saha et al. 2011). In his classic study, Egler (1952) documented distinct zonation bands of vegetation, primarily from mangroves to sawgrass marsh, along a transect from the coast inland. He noted the movement of Rhizophora mangle into Cladium jamaicense marsh, facilitated by storm tides that carry both salinity and Rhizophora seedlings far inland on the flat landscape. Ross et al. (2000) revisited the sites of Egler’s study and found that, during the past 40 years, a landward shift in the bands had occurred. In particular, vegetation in a zone termed the “white zone” (band 4 in Egler 1952) had shifted from a mixture of C. jamaicense and R. mangle to pure R. mangle. The authors attributed this change to sea level rise and water management practices that have reduced freshwater flow to the southern Everglades. Landward movement of mangroves into freshwater marsh in other areas of the southwestern Everglades has been identified through aerial photographs (Smith III et al. 2013).
Much of what has been documented concerning the spatial and temporal patterns of vegetation along the marine to terrestrial transect in southern Florida can be explained in terms of familiar ecological concepts. Higher tolerance to salinity favors mangrove species dominance at the marine end of this transect, while competition favors dominance by glycophytic species at the terrestrial (freshwater) end of the transect. Gradual rise in sea level and anthropogenic decreases in freshwater flow cause salinity intrusion and a landward shift of the vegetation bands, with halophytic mangroves replacing glycophytic freshwater marsh and hardwood hammocks. Other factors, such as changes in fire frequency, may also be affecting the ecotones between some of these bands (Egler 1952).
An additional factor, however, may also play a role in the dynamics of the shifting ecotones: positive feedbacks between a given vegetation type and its local soil conditions. Sternberg et al. (2007) hypothesized that feedback effects of two vegetation types, mangroves and hardwood hammocks, on local soil salinity help maintain the sharp ecotone frequently observed between them. Although mangrove vegetation is outcompeted by glycophytic vegetation under less saline conditions inland, mangroves, once established, can influence vadose zone salinity in their favor. A suite of salt tolerance mechanisms in mangroves allows them to transpire during periods of higher salinity, despite having rather conservative rates of water use overall. This facilitates higher evapotranspiration rates than would occur with the presence of only glycophytic vegetation. Mangrove roots access to water allows saline groundwater to move up in the vadose zone, further increasing its salinity. Thus, mangroves create locally high soil salinity conditions, while neighboring glycophytic vegetation, which downregulates evapotranspiration during the dry season, maintains locally low soil salinity. Each vegetation type creates soil conditions favoring itself over the other type. The resulting sharp change in soil salinity from one vegetation type to the other is confirmed by observations (Sternberg et al. 2007; Saha et al. 2011). This is an example of the more general “vegetation switch” mechanism (Agnew et al. 1993), which has been used to explain sharp ecotones between differing vegetation types, such as forest–grassland, forest–mire, Alpine treelines, etc. The vegetation switch can be thought of as a form of vegetation pattern generation or self-organization, that is, it arises through internal interactions to create structure (e.g., Temmerman et al. 2005, 2007; Kirwan and Murray 2007).
There are several implications of the existence of such a vegetation switch for ecotones in general and the halophyte–glycophyte ecotone in particular. First, as noted above, this positive feedback mechanism should act to sharpen ecotones between the vegetation types. Second, because each vegetation type has a positive effect on its local soil conditions that favor itself, the spatial position of the ecotone should be fairly resilient to small disturbances. For example, a minor disturbance that changes soil salinity by only a small amount might be counteracted by the local vegetation. The third implication is that, although the ecotone between the two vegetation types is sharp, its exact location on the marine–terrestrial transect may not be deterministic. The underlying abiotic conditions at a given point along a segment of the transect may be suitable for either vegetation type to thrive, but, once established at some point on that segment, one of the vegetation types can influence those conditions such that it can exclude the other type. The segment of the transect along which either vegetation type could exist in the absence of the other is called a region of bistability, and the two vegetation types that could each dominate the region are called alternative stable states (Scheffer 1997; Beisner et al. 2003; Folke et al. 2004). The exact location of the ecotone may depend on initial conditions. A fourth implication of the vegetation switch hypothesis is that a sufficiently large disturbance may be able to overcome the resilience of the ecotone in the region of bistability, causing a shift, called a regime shift, from the existing stable vegetation state on one side of the ecotone to the alternative stable vegetation state. For a regime shift to occur, however, the disturbance would have to exceed some threshold in strength and duration of the change in environmental conditions and be accompanied by input of propagules of the alternative vegetation type. But once a regime shift occurs, it may be difficult to reverse due to hysteresis. A fifth implication is that a mixture of the alternative vegetation types is unstable, and the system will move toward dominance of one or the other type, or possibly toward segregation of the two types on either side of a sharp ecotone. This transition may be slow, however, depending on the rates of competitive exclusion.
The existence of a vegetation switch mechanism in the mangrove–freshwater marsh ecotone zone changes the sort of dynamics that can occur when external conditions change. For example, a gradual rise in sea level might not lead to gradual landward movement of vegetation bands, as the vegetation types are self-maintaining and the ecotones would thus be resilient to small changes in sea level. The switch mechanism suggests that ecotones may be stable much of the time but be punctuated by large jumps in space when major disturbances occur.
The hypothesis of Sternberg et al. (2007), specifically the sharp mangrove–hardwood hammock ecotone, has been supported through simulation modeling (Teh et al. 2008; Jiang et al. 2012a), consistent with the idea of a vegetation switch mechanism. If the other implications of the switch mechanism also apply to the transition between halophytic and glycophytic vegetation, then we expect there to be segments of the transition that are bistable and that could undergo regime shifts from their current vegetation state to an alternative vegetation, if sufficiently large disturbances occurred. In particular, a storm surge with large overwash of saline water might shift a region currently covered with glycophytic vegetation to undergo a shift to halophytic vegetation. Ocean water intrusion through storm surges may affect large areas on a short time scale (e.g., cause a short-term salinity increase in the soil and groundwater of an inundated area). Whether such storm-related pulses lead to long-term effects on vegetation depends on many specific factors, such as the physiological and competitive properties of local vegetation, precipitation, overland freshwater flow, elevation gradient, depth and salinity of groundwater, and sufficient input of seedlings of the alternative vegetation type (Jiang et al. 2012a; White and Falkland 2010). If these conditions are satisfied, the possibility of a regime shift over a broad area affected by a storm surge exists, which is why storm surges are of special interest in relation to mangrove migration and other vegetation changes (Ross et al. 2009; Teh et al. 2008). There have been some relevant studies on transitions from glycophytic to halophytic vegetation types due to salinity inputs. For example, Baldwin and Mendelssohn (1998) studied the effects of a pulse of salinity and inundation coupled with clipping of aboveground vegetation on two adjoining plant communities, Spartina patens and Sagittaria lancifolia. The study concluded that the vegetation might shift to a salt-tolerant or flood-tolerant species, depending on the level of flooding and salinity at the time of disturbance. Also see Person and Ruess (2003) and Steyer et al. (2010) for relevant studies.
The mangrove–freshwater marsh (Cladium) ecotone is often very sharp, which suggests that positive feedback effects similar to those at the mangrove–hardwood hammock boundary, and thus a switch mechanism may occur. The question we ask here is whether regime shift through salinity overwash is a feasible mechanism for explaining some of the past inland movement of the boundary in the coastal Everglades of southern Florida, or for influencing future movement. The hypothesis of a regime shift requires two primary prerequisites: an increase in salinities and an invasion of mangrove propagules following a storm surge, or alternatively a prior presence of propagules. Through an analysis of the effects of increased salinity and mangrove propagules with a model based on a regime shift theory, we demonstrate how shifts resulting in long-term changes in the mangrove–marsh ecotone could occur after storm surge events and inquire whether, on the basis of empirical data, they are likely to occur in a particular region of the Everglades affected by a hurricane. First, coastal water level and salinity data from 2000–2010 are used as a hydrological baseline and to observe possible trends in increased coastal groundwater salinity. Second, vegetation dynamics are simulated based on hydrology data from Hurricane Wilma (Oct 2005). Third, long-term vegetation changes under given scenarios of salinity intrusion and the density of mangrove propagules transported to freshwater marsh sites by hypothetical storm surge are simulated.
Methods
An ecotone model is first developed to describe possible mechanisms that can maintain a sharp ecotone and, by extension, the possibility of regime shifts causing a discrete jump in the location of the ecotone. The model is applied to a single USGS study site on a transect across the mangrove–freshwater marsh ecotone. While much relevant data are available at this site, only rough estimates of some model parameters are available at this time, so parameters that need further research are identified.
Study Site
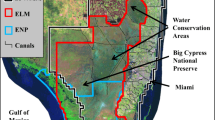
The mangrove–marsh ecotone studied is adjacent to the Harney River estuary (25°25' N, 81°03' W), a distributary of the Shark River estuary from the freshwater Everglades into the Gulf of Mexico (Fig. 1). Two U.S. Geological Survey (USGS) paired surface/groundwater gauges along the Harney River bookend a north–south transect. Shark 4 (SH4) is located ∼30 m south of the Harney River, within a mangrove fringe forest and USGS gauge Shark 5 (SH5) is located ∼300 m south in an interior coastal marsh. The SH4–SH5 transect is located on the middle reach of the estuary, about 9.5 km from the Gulf of Mexico. Historical aerial images show there has been some mangrove migration into the freshwater marsh habitat in this area of the southwestern Everglades, occurring between the late 1920s and early 1940s (Smith III et al. 2013).

Study area of mangrove–marsh transition. SH4 is USGS gauge Shark 4, located in mangrove fringe forest; SH5 is USGS gauge Shark 5, located in sawgrass; HR is Everglades National Park gauge on Harney River. The inset graph shows study area and Hurricane Wilma pathway
Model Spatial Scale
The model is based on a grid of 10 × 10 m spatial cells. Within each cell, water and salinity dynamics, as well as vegetation dynamics are described.
Model Water Budget and Salinity
The water budget was computed as a water balance on a daily basis. Vadose or rooting zone salinity, assumed homogeneous within a given spatial cell, is determined by infiltration rate, which is the difference between the precipitation, P, tidal input, T, and evapotranspiration, E. A groundwater lens, whose salinity can vary, sits on top of underlying seawater. Salinity of this groundwater lens in the same given spatial cell is determined by the difference between the infiltration rate and the daily change in the surface height of this groundwater. Infiltration rate (I NF) is calculated as follows,
The dynamics of salinity in the rooting zone are the same as used by Sternberg et al. (2007),
where ρ is the porosity, z is rooting zone height, and S V and S GW are the salinities of the pore water in the rooting zone and of the underlying groundwater, respectively, and I NF has units of distance per unit time. Positive values of infiltration indicate capillary rising from the groundwater to the rooting zone. Conversely, negative values of infiltration indicate percolation downward into the groundwater. The underlying saline ocean water, on which the groundwater sits, is assumed to be 35 (S O). The dynamics of salinity in the groundwater are given by the equations,
where H GW is the groundwater gauge level. When the groundwater level changes to compensate upward infiltration to the rooting zone (i.e., \( \left({I}_{\mathrm{NF}}+\rho \frac{{\mathrm{dH}}_{\mathrm{GW}}}{ dt}\right) \) is positive), additional water from underlying saline ocean water will move into the groundwater. Groundwater salinities do not affect the much larger pool of underlying saline ocean water.
To parameterize the model, water level or stage data (NAVD 88) were obtained from USGS paired surface/groundwater gauges SH4 and SH5. Daily water level changes were calculated as the difference from 1 day to the next. Daily rainfall data were obtained from the USGS Everglades Depth Network (EDEN) website (http://sofia.usgs.gov/eden/). Monthly averages and standard deviations were used to generate precipitation input for the model. Tidal height and salinity data were obtained from a nearby Everglades National Park gauge, Harney River (HR). The evapotranspiration, E, used in the model was calculated by multiplying potential evapotranspiration (PET) by salinity effect (f S). Daily PET data, which represent the evapotranspiration rate without moisture or salinity limitation, were obtained from the EDEN website. Evapotranspiration reductions due to salinity effect (f S) differ significantly between halophytic and glycophytic species (Saha et al. 2012). Glycophytic species decrease transpiration when salinity begins to reach levels that the plants cannot tolerate, while halophytic species can continue to transpire at relatively high salinity. To represent the differences in glycophytic and halophytic vegetation, we modified two functions for the salinity effect (f S) based on empirical data (Lin and Sternberg 1992; Sternberg et al. 2007).
where f S1 is the salinity effect for freshwater marsh, and f S2 is the salinity effect for mangroves. Both functions decrease with increasing S V, but f S1 declines much more sharply.
Vegetation Dynamics
Competition between mangrove and freshwater marsh (sawgrass: C. jamaicense) was simulated by extending an existing individual-based model, the Spatially Explicit Hammocks and Mangroves (SEHM) model (Jiang et al. 2012a). The SEHM model was developed for two competing halophytic and glycophytic tree types, based on a neighborhood competition model between individual plants from Berger and Hildenbrandt (2000) and self-reinforcement between of soil porewater salinity and vegetation. The same concepts are assumed to play a role in mangrove–marsh ecotones. Direct competition (i.e., for light) of vegetation would likely favor sawgrass over mangrove seedlings, though the reverse would occur once mangroves reached large size. But indirect effects on vadose zone salinity favor mangroves. Mangroves are able to utilize both shallow soil water and deeper groundwater. During the dry season, when the upper soil layer can become highly saline, mangroves have been observed to switch to less saline groundwater (Ewe et al. 2007). Sawgrass appears to be restricted to utilizing only the shallow water. Although sawgrass has some salinity tolerance, it will ultimately be outcompeted when mangrove density is high enough to raise dry season vadose zone salinity to high levels (Ewe et al. 2007). These opposing mechanisms of competition are hypothesized here to be sufficient to maintain a sharp ecotone, though other processes, such as fire, which can destroy mangrove seedlings, may contribute to the boundary (Egler 1952). Fire processes are implicitly considered to have a positive effect on marsh ecosystems that maintains open canopies, as with some models in savanna–forest systems (Staver and Levin 2012). We use the Overview, Design Concepts, and Details protocol (Grimm et al. 2006; Grimm et al. 2010) to describe the model in detail (see Supplementary Material). A simple overview of vegetation dynamics is described as follows.
We simulated competition between mangroves and freshwater marshes, including the effects of the abiotic factors of hydroperiod and rooting zone porewater. Freshwater marsh (e.g., Cladium) is relatively spatially homogeneous at a scale of 10 by 10 m and was simulated as biomass per unit area. The biomass of marsh in a given cell was determined by monthly gains from photosynthesis and losses due to respiration or mortality. Photosynthesis was modeled as the maximum possible rate multiplied by limitation factors, including salinity, light, and number of days of flooding. Mangrove biomass has indirect effects of limiting marsh spread both through influencing soil salinity and retarding fire percolation (analogous to a forest–savanna ecosystem; e.g., Staver and Levin 2012). Similar to SEHM (Jiang et al. 2012a), mangrove dynamics were modeled as individual-based. For each monthly time step, every tree has a growth increment that is a function of light, number of days of flooding, salinity of the particular spatial cell, and fire damage induced by biomass of marsh, which promotes the spread of fire. When a mangrove tree reaches maturity, new propagules are produced at monthly intervals by the tree and disperse to new locations according to a probability function. The probability of each propagule to produce a successful new recruit depends on the salinity of the soil porewater at the location the seedling reaches. At the end of the monthly time step, the probability of tree mortality is related to size-dependent factors, such as too small a diameter at breast height of the tree, or from reduced growth rate caused by competition or salinity.
Analysis and Simulation
Model simulations were performed on a 30 × 30 grid of 10-m square cells applied to the study site. Elevations of all the cells were assumed to be 8 cm above sea level (NAVD 88). Tidal signal inputs were a maximum at the riverward edge of the simulation landscape and decreased exponentially inland, based on empirical relationship from Krauss et al. (2009). On a daily time scale, the hydrodynamics submodel updated salinities of the soil porewater and groundwater. Monthly average values of salinity in each cell, which affect tree growth and propagule establishment, were then used in the vegetation dynamics submodels.
The model was applied to an actual hurricane that produced a storm surge at the study site. This application was designed to demonstrate the feasibility of the model. Because the size of the impact of the storm surge was small at the study site and because salinities at the Harney River study sites are usually less than 5 during the wet season, an effect on the mangrove–marsh ecotone was not expected. Hydrology data from January 2000 to January 2010 were analyzed to investigate the possibility of effects from storm surges during that period. Hurricane Wilma, a category-3 (Saffir–Simpson scale) hurricane, made landfall upon the western facing Florida Everglades mangrove coast during low tide on the morning of 24 October 2005. The eyewall of the hurricane crossed over Everglades City, about 50 km north of the Harney River. Hurricane-induced floodwater levels along the mouth and lower reach of the Harney River exceeded 3 m (Smith III et al. 2009); however, along the middle reach (9.5 km from the Gulf of Mexico) of the Harney River measured water levels increases were dampened to only 11.2 and 4.9 cm (NAVD 88) at USGS gauge SH4 and SH5, respectively. Both gauge stations were fully operational during the storm event. Salinity and water level changes after Hurricane Wilma were used as inputs to the simulation model in order to test the potential of the storm surge to produce a regime shift across some area of marsh vegetation.
To investigate how salinity intrusion and mangrove dispersal from larger hurricane events might trigger ecotone movement, we supplemented the simulation of Hurricane Wilma with simulations of vegetation dynamics under given scenarios of both different levels of salinity disturbance duration and different amounts of mangrove propagules carried by the storm surge into freshwater marsh sites. Due to lack of studies on mangrove propagule dispersal after the storm surge, we assumed pulse dispersal input to freshwater marsh sites with Poisson distribution on the spatial grid. Three levels of propagule density transported by the storm surge, 0, 1,000, and 2,000 propagules/ha, were tested. Long-term vegetation dynamics were also simulated under given scenarios of salinity disturbance duration caused by storm surge salinity intrusion. For each level of mangrove propagule density, three levels of salinity intrusion duration were assumed, 0 (no salinity overwash), 1, and 2 years. It was assumed that a level of salinity in the groundwater (i.e., freshwater lens, S GW) remained at a level of 35 for these durations.
Results
Analysis of 10-Year Time Series of Empirical Water Budget Data and Salinity
Precipitation data from 2000 to 2010 showed a seasonal pattern (Table 1), with high rainfall occurring in the wet season (May–November). Rainfall amounts on 24 October 2005, the date Hurricane Wilma made landfall, reach a monthly average of time period we investigated. Potential evapotranspiration for the study site was estimated by USGS and showed a seasonal pattern with a peak occurring between April and August (Table 1), corresponding to the solar cycle. Groundwater level data for SH4 (mangrove) and SH5 (marsh) showed a seasonal pattern, with highest levels around September and lowest around March of each year (Table 1). Groundwater level increased significantly on the day Wilma made landfall and the following few days. Groundwater level returned to its usual trajectory of seasonal variation soon after the storm surge.
The groundwater salinity patterns of SH4 and SH5 were different (Table 1). Groundwater salinity at SH4 increased during the dry season and declined when the relatively fresher Harney River high tides (diluted river salinities from rainfall and freshwater inflows) combined with precipitation to wash out salt embedded within the limestone and peat during wet season. However, groundwater salinity at SH5 was relatively stable due to its distance away from the riverbank (∼300 m). At SH5, groundwater salinity increased by a slight amount after Hurricane Wilma, whereas, groundwater salinity decreased at SH4. Surface water salinity at both sites doubled after Hurricane Wilma due to the storm surge that pushed water upstream from the Gulf of Mexico but was still lower than dry season salinity. We did not find any long-lasting salinity increase followed by the storm surge for either SH4 or SH5.
Simulations of Soil Pore Salinity
We compare simulations of soil porewater salinity to surface water, due to lack of field data of soil porewater salinity during that time period. Soil porewater salinity is close to surface water salinity during wet season but may differ during dry season with evaporation of surface water. Simulations of the soil porewater salinity captured the seasonal signal pattern of surface water from SH4, which is dominated mostly by mangroves (Fig. 2). While model freshwater marsh-dominated cells show a dampened seasonal pattern similar to mangrove sites, the model does not fit SH5 surface water data very well (Fig. 2). Soil porewater salinity from mangrove-dominated cells increased dramatically in the model during the late dry season due to strong evapotranspiration and little precipitation. Overall salinities of the freshwater marsh sites were lower than mangrove sites, especially during dry season. This occurs because evapotranspiration in the interior freshwater marsh slows down if soil porewater salinity starts to increase, thus dampening the infiltration of underlying saline water.

Simulation of soil porewater salinities averaging mangrove sites and marsh sites, respectively, compared with monthly average with standard error of surface water salinities from SH4 and SH5, respectively
Simulation of Vegetation Dynamics
The 10-year average hydrology data were first used in the simulation model to determine whether the sharp vegetation ecotone observed at SH5 would be produced. The vegetation distribution that is produced shows a clear boundary between mangroves and freshwater marsh, resembling field observations of the coastal Everglades (Fig. 3). Then we simulated the effect of the hurricane. As shown in Table 1, Hurricane Wilma did not cause significant changes in groundwater salinity. The water level increased for several days but then decreased again. Our simulations, which included pulse increases of water level and surface water salinity, without assuming any direct mortality to the original vegetation or input of mangrove seedlings due to the storm surge, did not show any long-term effects on freshwater marsh dynamics from Hurricane Wilma stemming from salinity intrusion. In this simulation, it was assumed that no wind damage occurred from the hurricane, which could have contributed to a regime shift. It has been reported that direct hurricane wind damage to the mangrove forest can be 30–80 %, depending on the intensity of hurricane and the distance from the eye of the hurricane (Armentano et al. 1995; Harcombe et al. 2009; Milbrandt et al. 2006). We repeated the simulation with assumptions of non-zero storm damage. Figure 4 shows the recovery of the basal area of mangroves projected by the model following 30 %, 50 %, and 80 % mortality, respectively, due to hurricane damage. The mangrove forest recovered at different rates, without any expansion of freshwater marsh into the mangrove zone.

Spatial distributions of individual mangroves (dark circles) and freshwater marsh (gray cells) output from simulation model. Area of circles represents canopy area of individual mangrove tree

Model-projected basal areas of mangroves following 30 %, 50 %, and 80 % destruction by hurricane, respectively, at year 80
Effects of Salinity Intrusion and Mangrove Dispersal
As mentioned in the “Methods,” because the climatic data from 2000 to 2010 produced no situations in which a regime shift was likely, we developed several artificial scenarios for simulation. We combined three levels of salinity intrusion duration and three levels of mangrove propagule density passively dispersed to freshwater marsh sites to further analyze storm surge effects on ecotone dynamics. Results show that, in the case of salinity intrusion being rapidly washed away after hurricane, the ecotone between mangroves and freshwater marsh is, as might be expected, relatively stable (Fig. 5 left panels). As salinity intrusion duration increased, however, the model predicted more marsh habitats changing to mangrove permanently. The results are consistent with Jiang et al. (2012b), in which a mathematical model showed that only a relatively long “press” disturbance of salinity for a long period of time might cause regime shift. In these cases, passive transport of mangrove propagules by a storm surge could facilitate a vegetation regime shift triggered by salinity intrusion (Fig. 5, middle and right panels).

Spatial distribution between freshwater marsh (gray cells) and individual mangrove trees (black circles) under nine different scenarios, which are combinations of three levels of mangrove seedlings transported into the marsh (none transported, a moderate number transported with 1,000 propagules/ha, and a large number transported with 2,000 propagules/ha) and three levels of salinity intrusion duration (no saltwater retention, 1-year salinity retention, and 2-year salinity retention)
Discussion
Earlier studies have reviewed the effects of large storms on coastal ecosystems (e.g., Craighead and Gilbert 1962; Craighead 1971; Ellison and Stoddart 1991; Roth 1992; Ellison 1993; Smith III et al. 1994; Field 1995; Michener et al. 1997; Paerl et al. 2001; Doyle et al. 2003; McLeod and Salm 2006; Gilman et al. 2006, 2007; Smith III et al.. 2009) or modeled possible effects of storms on mangrove ecosystems (e.g., Doyle et al. 2003). Our modeling approach differs from previous models by focusing on the dynamics of the ecotone between coastal halophytic and glycophytic vegetation (mangrove and freshwater marsh) and by modeling the ecotone as being maintained by self-reinforcing feedbacks between the biotic and abiotic environment. The hypothesis that halophytic and glycophytic vegetation represent alternative stable states over areas of coastal landscape implies that the ecotone is susceptible to regime shifts. Our model can be used to estimate the resilience of the ecotone to realistic effects of storm surge events in southern Florida. We explicitly investigated different levels of two significant factors, salinity intrusion and the transport of mangrove propagules, associated with the storm surge. These two factors are prerequisites for triggering vegetation changes, as both an increase in salinity and sufficient mangrove propagules are needed to create the positive feedback between plants and soil porewater salinity necessary to cause the regime shift. We observed that the spatial extent of the regime shift responded sensitively to the duration of salinity intrusion, it is just that increased salinity concentrations did not persist long enough after Hurricane Wilma to drive this change. It is uncertain whether other types of storms might have a different influence on ecotonal movement, as depicted theoretically with only a 2-year post-storm retention of salinity (Fig. 5). The ecotone was resilient to passive mangrove propagule transport if the increase in salinity was washed away by precipitation quickly after the storm surge.
Hurricane Wilma was not predicted to cause significant hydrology changes at the marsh–mangrove ecotone study site SH5 for several reasons. First, SH5 is 9.5 km upstream from the Gulf of Mexico on the Harney River, so the storm surge was greatly dampened at that point. Intrusion of water from Harney River by Hurricane Wilma was much fresher (∼5) than marine (∼35) water and even less saline than groundwater at SH4. In addition, saline water intrusion was reduced prior to Wilma from heavy rainfall during wet season. Furthermore, the slightly lower elevation of SH5 helped in the accumulation of freshwater from upstream during wet season. Other studies of hurricane effects have found similar brief durations of salinization and thus no long-term effects on vegetation (e.g., Chabreck and Palmisano 1973; Conner et al. 1989; Flynn et al. 1995) but have noted that longer durations may cause changes in plant communities (McKee and Mendelssohn 1989; Howard and Mendelssohn 2000). Longer durations of salinity can occur when a storm surge is followed by a prolonged dry period, especially in areas where saline water is trapped in swales (Hook et al. 1991) or behind berms (Pearlstine et al. 2010). For such reasons, hurricanes might cause more severe changes in hydrology at other locations, e.g., the Mississippi Delta from Hurricanes Katrina and Rita (Day Jr et al. 2007; Schriever et al. 2009). The “lower Saline Everglades” is an area where regime shifts might be expected. Egler (1952) noted that a succession of storm tides could lead to a buildup of salinity on the flat, poorly drained terrain, as well as “bring in vast numbers of Rhizophora fruits, which root and grow rapidly.” The transition of that area from a mixture of C. jamaicense and R. mangle to pure R. mangle noted by Ross et al. (2000) 40 years later could have occurred as such a shift.
Although sea level rise and storm surges have been considered to be primary drivers of coastal vegetation changes, the mechanisms by which these two factors trigger vegetation changes are different. On the one hand, sea level rise is a gradual change that can eventually push environmental conditions (e.g., groundwater salinity) past a threshold, beyond which an affected vegetation zone could shift from glycophytic to halophytic vegetation. On the other hand, a regime shift caused by storm surge is a large disturbance that pushes the system beyond the threshold all at once, such that it cannot return to the original vegetation state (glycophytic) but moves to the alternative state (halophytic). If the disturbance is not so large that it pushes the system outside its domain of ecological resilience, the ecosystem can return to its original state following a disturbance. Evapotranspiration is the process causing the largest water output from the Everglades ecosystem (Saha et al. 2012), and it tends to increase soil salinity, as salt is left in the soil. Plants can adjust their transpiration in response to changing soil salinity conditions and, therefore, modify local salinity conditions. If an external salinity disturbance lasts for a long time, mangroves can invade freshwater marsh habitats and then maintain high salinity via evapotranspiration. The passive transports of mangrove propagules then overcome the mangrove dispersal limitation on the upper inland. Although there are rare reports of mangrove seedling dispersal after a storm surge (Rathcke and Landry 2003), multiple experiments on mangrove dispersal indicate that pulse dispersal and retention after storm surge is possible (Rabinowitz 1978a, b; Sousa et al. 2007; Peterson and Bell 2012; Van der Stocken et al. 2013). Perhaps this research may encourage empirical studies to investigate mangrove dispersal and establishment after a storm surge.
References
Agnew, A.D.Q., J.B. Wilson, and M.T. Sykes. 1993. A vegetation switch as the cause of a forest/mire ecotone in New Zealand. Journal of Vegetation Science 2: 273–278.
Armentano, T. V., Doren, R. F., Platt, W. J., Mullins, T. (1995). Effects of hurricane Andrew on coastal and interior forests of Southern Florida: overview and synthesis. Journal of Coastal Research, (21), 111–144.
Baldwin, A.H., and I.A. Mendelssohn. 1998. Effects of salinity and water level on coastal marshes; an experimental test of disturbance as a catalyst for vegetation change. Aquatic Botany 61: 255–268.
Ball, M.C. 1980. Patterns of secondary succession in a mangrove forest of Southern Florida. Oecologia 44(2): 226–235.
Beisner, B.E., D.T. Haydon, and K. Cuddington. 2003. Alternative stable states in ecology. Frontiers in Ecology and the Environment 1: 376–382.
Berger, U., and H. Hildenbrandt. 2000. A new approach to spatially explicit modelling of forest dynamics: spacing, ageing and neighbourhood competition of mangrove trees. Ecological Modelling 132: 287–302.
Bertness, M.D. 2007. Atlantic shorelines: natural history and ecology. Woodstock, UK: Princeton University Press.
Chabreck, R.H., and A.W. Palmisano. 1973. The effects of Hurricane Camille on the marshes of the Mississippi River Delta. Ecology 54: 1118–1123.
Conner, W.H., J.W. Day Jr., R.H. Baumann, and J.M. Randall. 1989. Influence of hurricanes on coastal ecosystems along the northern Gulf of Mexico. Wetlands Ecology and Management 1: 45–56.
Craighead, F.C. 1971. The Trees of South Florida., Vol. 1: The Natural Environments and Their Succession. Coral Gables, Florida: University of Miami Press.
Craighead, F.C., and V.C. Gilbert. 1962. The effects of Hurricane Donna on the vegetation of southern Florida. Quarterly Journal of the Florida Academy of Sciences 25: 1–28.
da Ewe, S.M.L., L.S.L. Sternberg, and D.L. Childers. 2007. Seasonal water uptake patterns in the saline southeast Everglades ecotone. Oecologia 152: 607–616.
Day Jr., J.W., D.F. Boesch, E.J. Clairain, G.P. Kemp, S.B. Laska, W.J. Mitsch, K. Orth, H. Mashriqui, D.J. Reed, L. Shabman, C.A. Simenstad, B.J. Streever, R.R. Twilley, C.C. Watson, J.T. Wells, and D.F. Whigham. 2007. Restoration of the Mississippi Delta: lessons from hurricanes Katrina and Rita. Science 315(5819): 1679–1684.
Doyle, T.W., Girod, G.F. and Books, M.A. 2003. Chapter 12: modeling mangrove forest mitigation along the southwest coast of Florida under climate change. In: Integrated Assessment of the Climate Change Impacts on the Gulf Coast. Ning, Z.H., Turner, R.E., Doyle, T. and Abdollahi, K.K. (lead authors). Region. Gulf Coast Climate Change Assessment Council (GCRCC) and Louisiana State University (LSU) Graphic Services, pp 211–221.
Egler, F.E. 1952. Southeast saline Everglades vegetation, Florida, and its management. Vegetatio 3: 213–265.
Ellison, J.C. 1993. Mangrove retreat with rising sea level, Bermuda. Estuarine, Coastal and Shelf Science 37: 75–87.
Ellison, J.C., and D.R. Stoddart. 1991. Mangrove ecosystem collapse during predicted sea-level rise: Holocene analogues and implications. Journal of Coastal Research 7: 151–165.
Field, C.D. 1995. Impact of expected climate change on mangroves. Hydrobiologia 295: 75–81.
Flynn, K.M., K.L. McKee, and I.A. Mendelssohn. 1995. Recovery of freshwater marsh vegetation after a saltwater intrusion event. Oecologia 103: 63–72.
Folke, C., S. Carpenter, B. Walker, M. Scheffer, T. Elmquist, L. Gunderson, and C.S. Holling. 2004. Regime shifts, resilience, and biodiversity in ecosystem management. Annual Reviews of Ecology and Evolutionary Systematics 35: 557–581.
Gaiser, E.E., A. Zafiris, P.L. Ruiz, F.A.C. Tobias, and M.S. Ross. 2006. Tracking rates of ecotone migration due to salt-water encroachment using fossil mollusks in coastal South Florida. Hydrobiologia 569: 237–257.
Gilman, E.L., J. Ellison, V. Jungblut, H. Van Lavieren, L. Wilson, F. Areki, G. Brighouse, J. Bungitak, E. Dus, M. Henry, M. Kilman, E. Matthews, I. Sauni Jr., N. Teariki-Ruatu, S. Tukia, and K. Yuknavage. 2006. Adapting to Pacific Island mangrove responses to sea level rise and climate change. Climate Research 32: 161–176.
Gilman, E.L., J. Ellison, and R. Coleman. 2007. Assessment of mangrove response to projected relative sea-level rise and recent historical reconstruction of shoreline position. Environmental Monitoring and Assessment 124(1–3): 105–30.
Grimm, V., U. Berger, F. Bastiansen, S. Eliassen, V. Ginot, J. Giske, J. Goss-Custard, T. Grand, S.K. Heinz, G. Huse, A. Huth, J.U. Jepsen, C. Jorgensen, W.M. Mooij, B. Muller, G. Pe’er, C. Piou, S.F. Railsback, A.M. Robbins, M.M. Robbins, E. Rossmanith, N. Ruger, E. Strand, S. Souissi, R.A. Stillman, R. Vabo, U. Visser, and D.L. DeAngelis. 2006. A standard protocol for describing individual-based and agent-based models. Ecological Modelling 198(1–2): 115–126.
Grimm, V., U. Berger, D.L. DeAngelis, J.G. Polhill, J. Giske, and S.F. Railsback. 2010. The ODD protocol: a review and first update. Ecological Modelling 221(23): 2760–2768.
Harcombe, P.A., L.E. Mann Leipzig, and I.S. Elsik. 2009. Effects of hurricane Rita on three long-term forest study plots in east Texas, USA. Wetlands 29(1): 88–100.
Hook, D.D., M.A. Buford, and T.M. Williams. 1991. Impact of Hurricane Hugo on the South Carolina coastal plain forest. Journal of Coastal Research 8: 291–300.
Howard, R.J., and I.A. Mendelssohn. 2000. Structure and composition of oligohaline marsh plant communities exposed to salinity pulses. Aquatic Botany 68: 143–164.
Jiang, J., D. DeAngelis, T. Smith, S. Teh, and H.-L. Koh. 2012a. Spatial pattern formation of coastal vegetation in response to external gradients and positive feedbacks affecting soil porewater salinity: a model study. Landscape Ecology 27(1): 109–119.
Jiang, J., D. Gao, and D.L. DeAngelis. 2012b. Towards a theory of ecotone resilience: coastal vegetation on a salinity gradient. Theoretical Population Biology 82: 27–37.
Kirwan, M., and A. Murray. 2007. A coupled geomorphic and ecological model of tidal marsh evolution. Proceedings of the National Academy of Sciences 104: 6118–6122.
Krauss, K., T.W. Doyle, T.J. Doyle, C.M. Swarzenski, A.S. From, R.H. Day, and W.H. Conner. 2009. Water level observations in mangrove swamps during tow hurricanes in Florida. Wetlands 29(1): 142–149.
Krauss, K.W., A.S. From, T.W. Doyle, T.J. Doyle, and M.J. Barry. 2011. Sea-level rise and landscape change influence mangrove encroachment onto marsh in the Ten Thousand Islands region of Florida, USA. Journal of Coastal Conservation 15(4): 629–638.
Lin, G., and L.S.L. Sternberg. 1992. Effect of growth form, salinity, nutrient, and sulfide on photosynthesis, carbon isotope discrimination and growth of red mangrove (Rhizophora mangle L.). Aust. J. Plant Physiology 19: 509–517.
Lugo, A.E., and S.C. Snedaker. 1974. The ecology of mangroves. Annual Review of Ecology and Systematics 5: 39–64.
Maul, G.A., and D.M. Martin. 1993. Sea-level rise at Key-West, Florida, 1846–1992—America longest instrument record. Geophysical Research Letters 20(18): 1955–1958.
McKee, K.I., and I.A. Mendelssohn. 1989. Response of a freshwater marsh plant community to increased salinity and increased water level. Aquatic Botany 34: 801–816.
McLeod, E., and R.V. Salm. 2006. Managing Mangroves for Resilience to Climate Change. Switzerland: The World Conservation Union (IUCN), Gland. 64 pp.
Michener, W.K., E.R. Blood, K.L. Bildstein, M.M. Brinson, and L.R. Gardner. 1997. Climate change, hurricanes and tropical storms, and rising sea level in coastal wetlands. Ecological Applications 7(3): 770–801.
Milbrandt, E.C., J.M. Greenawalt-Boswell, P.D. Sokoloff, and S.A. Bortone. 2006. Impact and response of southwest Florida mangroves to the 2004 hurricane season. Estuaries and Coasts. 29(6): 979–984.
Paerl, H.W., J. Bales, L. Ausley, C. Buzzelli, L. Crowder, L. Eby, J. Fear, M. Go, B. Peirls, T. Richardson, and J. Ramus. 2001. Ecosystem impacts of three sequential hurricanes (Dennis, Floyd, and Irene0 on the United States’ largest lagoonal estuary, Pamlico Sound, NC. Proceedings of the National Academy of Sciences 98(10): 5655–5660.
Pearlstine, L.G., E.V. Pearlstine, and N.G. Aumen. 2010. A review of the ecological consequences and management implications of climate change for the Everglades. Journal of the North American Benthological Society 29: 1510–1526.
Person, B.T., and R.W. Ruess. 2003. Stability of a subarctic saltmarsh: plant community resistance to tidal inundation. Ecoscience 10(3): 351–360.
Peterson, J.M., and S.S. Bell. 2012. Tidal events and salt-marsh structure influence black mangrove (Avicennia germinans) recruitment across and ecotone. Ecology 93: 1648–1658.
Raabe, E.A., L.C. Roy, and C.C. McIvor. 2012. Tampa Bay coastal wetlands: nineteenth to twentieth century tidal marsh-to-mangrove conversion. Estuaries and Coasts 35: 1145–1162.
Rabinowitz, D. 1978a. Dispersal properties of mangrove propagules. Biotropica 10(1): 47–57.
Rabinowitz, D. 1978b. Early growth of mangrove seedlings in Panama, and a hypothesis concerning relationship of dispersal and zonation. Journal of Biogeography 5(2): 113–133.
Rathcke, B.J., and C.L. Landry. 2003. Dispersal and recruitment of white mangrove on San Salvador Island, Bahamas after Hurricane Floyd. In Proceedings of the ninth symposium on the natural history of the Bahamas, ed. D.L. Smith and S. Smith. Bahamas: Gerace Research Center, San Salvador.
Ross, M.S., J.F. Meeder, J.P. Sah, P.L. Ruiz, and G.J. Telesnicki. 2000. The Southeast Saline Everglades revisited: 50 years of coastal vegetation change. Journal of Vegetation Science 11: 101–112.
Ross, M.S., J.J. O’Brien, R.G. Ford, K.Q. Zhang, and A. Morkill. 2009. Disturbance and the rising tide: the challenge of biodiversity management on low-island ecosystems. Frontiers in Ecology and the Environment 7(9): 471–478.
Roth, L.C. 1992. Hurricanes and mangrove regeneration: effects of Hurricane Joan, October 1988, on the vegetation of Isla del Venado, Bluefields, Nicaragua. Biotropica 24: 375–384.
Saha, A., S. Saha, J. Sadle, J. Jiang, M. Ross, R. Price, L. Sternberg, and K. Wendelberger. 2011. Sea level rise and South Florida coastal forests. Climatic Change 107(1): 81–108.
Saha, A., C. Moses, R. Price, V. Engel, T. Smith III, and G. Anderson. 2012. A hydrological budget (2002–2008) for a large subtropical wetland ecosystem indicates marine groundwater discharge accompanies diminished freshwater flow. Estuaries and Coasts. 35(2): 459–474.
Scheffer, M. 1997. The Ecology of Shallow Lakes. London: Chapman and Hall.
Schriever, T.A., J. Ramspott, B.I. Crother, and C.L. Fontenot. 2009. Effects of Hurricane Ivan, Katrina, and Rita on a southeastern Louisiana herpetofauna. Wetlands 29(1): 112–122.
Smith III, T.J., M. Robblee, H. Wanless, and T. Doyle. 1994. Mangroves, hurricanes, and lightning strikes. BioScience 44(4): 256–262.
Smith III, T.J., G.H. Anderson, K. Balentine, G. Tiling, G.A. Ward, and K.R.T. Whelan. 2009. Cumulative impacts of hurricanes on Florida mangrove ecosystems: sediment deposition, storm surges, and vegetation. Wetlands 29: 24–34.
Smith III, T.J., A. Foster, G. Tiling-Range, and J.W. Jones. 2013. Dynamics of mangrove–marsh ecotones in subtropical coastal wetlands: fire, sea-level rise, and water levels. Fire Ecology 9(1): 66–77.
Sousa, W.P., P.G. Kennedy, B.J. Mitchell, and B.M. Ordonez. 2007. Supply-side ecology in mangroves: do propagule dispersal and seedling establishment explain forest structure? Ecological Monographs 77(1): 53–76.
Staver, A.C., and S.A. Levin. 2012. Integrating theoretical climate and fire effects on savanna and forest systems. The American Naturalist 180(2): 211–224.
Sternberg, L.D.L., S.Y. Teh, S.M.L. Ewe, F. Miralle-Wilhelm, and D.L. DeAngelis. 2007. Competition between hardwood hammocks and mangroves. Ecosystem. 10: 648–660.
Steyer, G. D., Cretini, K. F., Piazza, S., Sharp, L. A., Snedden, G. A., Sapkota, S. 2010. Hurricane influences on vegetation community change in coastal Louisiana. Open-File Report 2010–1105. U. S. Department of the Interior, U. S. Geological Survey.
Teh, S.Y., D.L. DeAngelis, L.D.L. Sternberg, F.R. Miralles-Wilhelm, T.J. Smith, and H.L. Koh. 2008. A simulation model for projecting changes in salinity concentrations and species dominance in the coastal margin habitats of the Everglades. Ecological Modelling 213(2): 245–256.
Temmerman, S., T.J. Bouma, G. Govers, Z. Wang, M. De Vries, and P.M. Herman. 2005. Impact of vegetation on flow routing and sedimentation patterns: three-dimensional modeling for a tidal marsh. Journal of Geophysical Research 110, F04019. doi:10.1029/2005JF000301.
Temmerman, S., T.J. Bouma, J. van de Koppel, D. van der Wal, M. De Vries, and P.M. Herman. 2007. Vegetation causes channel erosion in a tidal landscape. Geology 35: 631–634.
Van der Stocken, T., D.J.R. De Ryck, T. Balke, T.J. Bouma, F. Dahdouh-Guebas, and N. Koedam. 2013. The role of wind in hydrochorous mangrove propagule dispersal. Biogeosciences Discuss. 10: 895–925.
White, I., and T. Falkland. 2010. Management of freshwater lenses on small Pacific islands. Hydrogeology Journal 18(1): 227–246.
Zhang, K., H. Liu, Y. Li, H. Xu, J. Shen, J. Rhome, and T.J. Smith III. 2012. The role of mangroves in attenuating storm surges. Estuarine, Coastal and Shelf Science 102–103: 11–23.
Acknowledgments
We appreciate the thorough and helpful reviews of two reviewers for the journal and one for USGS peer review. JJ was supported as Postdoctoral Fellow at the National Institute for Mathematical and Biological Synthesis, an Institute sponsored by the National Science Foundation, the U.S. Department of Homeland Security, and the U.S. Department of Agriculture through NSF Award #EF-0832858, with additional support from The University of Tennessee, Knoxville. This research was partially supported the USGS project “A land of flowers on a latitude of deserts” funded by the National Climate Change and Wildlife Science Center and supported by the FISCHS Project (Future Impacts of Sea Level Rise on Coastal Habitats and Species) at the USGS Southeast Ecological Science Center, funded by USGS Ecosystems Mapping and the USGS Greater Everglades Priority Ecosystems Science. This material was developed in collaboration with the Florida Coastal Everglades Long-Term Ecological Research program under National Science Foundation Grant No. DBI-0620409.
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 349 kb)
Rights and permissions
About this article
Cite this article
Jiang, J., DeAngelis, D.L., Anderson, G.H. et al. Analysis and Simulation of Propagule Dispersal and Salinity Intrusion from Storm Surge on the Movement of a Marsh–Mangrove Ecotone in South Florida. Estuaries and Coasts 37, 24–35 (2014). https://doi.org/10.1007/s12237-013-9666-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-013-9666-4