
Abstract
In this paper, we provide screening-level analysis of plausible Everglades ecosystem response by 2060 to sea level rise (0.50 m) interacting with macroclimate change (1.5 °C warming, 7% increase in evapotranspiration, and rainfall that either increases or decreases by 10%). We used these climate scenarios as input to the Ecological Landscape Model to simulate changes to seven interactive hydro-ecological metrics. Mangrove forest and other marine influences migrated up to 15 km inland in both scenarios, delineated by the saltwater front. Freshwater habitat area decreased by 25–30% under our two climate change scenarios and was largely replaced by mangroves and, in the increased rainfall scenario, open water as well. Significant mangroves drowned along northern Florida Bay in both climate change scenarios due to sea level rise. Increased rainfall of 10% provided significant benefits to the spatial and temporal salinity regime within the marine-influenced zone, providing a more gradual and natural adjustment for at-risk flora and fauna. However, increased rainfall also increased the risk of open water, due to water depths that inhibited mangrove establishment and reduced peat accumulation rates. We infer that ecological effects related to sea level rise may occur in the extreme front-edge of saltwater intrusion, that topography will control the incursion of this zone as sea level rises, and that differences in freshwater availability will have ecologically significant effects on ecosystem resilience through the temporal and spatial pattern of salinity changes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The fate of the Everglades is of global concern. In 2000, Congress authorized a unique federal/state partnership to guide the largest hydrologic restoration project ever undertaken in the United States, with an estimated price-tag of $10.5 billion. The Comprehensive Everglades Restoration Plan (CERP) was developed “for the purpose of restoring, preserving, and protecting the South Florida ecosystem” (USACE 1999). Key goals were to “provide for the protection of water quality in, and the reduction of the loss of fresh water from, the Everglades” (USACE 1999). A timeline of 35+ years was set, and several projects have already been completed.
In recent years there has been a growing recognition that macroclimate change (for example, changes in temperature and precipitation) will be a major driver of coastal wetland transformations this century, and these factors can no longer be overlooked in assessing coastal wetland vulnerability (Gabler et al. 2017; Osland et al. 2016). In its 5th Biennial Review “Progress toward restoring the Everglades,” the National Research Council (NRC) expressed grave concern that future climate threats to the Everglades were not being considered beyond mere submergence in the Comprehensive Everglades Restoration Plan (NRC 2014b). The report called for scenarios-based modeling that provides indications of the sensitivity of the South Florida Ecosystem to temperature and precipitation variability, to better understand possible climate impacts.
In this paper, we take an important step forward in addressing this need, presenting the first landscape-scale modeling of Everglades ecosystem response to plausible scenarios of temperature, rainfall, and sea level rise. Scenarios-based modeling is a primary tool for decision-making under uncertainty, as a means of what-if analysis rather than prediction (Moss et al. 2010). A vision of plausible outcomes can inform strategies to build resilience and robustness into restoration efforts as we look ahead to the middle of the 21st century. This project has significance beyond the Everglades, providing a vision of potential future changes that has implications for climatically similar regions globally, because coastal wetland ecosystems are functionally similar worldwide (Gabler et al. 2017).
Our objective was to provide screening-level analysis of two basic questions central to Everglades restoration in the face of climate change. First, what type of ecological responses may occur in the southern Everglades under a “mid-range” estimate of future SLR, beyond mere inundation? Second, how may changes in macroclimate (air temperature, evapotranspiration, and rainfall) interact with sea level rise to alter the vulnerability or resilience of this iconic coastal wetland?
Materials and Methods
Study Area
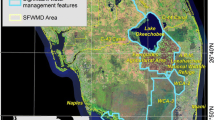
To better inform restoration efforts we focused this project on the Everglades National Park (ENP) which is in the southern portion of the Everglades Landscape Model (ELM) domain (Fig. 1). The Everglades Region of South Florida, USA, includes neotropical estuaries, wetlands, and uplands with agricultural and urban land uses in close proximity. The topography across the Everglades landscape is extremely flat, with an elevation gradient of just 5 cm km−1 from its headwaters to Florida Bay. The Everglades is ombrotrophic, relying primarily on direct rainfall and rain-derived inflow from basins to the north. Like other marl and peat ecosystems, the Everglades’ ecological structure and function require that water availability exceeds evapotranspiration (Mitsch and Gosselink 2000; Nungesser et al. 2015). Although rainfall is highly variable, historical annual precipitation in the Everglades (132–152 cm) surpassed annual evapotranspiration by 8–20 cm year−1 (Fernald and Purdum 1998; Nungesser et al. 2015).

The South Florida Water Management Model (SFWMM) domain, the regional domain of the Everglades Landscape Model (ELM), and the Everglades National Park (ENP) boundary in South Florida
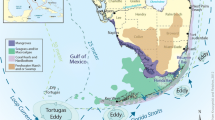
The ENP has two main flow pathways from upstream sources. The larger is Shark River Slough; it angles to the southwest and drains into the Gulf of Mexico (Fig. 2). To the east is the smaller Taylor Slough, which flows southward into the northeast corner of Florida Bay. The ENP includes a mosaic of habitats including marl prairie, cypress domes, hardwood hammocks and pinelands, and a unique “ridge and slough” habitat (discussed in more detail in a subsequent section).

The Everglades National Park, including the location of select FCE LTER monitoring station sites shown along a freshwater-estuary-marine transect in Shark River Slough (actual site names have the prefix “SRS,” omitted here for brevity) and the other in Taylor Slough (where actual site names have the prefix “TS/Ph”); the approximate boundaries of these two main flow-ways are delineated with dotted white curves. (Note: FCE LTER has monitoring stations at 17 sites, but this project focuses only on those along the two transects, as displayed here)
To better understand the hydro-ecology of the important transition from freshwater through estuaries to marine waters, the Florida Coastal Everglades Long Term Ecological Research Project (FCE LTER) set up long term monitoring stations along Shark River Slough and Taylor Slough (locations provided in Fig. 2). Our project draws from long term data sets from these monitoring stations, and in turn contributes to the FCE LTER goal of understanding the ways in which hydrology, nutrients, climate, and human activities affect patterns in the Florida Coastal Everglades. Most areas in the exceptionally flat coastal zone lie below 0.40 m. An inland depression very close to mean sea level runs in an arc roughly parallel to the curving coastline, with Whitewater Bay occupying its deepest part (Fig. 2).
Scenarios
The optimal number of scenarios is generally considered to be three or four (Peterson et al. 2003). We determined that comparing a baseline (no climate change) scenario with two clearly distinct scenarios of plausible future climate change would provide the most clear, useful and visible alternative futures for restoration planning (Table 1). The mid-21st century (2050–2060) is optimal because Everglades restoration commonly use 50 years as the planning horizon (Obeysekera et al. 2015).
Our Baseline scenario projects how the system may respond over a four decade future period under current conditions (Obeysekera et al. 2015). Based on a synthesis of downscaled data, a set of plausible climate scenarios were developed (Obeysekera et al. 2015; Obeysekera et al. 2011). The key climate uncertainty is whether rainfall will increase or decrease, and by how much (Obeysekera et al. 2011). We chose two climate change scenarios that represent a useful contrast within the range of plausible-rainfall outcomes: a 10% decrease or increase, which we refer to as “−RF” and “+RF”, respectively, and collectively as our two “climate change scenarios.” While our two climate change scenarios contrast in rainfall, they both include the same warming (1.5 °C), increase in evapotranspiration (7%), and SLR (0.5 m). These projections fall within the conservative end of the recent assessments of temperature, precipitation, and SLR (Carter et al. 2014; Melillo et al. 2014; SFRCCC 2015). Our simulations provide an opportunity to evaluate the sensitivity of the ecosystem to alternative rainfall outcomes in the context of similar warming and SLR. Our climate change scenarios simulate several decades of altered climate and sea level dynamics in the absence of changes to existing water management practices or hydrologic restoration.
Water Level Distributions and Inflows
The climate scenarios from Table 1 were in turn used by Obeysekera et al. (2015) to drive the South Florida Water Management Model (SFWMM). This is a hydrologic model designed to simulate complex regional water management leading to altered water level distributions and flow throughout southern Florida (Fig. 1) (Obeysekera et al. 2015; SFWMD 2005). The SFWMM simulates rule-based water management and the subsequent distribution of water levels and daily flows through water control structures for the South Florida urban, agricultural and natural systems from Lake Okeechobee to the southern Everglades (Tarboton et al. 1999). An overview of the SFWMM can be found in Online Resource 1 in the supplemental material provided with this paper.
Construction of the Baseline scenario and all climate change scenarios assumes current (ca. 2012) water management infrastructure and operations (i.e., no CERP or other restoration projects). Model input for the Baseline scenario used 1965–2000 climate data. To construct the model input for the climate change scenarios, the Baseline scenario was modified with the appropriate climate and sea level changes. It is important to note that the full 0.50 m sea level hike was implemented on day 1 of the simulation, rather than as a gradual increase. Although not realistic in rate, this was taken as a necessary simplifying assumption, and considered to be a useful if imperfect proxy for the much more gradual, year-to-year SLR that is projected to occur over the next half century.
The resulting climate-scenario-based flows and water level distributions led to important assessments as to possible climate effects on a range of south Florida concerns such as water supply, soil accretion, restoration planning, wildlife populations and vegetation patterns over the next 50 years (Aumen et al. 2015; Catano et al. 2015; Havens and Steinman 2015; Nungesser et al. 2015; Obeysekera et al. 2015; Orem et al. 2015; van der Valk et al. 2015).
Because the results from this prior hydrologic modeling effort are used as boundary conditions for our model, some key water availability outcomes are important to note. First, the +RF scenario led to proportional changes to managed water control structure inflows and outflows in both of those basins, with flows to the ENP increasing approximately 35%. Second, the −RF scenario was apparently water-limited, and managed inflows into ENP were approximately halved relative to the Baseline simulation. Thus, through water management decisions embedded within the SFWMM, changes in rainfall result in indirect changes in managed inflow to the ENP.
Everglades Landscape Model (ELM)
In this paper, we take three scenarios (Table 1) and water level distributions from prior modeling efforts (described above) a step further by using them as input to an integrated hydro-ecological landscape model. The ELM is a dynamic regional-scale integrated model that simulates how changes in temperature, precipitation, and sea level may alter a complex, living system.
Wetland responses to macroclimate and SLR stressors depend upon feedbacks among hydrology, water quality, soil processes, plant communities, and human choices (Kirwan and Megonigal 2013). The integrated hydro-ecological dynamics of the ELM incorporate dynamic feedbacks among hydrology, nutrients, soils, periphyton, vegetation, and habitat succession. The ELM has been reviewed and accepted for formal CERP applications by an independent panel (Mitsch et al. 2007) and its calibration and validation are detailed in Fitz and Paudel (2012) and Fitz and Trimble (2006). The ELM has successfully been used to simulate feedbacks among ecosystem processes and evaluate important questions related to restoration alternatives (Fitz et al. 2004; Fitz et al. 2011; Fitz and Sklar 1999; Orem et al. 2014; Osborne et al. 2017 (in press)). For this project, we used the regional (10,394 km2) ELM v2.9 application at 0.25 km2 grid resolution, and a multidecadal time domain. Detailed information on methods used to calibrate and validate the model, as well as algorithms and data updates can be found online (www.ecolandmod.com). For performance assessments we used the long term monitoring data sets of extensive water quality measurements provided by the FCE LTER for Shark River Slough (Gaiser and Childers 2016) and Taylor Slough (Troxler 2017a; Troxler 2017b; Troxler and Childers 2008).
To understand the comparative scenario differences in landscape dynamics, we used seven fundamental ELM-simulated hydro-ecological metrics or performance measures: (1) surface water salinity (in practical salinity units), (2) surface water depth, cm, (3) peat accumulation rate, mm/year, (4) surface water velocity, m/d, (5) phosphorus concentration in surface water, μg/L, (6) phosphorus accumulation in soil, mg P/kg soil, and (7) habitat distribution change among mangrove forests, freshwater wetland habitats, and open water.
The quantitative calibration for the ELM peat accretion module relies largely on northern Everglades data (Fitz and Trimble 2006), due to the paucity of historical observations for peat accretion rate in most of the Everglades. Applying this module to our study area introduces distinct limitations for three different peat zones in our study area. First, in the freshwater remnant, we recognize that the peat accretion rate is variable and uncertain for the southern Everglades. The peat accretion component of the ELM integrates feedbacks among hydrology, biology, and eutrophication, with a variety of spatial trends throughout the system. It is known that excessive ponding depths and excessive dry-downs both tend to decrease peat accretion, due to lower plant productivity/turnover. In the case of dry-downs, this is exacerbated by increased oxidation. Higher phosphorus loads generally increase plant productivity/turnover, thus enhancing peat accumulation. Second, in the zone of new marine influence, freshwater peat is subject to seawater, which affects peat dynamics through a range of factors such as sulfate reduction. And third, mangrove peat is subject to rising sea level in combination with hurricanes. Without data to calibrate the dynamics of these three peat zones, our peat accretion results are particularly speculative. For this reason, we suggest the reader interpret slower peat accretion rates in our simulations as indicators of peat stress rather than rates in a more literal sense.
Succession of all habitat types in the ELM are determined by interactions among a suite of three parameters: (1) water depth and duration, (2) soil phosphorus concentration and duration; and (3) surface water chloride concentration and duration. For simplicity, here we aggregated four mangrove habitat-types into a single mangrove habitat class, and likewise aggregated all vegetated freshwater habitat-types into one class. With a 500 m grid resolution, the regional model has a relatively coarse scale that is nevertheless useful for evaluating trends across broad spatial regions, particularly across decadal time scales. Our succession dynamics appear reasonable; for instance, the simulated landward extent of mangroves after 3 decades (1965–2000) in the Baseline scenario simulation generally followed the contour of observed mangrove extent in 1995.
Additional details on the workings of the ELM and how it was adapted for use in this project can be found in Online Resources 2 and 3 in the supplemental material provided with this paper.
Results
Our key results are described below for individual hydro-ecological metrics and presented in Figs. 3–13 (complete quantitative performance measure graphics are available online at http://www.ecolandmod.com).

Simulation maps of daily mean surface water salinity for the Baseline scenario (top left), the decreased rainfall scenario (−RF, bottom left), and the increased rainfall scenario (+RF, bottom right)

Daily mean surface water depths relative to land surface (top), and daily mean surface water salinity (bottom) for FCE LTER sites along the Shark River Slough transect (with relative positions indicated in the scale provided)

Daily mean surface water surface water salinity shown over the period of simulation for three locations along the Shark River Slough transect, top: FCE LTER sites SRS-3, middle: a location between FCE LTER sites SRS-3 and −4, and bottom: FCE LTER site SRS-4

Simulation maps of daily mean surface water depth relative to land surface for the Baseline scenario (top left), the decreased rainfall scenario (−RF, bottom left), and the increased rainfall scenario (+RF, bottom right)

Simulation maps of daily mean peat accumulation rate for the Baseline scenario (top left), the decreased rainfall scenario (−RF, middle left), and the increased rainfall scenario (+RF, middle right), with the differences between the climate change scenarios and the Baseline scenario depicted in the bottom row, with decreased rainfall (−RF) at bottom left, and increased rainfall (+RF) at bottom right

Maps showing the differences between the climate change scenarios and the Baseline scenario in terms of surface water velocity for the decreased rainfall scenario (-RF, left) and the increased rainfall scenario (+RF, right). For reference, the saltwater front is shown as a red contour, and the 0.40 m topographic contour is shown in orange

Maps showing the differences between the climate change scenarios and the Baseline scenario in terms of surface water phosphorus concentration for the decreased rainfall scenario (−RF, left) and the increased rainfall scenario (+RF, right). For reference, the saltwater front is shown as a red contour, and the 0.40 m topographic contour is shown in orange

Maps showing the differences between the climate change scenarios and the Baseline scenario in terms of phosphorus accumulation rate for the decreased rainfall scenario (-RF, left) and the increased rainfall scenario (+RF, right). For reference, the saltwater front is shown as a red contour, and the 0.40 m topographic contour is shown in orange

Simulation maps of habitat distribution for the Baseline scenario (top left), the decreased rainfall scenario (-RF, middle left), and the increased rainfall scenario (+RF, middle right)

Sankey diagram showing areal coverage of the three habitat classes for the Baseline scenario (center column), the decreased rainfall scenario (−RF, left column) and the increased rainfall scenario (+RF, right column) with estimated exchanges illustrated

Cartoons illustrating broad differences in habitat distribution among scenarios a Baseline, b decreased rainfall, −RF, and c increased rainfall, +RF; depicted along the Shark River Slough transect, with the numbers at the bottom corresponding to FCE LTER monitoring station site numbers (the SRS prefix for site names is omitted here). The geographic location of the Shark River Slough transect and the numbered FCE LTER sites are provided in Fig. 2
We provide three overlays on all of our simulation maps because we found them to provide useful references for comparisons among maps (Figs. 3, and 6–11): (1) the inland contour of 0.40 m elevation; and (2) the FCE LTER transect sites, with site SRS-3 labeled; and (3) a vector delimitation of the saltwater front in the surface water. We designate the saltwater boundary (0.18 psu, or chloride concentration of 100 mg/L) by simply doubling the typical Everglades fresh surface water salinity (which is given as ~50 mg/L chloride concentration by Price and Swart (2006)). As defined, our saltwater front is close to the extreme limit of marine influence; for comparison, seawater has a chloride concentration of 19,400 mg/L.
Surface Water Salinity
Relative to the Baseline scenario, the saltwater front transgressed up to 15 km inland in both scenarios, with little difference between climate change scenarios (Fig. 3). Its relationship to the 0.40 m topographic contour can be considered using four spatially defined groups: (1) in the northwest portion of the study area, the saltwater front and topographic contour coincide, (2) in the area northeast of Lostman Creek (i.e., the area marked by a white box in Fig. 2), the saltwater front bows seaward with respect to the topographic contour, particularly in the +RF scenario, (3) from the Lostman Creek area, across Shark River Slough, and through Taylor slough, the saltwater front is 2–3 km inland of the topographic contour, and (4) in the eastern edge of the study area near canals, the gap widens as the saltwater front bends sharply north approximately 10 km inland of the topographic contour.
Despite the similarity of the landward incursion of the saltwater front between the two climate change scenarios, the salinity gradient from this boundary seaward is very different for the −RF and +RF scenarios. For instance, at FCE LTER monitoring site SRS-3 the surface water has approximately 1.8 psu salinity in the +RF scenario, and 9 psu in the -RF scenario (Fig. 4). For comparison, seawater has 35 psu. This 7 psu offset in surface water salinity persists to the coast, with the -RF scenario becoming full strength seawater at the end of the transect and +RF ending at 25 psu. The shape of the salinity curve is nearly identical for the two scenarios, but the 7 psu offset means that the +RF scenario curve is shifted 5–10 km downstream compared to the −RF scenario. To get the most relevant salinity values for our SRS transect sites we used ELM outputs from canal/river/creek vectors, because the monitoring sites are at or near the margins of Shark River. Being at slightly higher elevation, the surrounding area exhibits lower salinity values than the river transect, but the trend of lower salinities in the +RF scenario compared to the −RF scenario can be seen throughout the marine-influenced zone in our study area (Fig. 3).
Temporal patterns in surface water salinities also exhibit a milder salinity regime for +RF compared to −RF. For instance, the daily pattern of salinity at three locations along the Shark River Slough transect (Fig. 5) shows pulses of freshwater associated with rainfall events and related managed inflows. At the upstream FCE LTER site SRS-3 location, the Baseline scenario exhibits frequent intra-annual surface water salinity spikes that are relatively subtle compared to much higher (18–22 psu) spikes at the same points in time for the −RF scenario. The +RF scenario exhibits an intermediate response, sometimes producing spikes similar but somewhat dampened compared to the −RF scenario, and at other times closely adhering to the Baseline scenario pattern of very low magnitude spikes. At a location further downstream (between SRS-3 and −4), the Baseline scenario changes little, the +RF shifted its minimum value upward slightly, and the −RF pattern changed greatly to a typical salinity of 22 with frequent brief excursions to below the saltwater threshold as defined in this paper. Further downstream at the FCE LTER site SRS-4, the Baseline scenario is typically just above the saltwater threshold but the two climate changes scenarios are typically much higher (18–22 psu), with the −RF scenario being frequently at the higher end of the range, and the +RF scenario at the lower end with frequent low-salinity excursions.
Water Depth
The three scenarios exhibit distinct water depth patterns as viewed in the simulation maps (Fig. 6) and a graph of surface water depth for the Shark River Slough transect (Fig. 4). In the freshwater remnant, the +RF scenario has only slightly elevated water depths compared to the Baseline, whereas the −RF scenario has fresh water levels more than 10 cm lower than the Baseline, and lower than the +RF even at the marine end of the transect. In the marine-influenced portion of the simulation maps, water depth reaches greater depth at the coastline, for +RF compared to −RF (70 cm and 80 cm higher than the Baseline scenario for the −RF and +RF scenarios, respectively). The scarp where water depth breaks from the relative flatness of the freshwater remnant to the more steeply deepening water in the marine influence zone coincides with the saltwater front in most places for both climate change scenarios. In the +RF scenario, there are two places where the scarp occurs inland of the saltwater front: in the area northeast of Lostman Creek (marked with a white box in Fig. 2) it occurs at the 0.040 m topographic contour, and in Shark River Slough it occurs 1–2 km inland of the 0.40 topographic contour.
Peat Accretion
Here and in Fig. 7, we report our peat accretion simulations as rates, but we remind the reader that in light of the current limitations of the peat accretion module, we consider our peat accretion rates to be best read as indicators of peat stress rather than rates per se. Within the remnant freshwater habitat, peat accumulation rates are similar for the Baseline and +RF scenarios, and modestly elevated for the −RF scenario (Fig. 7). For both climate change scenarios, a wide swath of slower peat accumulation rates is visible in the new marine-influence zone. In Shark River Slough the most intense slowing of peat accretion occurs in a narrow arc just seaward of the saltwater front, and a mixed pattern (increases and decreases) can be seen seaward of that band.
Surface Water Velocity
The coastal region in both climate change scenarios exhibits more rapid surface water flow rates compared to the Baseline scenario (Fig. 8). The most consistent velocity difference between the two climate change scenarios is found in the remnant freshwater region. In the −RF scenario, surface water velocity slowed by more than 75 m/d compared to the Baseline scenario through most of the freshwater portion of Shark River Slough and the broad flow-way to the northwest.
This trend of diminished flow rates extends through the freshwater remnant. In the +RF scenario, surface water velocity increases by 20–30 m/d through most of the freshwater remnant (compared to the Baseline scenario), but slows by more than 50 m/day in a broad area bound by the scarp of more steeply increasing water depths (see Fig. 6).
Phosphorus
In fresh surface water, phosphorus concentration in the +RF scenario is similar to the Baseline scenario, except for an increase of up to 5 μg/L in the Long Pine Key area (Fig. 9; location of Long Pine Key is provided in Fig. 2). In the −RF scenario, Shark River Slough exhibits a slightly elevated surface water phosphorus concentration (1–3 μg/L), while the rest of the freshwater remnant exhibits a slightly lower phosphorus concentrations (up to 10 μg/L in part of the Long Pine Key area), compared to the Baseline scenario. The pattern in the marine-influenced subregion is similar between climate change scenarios, with elevated phosphorus concentrations in surface water compared to the Baseline scenario particularly at the coastal margins and in the band between the saltwater front and the 0.40 m topographic contour (except for the area northeast of Lostman Creek, marked with a white box in Fig. 2).
In terms of phosphorus accumulation in the soil of the freshwater remnant, the −RF scenario exhibited a mix of higher and lower rates compared to Baseline, whereas in the +RF scenario phosphorus accumulation in the soil increased substantially compared to the Baseline scenario in most of the freshwater remnant (Fig. 10). In both climate change scenarios, surface water phosphorus concentrations exhibit changes particularly in proximity to the saltwater front, either reversing or intensifying adjacent patterns. The narrow band between the saltwater front and the 0.40 m topographic contour in the vicinity of Long Pine Key exhibits higher phosphorus accumulation (>15 mg/m2/year), but as this band bends north toward Shark River Slough it abruptly shifts to lower phosphorus accumulation compared to the Baseline scenario. In the coastal area in both climate change scenarios, phosphorus concentration in surface water and phosphorus accumulation in the soil both increased.
Habitat Distribution
Changes in the distribution of freshwater habitat, mangrove forest, and open water are displayed in simulation maps (Fig. 11), with areal extent of habitats for the two climate change scenarios compared to the baseline values in a Sankey diagram (Fig. 12), and the habitat variation along the Shark River Slough transect for the three scenarios illustrated in cartoons (Fig. 13). Mangroves migrated up to 15 km inland in both climate change scenarios, stopping 1–2 km seaward of the saltwater front (except in the area northeast of Lostman Creek, marked with a white box in Fig. 2, where the mangrove boundary is as much as 10 km seaward of the saltwater boundary) (Fig. 11). A quarter or more of the initial freshwater habitat was lost (25% for +RF scenario, 30% for −RF scenario; Fig. 12), with more freshwater habitat visible along Shark River Slough and in the area northeast of Lostman Creek, marked with a white box in Fig. 2, in the +RF scenario (see gray curves applied to the Baseline scenario map on Fig. 11).
Because of conversion from freshwater habitat, mangroves made a net gain in areal coverage in the −RF scenario (130% increase) (Fig. 12). Nonetheless, significant mangrove forest was lost to open water under both climate change scenarios, particularly along southern coast where the Everglades meets Florida Bay, and extended northwestward to connect with Whitewater Bay (location of Whitewater Bay is provided in Fig. 2). In the +RF scenario, open water spread in a near-continuous swath to Shark Rive Slough and somewhat beyond, divided from the open water in the northwest corner of the ENP by the preservation of terrestrial habitat in the area near Lostman Creek (Fig. 11; area near Lostman Creek marked with a white box in Fig. 2). The arc of open water traces the inland valley of near-sea level elevation between the Gulf Coast of the Everglades and the mainland visible in Fig. 2.
Our transect through Shark River Slough provides a useful comparison among the three scenarios for habitat changes, as exhibited in the simulation maps (Fig. 11) and as illustrated in Fig. 13. The 13.4 km segment between FCE LTER monitoring sites SRS-3 and SRS-4 is freshwater habitat in the Baseline scenario, mangrove forest in the −RF scenario, and open water with small isolated patches of mangroves in the +RF scenario. In the +RF scenario mangroves recede seaward in Shark River Slough, with open water meeting freshwater habitat near the saltwater front, and extending south nearly to SRS-5.
In Taylor Slough, the two climate scenarios had more similar effects to each other compared to the Baseline. In the Baseline scenario (and in the ENP today), FCE LTER site TS/Ph-3 is just north of the boundary between freshwater habitat and mangrove forest, with the latter occupying the final ~15 km to the coast (Fig. 11). In the two climate change scenarios, open water replaces most of that mangrove forest, and most of the freshwater habitat in Taylor Slough between TS/Ph-3 and TS/Ph-2 is replaced by new mangrove forest. The net effect is that in the climate change scenarios the net areal coverage is similar to Baseline but mostly consists of new mangrove forest, with old mangrove forest largely converted to open water. The main difference between the climate change scenarios in the southeast is that a larger “island” remnant of the southeastern freshwater marsh remains in the +RF scenario (Fig. 11).
Discussion
Our ELM simulations provide a glimpse of potential hydro-ecological changes in the ENP that might accompany a SLR of 0.5 m in combination with a warming of 1.5 °C and a subsequent increase of 7% evapotranspiration. Further, by comparing an increase or decrease of rainfall by 10%, our simulations offer indications as to the sensitivity of the system to freshwater availability in the context of future warming and SLR. It is essential to bear in mind that, as with all scenario evaluations, our simulated hydro-ecological changes are rough plausible responses relative to a baseline for comparison and are not intended to be predictions. The goal of any such scenario evaluation is to gain a better understanding of some potential system responses to perturbations, along with providing improved perspectives on the uncertainties associated with future hydro-ecological dynamics. Below we will discuss what we glean from our simulations in terms of three themes (a) Incursion of the saltwater front, (b) Loss of mangrove fringe, and (c) Climate effects on the freshwater remnant.
Two overarching themes emerge from our results, which we will expand upon in our discussion. First, sea level rise caused the greatest impact in our climate change scenario simulations compared to the Baseline scenario, driving the most obvious changes to all of our hydro-ecological metrics in the ENP. Despite this, our simulations make it clear that microclimate (specifically, increase or decrease of rainfall in our simulations) has a profound effect on ecological outcome.
Saltwater Intrusion and its Consequences
Saltwater intrusion
The saltwater front advanced up to 15 km inland in our two climate change scenarios (this boundary is shown as a red contour in all of our simulation maps, Figs. 3, 6–11). It has long been recognized that wetlands in low-elevation areas are endangered by SLR through inundation, erosion, and salinization (Gornitz 1991). The southern Everglades is particularly vulnerable to marine encroachment because it is low-lying and flat (Price et al. 2006).
The location of the saltwater front is similar between the two climate change scenarios, is closely associated with the 0.40 m topographic contour, and agrees broadly with the landward inundation of seawater estimated in a study using LIDAR elevation data and an imposed 0.50 m SLR (Zhang 2011). The places where the saltwater front deviates from the 0.40 m topographic contour provide clues as to additional influences on the encroachment of the saltwater. First, in our two climate change scenarios, the saltwater front bows seaward of the 0.40 m topographic contour in the area northeast of Lostman Creek (particularly in the +RF scenario; Fig. 3) This may be in response to runoff from increased rainfall funneling out of that secondary slough (mainly from unmanaged flows from southeast Big Cypress National Preserve; location of preserve is shown in Fig. 1). Second, in the area around Long Pine Key (location of Long Pine Key is provided in Fig. 2), the fact that the saltwater front is further inland than the 0.40 m topographic contour may be due to comparatively less freshwater downstream flows in this region (Fig. 3).
Third, in the eastern part of the study area where the saltwater front bends northward this appears to be a response to salinization related to the canals “short-circuiting” what would otherwise be overland marsh flows. Historically, canals open to the sea brought saline water inland in the southern Everglades (Fitterman and Deszcz-Pan 1998). Although such canals in the ENP have since been plugged or gated to prevent further inland flow of saltwater, a concern from the beginning of CERP has been the potential for SLR to exceed the gates on the existing canals (USACE 1999).
Our saltwater front was defined at a salinity so low that it would pass the EPA drinking water standard (which is ≤ 250 mg/L chloride concentration (EPA 2013)). It is categorized in marine science as the threshold between “fresh” and “oligohaline” water is most marine water classification systems (Caljon 2012; Venice_System 1959). Although delineated at extremely low in salinity, our saltwater front effectively doubles as the boundary for the most intense changes in all of our hydro-ecological metrics (Figs. 3, 6–11).
Peat stress in the marine-influenced subregion
Through vertical accretion of organic matter and storm-derived sediment, ground surface in coastal wetlands has a limited potential to keep pace with SLR (NRC 2014b). Encroachment of seawater into previously freshwater regions can exacerbate peat subsidence due to salinization, sulfate reduction and drowning of vegetation. Peat accretes vertically by a combination of sediment deposition and subsurface accumulation of plant detritus and roots. Under excessive flooding, plant growth declines, and sediment elevation can lag behind SLR, creating an accretion deficit. If the deficit widens, plant stress can lead to death of plants, collapse of sediment volume, and submergence. The potential for encroaching seas to trigger peat collapse is a critical concern, and there are some documented cases of it having already begun in some parts of the coastal Everglades, but the dynamics are incompletely understood (Chambers et al. 2014; Hackney and Williams 2012; NRC 2014a).
As may be expected, then, peat stress (expressed as acute slow-down of peat accretion in our simulations) appears to be particularly high in the zone of newly deepened and salinized water occupying what is today freshwater marsh for our two climate change scenarios (Fig. 7). Throughout most of the new marine-influenced subregion, peat accumulation rates tended to slow compared to Baseline rates due deterioration of freshwater habitat, altered nutrient availability, and increases in water depth and salinity, which in turn decrease plant productivity and turnover averaged over decadal time scales. Although the ELM incorporates many of the suspected interactions that are thought to drive peat collapse, in the absence of further process-based research results, the model results are best understood as indications of intensity of peat stress rather than specific quantification of this important consequence of sea level rise. Ongoing research should provide enhanced guidance for the ELM modules.
Limitations of the model such as the abrupt SLR were previously noted, and the ramifications on peat stress should be borne in mind. The complexity of the water management system made us reliant upon the SFWMM scenarios-based water level distributions and control structure flows to drive the managed water control structure flows within the ELM domain. In order to use the SFWMM output it was necessary to adopt its assumptions, including the full SLR of 0.46 m occurring all at once on “day 1” of the simulation, i.e., as an initial condition in the climate change scenarios. The benefit of the realistic flows and water levels provided by the SFWMM hydrologic simulations were deemed worth the trade-off in SLR rate.
While justifiable for a screening-level analysis (Obeysekera et al. 2015), and necessary from a technical perspective for this project, it is important to note that this instantaneous increase of sea level has significant ecological impact on the ELM simulations for our climate change scenarios. We understood a priori that such an abrupt SLR would rapidly kill off freshwater vegetation in reality, as it does in the model. Such dramatic reduction in plant productivity/turnover leads to low (or negative due to decomposition processes) peat accumulation: prolonged duration of this multi-year dynamic will reduce the long term accumulation rate. We understand that the severe sea level perturbation is unrealistic, and there are no field data available to determine appropriate recovery times to such a severe sea level perturbation. Our results must be understood within this limitation.
Mangrove encroachment
During the 20th century mangroves in the southern Everglades have already migrated inland at the expense of freshwater marsh in many areas while declining in coverage along their seaward fringe (NRC 2014b; Ross et al. 2000; Wanless et al. 1994). In our simulations, marine influence spread over 25% or more of the ENP freshwater habitat (Figs. 11–13). The landward boundary of mangroves is extremely similar between the two climate change scenarios, appearing to be largely driven by the saltwater front (see gray curves applied to the Baseline scenario map on Fig. 11). However, the additional water depth exhibited by the +RF scenario appears to inhibit mangrove establishment, particularly in Shark River Slough (Figs. 6, 11, 13). In our Shark River Slough transect the segment between FCE LTER monitoring site SRS-3 and the downstream site SRS-4 is occupied by freshwater habitat in the Baseline scenario, mangrove forest in the −RF scenario, and mostly open water in the +RF scenario. Indeed, in the +RF scenario, mangrove forest retreats seaward to FCE LTER SRS-5 along our Shark River Slough transect. The unrealistically abrupt rise in sea level, discussed in the previous section with regard to its implications for peat stress, may also have contributed to the excess open water in the +RF scenario.
Rainfall influence on salinity regime
A future reduction in precipitation (which may well exceed 10%) is considered more likely for the southern region of the state and is thought to pose the greatest challenges to preserving and restoring Everglades (Nungesser et al. 2015; Obeysekera et al. 2015; Obeysekera et al. 2011). A rainfall increase of 10% has been called a less likely “best case scenario” for the Everglades (Obeysekera et al. 2015), potentially “holding the sea at bay” in some ways (Gaiser et al. 2012; NRC 2014b; Saha et al. 2012). Our results for surface water conditions do provide some indications (“good news/ bad news” if you will) as to how an increase in future rainfall may mitigate coastal wetland impacts from SLR, warming and increased evapotranspiration. We have noted the “bad news” above: increased freshwater availability does little to mitigate the boundaries of marine influence and freshwater habitat loss, its additional water depths may exacerbate peat stress; greater open water expanse is a possible outcome. The “good news” is that the salinity regime within the marine-influenced zone was markedly less severe both spatially and temporally for the +RF scenario compared to the −RF scenario.
Throughout the zone of marine-influence, each subsequent threshold of salinity is met several kilometers further seaward for the +RF scenario compared to the −RF scenario (Fig. 3). The spatial salinity gradient is likely to drive biological response and the severity of ecological stress related to SLR. It is known that different biota have different tolerances of salinity, and for many species there is a threshold of salinity above which adverse effects are severe. For some species the tolerance threshold is close to the freshwater end of the mixing continuum (Schallenberg et al. 2001). Investigations have been undertaken to identify salinity thresholds for important coastal Everglades flora and fauna such as brackish sawgrass, so as to predict ecological impacts from SLR (Stabenau et al. 2011); our salinity gradients can inform biological projections for possible ecosystem responses in the coming decades.
Temporal variability of salinity is also ecologically important, and our two climate change scenarios produce distinct salinity patterns with respect to seasons, rainfall events, and tidal influence (Fig. 5). In recent decades, the coastal Everglades has already shifted to a salinity regime featuring more frequent high salinity events and fewer low salinity events, and the subsequent shifts of freshwater flora and fauna to salt-tolerant communities render them unrecognizable today (USACE 1999). Although both of our climate change scenarios continue this trend landward (Fig. 5), it is much less severe in the +RF scenario. Shortening the duration of elevated salinity by days can drastically reduce mortality rates among salt-sensitive macrophytes (Schallenberg et al. 2001). The lower average salinity conditions and increased influence of upstream flow dynamics under the +RF scenario would likely result in significantly different zonation of biological communities in the marine-influenced zone compared to the −RF scenario. Our results support the idea that restored sheet flow across the Everglades may provide lower salinity and less frequent high salinity events.
Phosphorus in the marine-influenced subregion
The ELM sets seawater phosphorus concentration higher than that of incoming freshwater, consistent with available literature (Brand 2002; Zapata‐Rios et al. 2012). For this reason, the marine zone surface water is higher in phosphorus concentration, and greater phosphorus accumulates in the affected region (Figs. 9, 10). The substantial spatial heterogeneity of phosphorus metrics in the marine-influenced subregion stemmed from interactions of multiple hydro-ecological processes, an important area for future research, in terms of both modeling and field work.
Loss of Mangrove Fringe
At some point, fragmentation and drowning of mangroves is expected for the coastal Everglades (Pearlstine et al. 2010; Saha et al. 2011). The rate of SLR that Everglades mangroves can withstand without drowning is as yet uncertain, due in large part to uncertainties related to peat dynamics discussed earlier. In some Everglades mangrove swamps, mangrove peat elevation is already changing: storm deposits have substantially increased the accretion rate (Smoak et al. 2013), while hurricane surges have devastated some southwestern mangrove forests, resulting in rapid loss of surface elevation (Smith et al. 2009). The pivotal roles of hurricanes for either building up or tearing down mangrove peat make it particularly hard to estimate changes in peat accretion rate with climate change.
The geologic record provides clues as to the average SLR rates that Everglades mangroves can accommodate. Coming out of the last ice age, sea level rose at an average rate of 2.5–5 mm/year, a rate too fast for mangroves to stabilize (Wanless et al. 1994). The historical Everglades mangrove fringe took hold during the last 3200 years, when the rate of SLR slowed to an average of 0.4 mm/year (Wanless et al. 1994). In the last century, sea level in south Florida began to rise more quickly, at an average rate of 3 ± 2 mm/year tidal gauges in south Florida (Wdowinski et al. 2016), and the average rate increased to 9 ± 4 mm/year in southeast Florida after 2006 (Wdowinski et al. 2016). Wanless et al. (1994) used the historical record to forecast that a SLR rate of 9 mm/year would bring “catastrophic inundation of southern Florida, loss of coastal wetlands, and loss of freshwater resources.”
Because mangrove peat accretion is still poorly understood, our simulations of mangrove response along Florida Bay mainly they serve to highlight differential vulnerability between the two main mangrove fringes (Fig. 11). Fortunately, the mangrove fringe along the ENP Gulf Coast is protected by greater elevations (mostly lying above 0.40 m elevation), and thus suffered little fragmentation or drowning in our climate change scenarios. But the mangrove fringe along the northern coast of Florida Bay suffered significant losses to open water in both of our climate change scenarios. Our Taylor Slough transect sites mark the displacement of mangrove forest upslope: with much of the older mangrove forest replaced by open water, and most of the areal coverage of mangrove being new growth across former freshwater habitat. Some of the original mangrove forest along Florida Bay remains as isolated patches.
While it is useful to envision possible habitat succession outcomes in the southern Everglades, the actual outcome of 0.5 m SLR is harder to predict. We must emphasize that while our final amount of SLR is reasonable, the rates are unrealistically severe and can be expected to produce more catastrophic ecological responses accordingly. A more gradual SLR may have provided more opportunity for peat accumulation to keep pace with added water, and for habitats to recover, adapt, or migrate. In addition, habitat succession dynamics are uncertain, given the novel changes in sea level and climate drivers, and mangrove succession dynamics are less developed in ELM compared to habitats such as cattail-sawgrass (e.g., Fitz and Sklar (1999) and Fitz and Trimble (2006)). Assimilation of more recent (FCE LTER and other) research results are being synthesized for incorporation in the ongoing update to the ELM v3.0. The potential for mangrove sediment accretion rate to keep up with SLR is still an open question, and we expect to update ELM modules as more peat research becomes available.
In addition to these modeling considerations, the southern mangrove swamp has some unique features that will affect its vulnerability to SLR. A source of increased vulnerability is the exceptionally low productivity in parts of this mangrove fringe (e.g., Taylor Slough), which may in turn hamper the prospect of peat accretion keeping pace with SLR (Gaiser et al. 2006). The biophysical mechanisms behind this low productivity are not well understood, and may therefore not be fully captured in the ELM. Conversely, a natural source of temporary protection from SLR along the northern coast of Florida Bay is provided by the 0.65–1.00 m elevation natural levee known as Buttonwood Ridge (too small to show in our location maps). This feature currently prevents tidal exchange between the wetland and Florida Bay except through the narrow channels that incise the ridge, such as Taylor River (Craighead 1964; Langevin et al. 2005; Stabenau et al. 2011). The Buttonwood Ridge makes it possible for a range of flora and fauna in Taylor Slough that are sensitive to saltwater to occupy this coastal region (Stabenau et al. 2011). Our ELM simulations are not able to include this critical but small-scale feature, due to limitations of the spatial resolution. The lack of this feature in our model means that our climate change scenarios to some extent (by default) simulate the severe hydro-ecological consequences of saltwater overtopping the Buttonwood Ridge. The interaction of rising seas and the Buttonwood Ridge will be a critical driver of coastal wetland response to climate change and SLR in the coming decades. It does seem logical that if SLR were to overtop the Buttonwood Ridge at some future time, this could trigger a fundamental regime change in the coastal region, initiating widespread tidal connectivity within the southern coastal Everglades, similar to what is found in the Shark River Slough today.
Human activity is an additional factor determining tidal wetland stability in the face of SLR that cannot be overlooked (Kirwan and Megonigal 2013). In our simulations, current water management rules are followed, and no restoration measures are included. By providing a vision of what could happen in the absence of restoration, it is our hope that these simulations may spur restoration strategies that may mitigate or delay some of these changes.
The pattern whereby mangroves expand inland while their seaward fringe deteriorates or recedes has already been observed in the Pacific coast of Mexico, in response to SLR and El Niño (López‐Medellín et al. 2011). The loss of seaward fringe is not fully compensated for by upslope migration of mangroves, even where there is a net gain in areal coverage, as we saw in our −RF scenario. New growth mangrove saplings lack the complexity of a mature mangrove forest and are unable to fulfill the same ecological role as an old growth stand. Recent studies have demonstrated that the most valuable ecosystem services (e.g., providing nurseries and feeding grounds for many fish and crab species, as well as coastal storm protection) are greatest at the interface of coastal water and the seaward fringes (Aburto-Oropeza et al. 2008). Such ecosystem services from fringe mangroves appear to decline in a non-linear fashion with distance inland for at least some mangrove forests (Barbier et al. 2008; Koch et al. 2009).
In both of our climate change scenarios, open water occupies the low-lying arc from northern Florida Bay to Whitewater Bay (Fig. 11), two bodies of water that were previously divided by land. In the +RF scenario the open water continues northward through a swath of Shark River Slough and the area near Lostman Creek (marked with a white box in Fig. 2). In the northwestern part of our study area, this low-lying arc appears as a narrow stretch of open water in the Baseline scenario. In our +RF scenario this arc of open water widens and extends further southward into the Lostman Creek area, with a patch of less than 20 km of mangrove forest preventing full connectivity from the northwest corner of the study area to Florida Bay.
Climate Effects on the Freshwater Remnant
We have noted that marine influence eliminates 25% or more of freshwater habitat in response to SLR, with little difference between climate change scenarios. However, differential rainfall causes the remnant freshwater habitat in the ENP to have very different surface water depth (and thus, it can be inferred, hydroperiod), peat accumulation rate, surface water velocity, and phosphorus content of both surface water and soil.
Some of the most important questions for ENP planning involve the dynamic surface of freshwater peat: How will it respond to climate change? How can restoration projects and water management changes minimize subsidence and foster vertical peat accretion? In general, long hydroperiod with moderate depths and higher phosphorus loads together set the stage for emergent vegetation productivity and subsequent vertical peat accretion. Dry-downs promote compaction through the collapse of pore spaces, and accelerated decomposition due to oxygen permeating to greater depths in the sediment column. Accordingly, the National Research Council forecasted accelerated peat decomposition for the freshwater Everglades under the future climate scenario referred to in this paper as the −RF scenario (NRC 2014a). They call for increases in depth of freshwater, to promote higher rates of peat accretion that may thereby mitigate some of impacts from SLR and associated saltwater intrusion. For this reason, the scenario of 10% increase in rainfall is widely characterized as the “best case scenario” for Everglades resilience in the decades to come (Nungesser et al. 2015).
In our +RF scenario peat accumulation rate exhibited negligible change compared to our Baseline scenario throughout the freshwater ENP (Fig. 7). Although contrary to expectations, this result is consistent with the observation that water depth was also very similar in the freshwater remnant in the +RF compared to the Baseline scenario (Fig. 4) due in large part to the assumption of present-day water management decisions built into the model. In our −RF the loss of over 10 cm in surface water depth compared to the Baseline scenario (Figs. 4, 6), was accompanied by very mild acceleration of peat accumulation rates through most of the freshwater remnant.
An additional anticipated benefit of increased freshwater to the Everglades is the potential for greater surface water velocity (Nungesser et al. 2015). An important and unique Everglades habitat type particularly prevalent in Shark River Slough is “ridge and slough landscape,” a wetland patterning of elongate sawgrass ridges alternating with slightly lower troughs (sloughs) which supports wading bird migration and reproduction (Larsen et al. 2011). These features are aligned in the direction of flow and require water velocities of >1 cm/s for healthy development and maintenance. Slowing of water flow rate due to compartmentalization and reduction of freshwater in the Everglades has threatened and in some cases erased this patterning (Larsen et al. 2011). Flow velocities currently vary seasonally from up to 2 cm/s in the wet season to <0.1 cm/s in the dry season (Riscassi and Schaffranek 2004). In our climate change simulations, surface water flow velocities slowed with less rainfall, and accelerated with more rainfall, but the magnitude was extremely modest (less than 1 μm/s).
The Everglades ecosystem is oligotrophic and phosphorus limited, but managed Everglades inflows may carry higher nutrient loads. A major challenge of CERP is increasing water flows without increasing the nutrient load to the ENP. Planning for CERP in the coming decades must balance the trade-offs between reducing phosphorus loads while increasing Everglades water inflows (Sklar et al. 2005). Even slight increases in water phosphorus concentration cause cascading effect in flora and fauna populations, periphyton type, and primary productivity (Gaiser et al. 2005; Richardson et al. 2007). In the +RF scenario, phosphorus concentration in fresh surface water exhibits little change (Fig. 9), but phosphorus accumulation in soil shows increases through most of the freshwater zone (Fig. 10). From a mass balance perspective, increase in water volume (while phosphorus concentration is held constant) increases the net amount of phosphorus delivered to the system. Because phosphorus is rapidly removed from the water column in this highly phosphorus limited system, phosphorus accumulation rate (or analogous metrics of phosphorus concentration in biota) is a more accurate descriptor of phosphorus eutrophication than highly transient metrics of phosphorus concentration in surface waters (Gaiser 2009). In our -RF scenario, the freshwater remnant appeared to have little net change in phosphorus content in surface waters or soil, with mild increases in some areas being roughly matched by mild decreases in other areas.
Management Implications
An important research priority is to better understand the conditions which favor peat accretion both in fresh and saline water (including vegetation response to salinity thresholds) so that water management and restoration efforts can implement strategies to enhance this important bulwark against SLR and climate change. Our simulations provide a vivid illustration of the potential for open water expansion if peat accretion rates fall behind the rate at which water depth increases (in response to both SLR and possible increased freshwater volume).
Our simulations indicate that the Everglades’ resilience to warming and SLR is highly sensitive to freshwater availability. Additional freshwater supply may profoundly mitigate the impacts of SLR by decreasing the average salinities within the marine-influenced zone, by decreasing the frequency of high salinity events, and by increasing the frequency of low salinity events. If rainfall increases in the future, or if freshwater flow increases through restoration, the marine-influenced subregion would undergo a more gradual transition to higher salinity regimes. In addition, our simulations illuminate the need for restoration planners to balance the benefits of greater freshwater flow with the risks associated with greater water depths.
When it is practical to do so, it may be useful to monitor the migration of the oligohaline transition zone accompanying SLR. Our saltwater front was defined by the extremely low salinity of 0.18 practical salinity units, or 100 mg/L chloride concentration (for comparison, seawater has 35 practical salinity units, or 19,400 mg/L chloride concentration), and yet it was strongly associated with impacts of peat stress, phosphorus content, and habitat succession. These impacts would be predicted to occur landward of the saltwater intrusion front as it is currently designated and monitored.
Both of our climate change scenarios resulted in similar encroachment of the saltwater front and the mangrove boundary. This serves as a reminder that some aspects of sea level rise may be inevitable. Strategies for reducing ecosystem vulnerability and mitigate impacts must be combined with strategies that promote adaptation and resilience in the face of future changes, including such measures as assisted migration. Managing for resilience may include identifying anthropogenic stressors that can be reduced, and key ecosystem features that can be protected (West et al. 2009). The regime shifts in our simulations are a reminder of the increasing number of species likely to be stressed beyond their ability to recover. Several strategies have been proposed by West et al. (2009), Pearlstine et al. (2010) for promoting species survival in the long term. Periodic scenarios modeling of both climate and restoration scenarios can contribute to ongoing adaptive planning efforts by informing monitoring designs, identifying data gaps for long term hydro-ecological monitoring efforts such as FCE LTER, and suggesting modifications in management and restoration.
Limitations
This project represents screening-level analysis of potential landscape responses to future climate and sea level scenarios. Our simulations should be viewed as broad brush strokes of plausible outcomes, to spur discussion and prioritize future research, rather than a source of quantitative information. The aspect of the ELM most closely calibrated for use in this project was salinity (see Online Resource 3 in supplemental materials). We have noted simplifying assumptions that went into the ELM, including unrealistically abrupt sea level rise in our simulations, and incomplete understanding of peat dynamics. In addition, our scenarios all assume current water management, and do not include any restoration strategies included in scenarios. Consequently, increased rainfall did not necessarily result in ecosystem benefits anticipated for increased freshwater availability (e.g., water is potentially “wasted” by diversions from the Everglades under current management criteria). One of the Everglades restoration (CERP) goals is to increase water deliveries to ENP. Restoration targets (and updated CERP simulations) for flows to ENP are roughly 80% above the Base condition flow (NRC 2010), whereas the +RF scenario reflects only ~35% increases in flows to ENP. Moreover, CERP water management timing, magnitudes, and spatial distributions are quite different from current water management, so the +RF scenario is a relatively modest proxy for increased restoration flows to ENP.
Other limitations include uncertainty in climate and sea level rise projections. Most importantly, it should be noted that precipitation may increase or decrease by much more than 10%, so climate change effects on the Everglades may be more consequential than our simulations suggest. A related limitation in our model is the effects of internal climate variability introduced by multi-decadal oscillations in ocean temperature. Our model’s interranual variability was simulated using climate data for the region for the period 1965–2000; the years 1965–1994 were marked by a negative phase Atlantic Multidecadal Oscillation (AMO). In this sense, our future simulations are frozen in a negative AMO. In the past, change to a positive AMO has been associated with substantial increases in wet season rainfall, and doubling of net average annual inflow into Lake Okeechobee (Enfield et al. 2001).
While our simulations demonstrate the sensitivity of the system to ±10% rainfall, they provide indications as to trajectories of change that might accompany greater changes in rainfall. For instance, the shallow salinity gradient and the inland arc of open water in our +RF scenario would likely be more extreme with a greater increase in rainfall. With a greater decrease in rainfall, the steep salinity gradient revealed in our −RF scenario may be yet steeper.
More intense hurricanes are also associated with positive AMO phase, and with the longterm trend in ocean heat gain. While outside of the scope of the model as it currently stands, increased hurricane intensity has the potential to overshadow the effects of warming, sea level rise, and annual rainfall on the Everglades. The impacts of hurricanes extend beyond the climate parameters currently captured in the model, and include such effects as sediment deposition by storm surge, mangrove mortality, and coastal erosion.
Our scenarios also lack changes in seasonality; the timing and seasonal distribution of rainfall are as important to Everglades habitats as the annual amount. Our simulations lack a sub-surface saltwater intrusion component, which could affect both salinity and nutrient content in the overlying surface water, potentially accelerating landward encroachment of marine influence. Currently our ecological modules lack non-linearity of response; important changes such as habitat regime shifts and peat collapse can be sudden. One of the consequences of increased air temperature is increased surface water temperature, which is not included in our model at this stage. Warmer water temperatures would have far-reaching effects on microbial activity, phosphorous sorption dynamics in sediment, and distribution of fauna such as temperature-sensitive fish. Further work is needed to encode and calibrate such modules.
Future work can reduce uncertainties, add detail to landscape-scale modeling of climate effects, and add new modules for better characterization of ecological dynamics using the ELM.
Conclusions
Our simulations provide screening-level visions of how the Everglades may respond hydro-ecologically to sea level rise in combination with changes to macroclimate, and in so doing underline the need for future restoration planning to take these interacting factors into account. It has been said that “restoration under climate change is more important than ever before and might be most properly defined in terms of reducing ecosystem vulnerability and promoting adaptation and resilience” (Pearlstine et al. 2010). If the Everglades coastal wetland is a sentinel of climate change, our simulations make it clear that macroclimate changes such as temperature and rainfall regime are major drivers of vulnerability and resilience. Restoration planners cannot afford to overlook the interactions of these simultaneous threats as they look to the coming decades. By “visioning” the future of the Florida coastal Everglades our over-arching aim is to enable decision-making and positive actions in the face of the uncertainties associated with the unfolding threats of sea level rise combined with climate change in the coming decades.
References
Aburto-Oropeza O, Ezcurra E, Danemann G, Valdez V, Murray J, Sala E (2008) Mangroves in the Gulf of California increase fishery yields. Proc Natl Acad Sci USA 105:10456–10459
Aumen NG, Havens KE, Best GR, Berry L (2015) Predicting ecological responses of the Florida Everglades to possible future climate scenarios: Introduction. Environ Manag 55:741–748
Barbier EB et al. (2008) Coastal ecosystem-based management with nonlinear ecological functions and values. Science 319:321–323
Brand LE (2002) The transport of terrestrial nutrients to South Florida coastal waters The Everglades. Florida Bay, and Coral Reefs of the Florida Keys CRC Press, Boca Raton, FL, p 353–406
Caljon A (2012) Brackish-water phytoplankton of the Flemish lowland, vol 18. Springer Science & Business Media, The Hague, Netherlands
Carter L et al. (2014) Southeast and the caribbean climate change impacts in the United States: The third national climate assessment. 396–417
Catano CP et al. (2015) Using scenario planning to evaluate the impacts of climate change on wildlife populations and communities in the Florida Everglades. Environ Manag 55:807–823
Chambers LG, Davis SE, Troxler T, Boyer JN, Downey-Wall A, Scinto LJ (2014) Biogeochemical effects of simulated sea level rise on carbon loss in an Everglades mangrove peat soil. Hydrobiologia 726:195–211
Craighead F (1964) Land, mangroves and hurricanes. Fairchild Tropical Garden Bulletin, 19; 5-32. 1971. The trees of South Florida. The natural environments and their succession, vol 1. University of Miami Press, Coral Gables, FL
Enfield DB, Mestas‐Nuñez AM, Trimble PJ (2001) The Atlantic multidecadal oscillation and its relation to rainfall and river flows in the continental US. Geophys Res Lett 28:2077–2080
EPA (2013) Secondary Drinking Water Regulations: Guidance for Nuisance Chemicals. U.S. Environmental Protection Agency: Secondary drinking water regulations; guidance for nuisance chemicals Report 816–f–10–079. https://www.epa.gov/dwstandardsregulations/secondary-drinking-water-standardsguidance-nuisance-chemicals. Accessed 8 Jan 2017
Fernald E, Purdum E (1998) Water Resour Atlas, ch. 6. Florida State University: Institute of Public Affairs Tallahassee, FL, p 114–119
Fitterman DV, Deszcz-Pan M (1998) Helicopter EM mapping of saltwater intrusion in Everglades National Park, Florida. Explor Geophys 29:240–243
Fitz C, Sklar F, Waring T, Voinov A, Costanza R, Maxwell T (2004) Development and application of the Everglades Landscape Model. In: Landscape Simulation Modeling. Springer, New York, NY, p 143–171
Fitz HC, Kiker GA, Kim J (2011) Integrated ecological modeling and decision analysis within the Everglades landscape. Crit Rev Environ Sci Technol 41:517–547
Fitz HC, Paudel R (2012) Documentation of the Everglades landscape model: ELM v2.8.4 Ft Lauderdale Research and Education Center, IFAS, University of Florida, Ft Lauderdale, FL, http://wwwecolandmodcom/publications 364 pages
Fitz HC, Sklar FH (1999) Ecosystem analysis of phosphorus impacts and altered hydrology in the Everglades: a landscape modeling approach. In: Reddy KR, O’Connor GA, Schelske CL (eds) Phosphorus Biogeochemistry in Subtropical Ecosystems. Lewis Publishers, Boca Raton, FL, p 585–620
Fitz HC, Trimble B (2006) Documentation of the Everglades landscape model: ELM v2. 5. South Florida Water Management District, West Palm Beach, FL
Gabler CA et al. (2017) Macroclimatic change expected to transform coastal wetland ecosystems this century. Nat Clim Change 7:142–147
Gaiser E (2009) Periphyton as an indicator of restoration in the Florida Everglades. Ecol Indic 9:S37–S45
Gaiser E, Childers D (2016) Water Quality Data (Extensive) from the Shark River Slough, Everglades National Park (FCE), from October 2000 to Present. Environmental Data Initiative. doi:10.6073/pasta/4606a6abdda742225de953d079934ce4. Accessed 8 Jan 2017
Gaiser E et al. (2012) The Florida Everglades Wetland Habitats of North America Ecology and Conservation Concerns. In: Batzer D, Baldwin A (eds) University of California Press, Berkeley and Los Angeles, California
Gaiser EE et al. (2005) Cascading ecological effects of low-level phosphorus enrichment in the Florida Everglades. J Environ Qual 34:717–723
Gaiser EE, Zafiris A, Ruiz PL, Tobias FA, Ross MS (2006) Tracking rates of ecotone migration due to salt-water encroachment using fossil mollusks in coastal South Florida. Hydrobiologia 569:237–257
Gornitz V (1991) Global coastal hazards from future sea level rise. Palaeogeogr Palaeoclimatol Palaeoecol 89:379–398
Hackney CT, Williams A (2012) Impact of Sea Level Rise and Salt Intrusion On Everglades Peat: Review and Recommendations Jacksonville. Department of Biology, University of North Florida, FL, Online at http://14123210
Havens KE, Steinman AD (2015) Ecological responses of a large shallow lake (Okeechobee, Florida) to climate change and potential future hydrologic regimes. Environ Manag 55:763–775
Kirwan ML, Megonigal JP (2013) Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504:53–60
Koch EW et al. (2009) Non‐linearity in ecosystem services: temporal and spatial variability in coastal protection. Front Ecol Environ 7:29–37
Langevin C, Swain E, Wolfert M (2005) Simulation of integrated surface-water/ground-water flow and salinity for a coastal wetland and adjacent estuary. J Hydrol 314:212–234
Larsen L et al. (2011) Recent and historic drivers of landscape change in the Everglades ridge, slough, and tree island mosaic. Crit Rev Environ Sci Technol 41:344–381
López‐Medellín X, Ezcurra E, González‐Abraham C, Hak J, Santiago LS, Sickman JO (2011) Oceanographic anomalies and sea‐level rise drive mangroves inland in the Pacific coast of Mexico. J Veg Sci 22:143–151
Melillo JM, Terese (T.C.) R, Gary WY (eds) (2014) Climate change impacts in the United States: the third national climate assessment. U.S. Global Change Research Program. Washington DC, p 841. doi:10.7930/J0Z31WJ2
Mitsch W, Gosselink J (2000) Wetlands, 3rd edn. Wiley, New York
Mitsch WJ, Band LE, Cerco CF (2007) Everglades Landscape Model (ELM), Version 2.5
Moss RH et al. (2010) The next generation of scenarios for climate change research and assessment. Nature 463:747–756
NRC (2010) National Research Council: Progress Toward Restoring the Everglades: The Third Biennial Review. The National Academies Press, Washington DC, p 326. doi:10.17226/12988
NRC (2014a) National Research Council: Progress Toward Restoring the Everglades: The Fifth Biennial Review. The National Academies Press, Washington DC, p 302
NRC (2014b) “Chapter 5: Climate Change and Sea-Level Rise: Implications for Everglades Restoration”, in Progress Toward Restoring the Everglades: The Fifth Biennial Review, 2014. National Academies Press, Washington DC
Nungesser M, Saunders C, Coronado-Molina C, Obeysekera J, Johnson J, McVoy C, Benscoter B (2015) Potential effects of climate change on Florida’s Everglades. Environ Manag 55:824–835
Obeysekera J, Barnes J, Nungesser M (2015) Climate sensitivity runs and regional hydrologic modeling for predicting the response of the greater Florida Everglades ecosystem to climate change. Environ Manag 55:749–762
Obeysekera J, Irizarry M, Park J, Barnes J, Dessalegne T (2011) Climate change and its implications for water resources management in south Florida. Stoch Environ Res Risk Assess 25:495–516
Orem W, Fitz HC, Krabbenhoft D, Tate M, Gilmour C, Shafer M (2014) Modeling sulfate transport and distribution and methylmercury production associated with Aquifer Storage and Recovery implementation in the Everglades protection area. Sustain Water Qual Ecol 3:33–46
Orem W, Newman S, Osborne TZ, Reddy KR (2015) Projecting changes in Everglades soil biogeochemistry for carbon and other key elements, to possible 2060 climate and hydrologic scenarios. Environ Manag 55:776–798
Osborne T, Fitz H, Davis S,III (2017) Restoring the foundation of the Everglades ecosystem: assessment of edaphic responses to hydrologic restoration scenarios. Restoration Ecol. doi:10.1111/rec.12496
Osland MJ, Enwright NM, Day RH, Gabler CA, Stagg CL, Grace JB (2016) Beyond just sea‐level rise: considering macroclimatic drivers within coastal wetland vulnerability assessments to climate change. Glob Change Biol 22:1–11
Pearlstine LG, Pearlstine EV, Aumen NG (2010) A review of the ecological consequences and management implications of climate change for the Everglades. J North Am Benthol Soc 29:1510–1526
Peterson GD, Cumming GS, Carpenter SR (2003) Scenario planning: a tool for conservation in an uncertain world. Conserv Biol 17:358–366
Price RM, Swart PK (2006) Geochemical indicators of groundwater recharge in the surficial aquifer system, Everglades National Park, Florida, USA Geological Society of America Special Papers 404:251–266
Price RM, Swart PK, Fourqurean JW (2006) Coastal groundwater discharge–an additional source of phosphorus for the oligotrophic wetlands of the Everglades. Hydrobiologia 569:23–36
Richardson CJ, King RS, Qian SS, Vaithiyanathan P, Qualls RG, Stow CA (2007) Estimating ecological thresholds for phosphorus in the Everglades. Environ Sci Technol 41:8084–8091
Riscassi AL, Schaffranek RW (2004) Flow velocity, water temperature, and conductivity in Shark River Slough, Everglades National Park, Florida: June 2002–July 2003
Ross M, Meeder J, Sah J, Ruiz P, Telesnicki G (2000) The southeast saline Everglades revisited: 50 years of coastal vegetation change. J Veg Sci 11:101–112
Saha AK, Moses CS, Price RM, Engel V, Smith TJ, Anderson G (2012) A hydrological budget (2002–2008) for a large subtropical wetland ecosystem indicates marine groundwater discharge accompanies diminished freshwater flow. Estuaries Coasts 35:459–474
Saha AK et al. (2011) Sea level rise and South Florida coastal forests. Clim Change 107:81–108
Schallenberg M, Hall C, Burns C (2001) Climate change alters zooplankton community structure and biodiversity in coastal wetlands Report of Freshwater Ecology Group, University of Otago, Hamilton
SFRCCC (2015) South Florida Regional Climate Change Compact: A unified sea level rise projection for Southeast Florida. Southeast Florida Regional Climate Change Compact, Miami, FL
SFWMD (2005) South Florida Water Management District: Documentation of the South Florida Water Management Model. South Florida Water Management District, West Palm Beach, FL, Version 5.5
Sklar FH et al. (2005) The ecological-societal underpinnings of Everglades restoration Frontiers in Ecology and the Environment 3:161–169
Smith TJ, Anderson GH, Balentine K, Tiling G, Ward GA, Whelan KR (2009) Cumulative impacts of hurricanes on Florida mangrove ecosystems: sediment deposition, storm surges and vegetation. Wetlands 29:24–34
Smoak JM, Breithaupt JL, Smith TJ, Sanders CJ (2013) Sediment accretion and organic carbon burial relative to sea-level rise and storm events in two mangrove forests in Everglades National Park. Catena 104:58–66
Stabenau E, Engel V, Sadle J, Pearlstine L (2011) Sea-level rise: Observations, impacts, and proactive measures in Everglades National Park Park. Science 28:26–30
Tarboton K, Neidrauer C, Santee E, Needle J (1999) Regional hydrologic modeling for planning the management of South Florida’s water resources through 2050. Am Soc Agric Eng Paper, Paper Number 992062
Troxler T (2017a) Water Quality Data (Extensive) from the Taylor Slough, Everglades National Park (FCE), from April 1996 to Present. Environmental Data Initiative. doi:10.6073/pasta/8e3c2cc6502bf658dbf6f42be1822f28. Accessed 8 Jan 2017
Troxler T (2017b) Water Quality Data (Extensive) from the Taylor Slough, Everglades National Park (FCE), South Florida from July 1999 to Present. Environmental Data Initiative. doi:10.6073/pasta/241bc3e309edcb2aaaa4328d99bf0680. Accessed 8 Jan 2017
Troxler T, Childers D (2008) Water Quality Data (Extensive) from the Taylor Slough, just outside Everglades National Park (FCE), from August 1998 to December 2006. Environmental Data Initiative.doi:10.6073/pasta/3a668167984681792eb010cd334d19af. Accessed 8 Jan 2017
USACE (1999) U.S. Army Corps of Engineers and South Florida Water Management District: The Central and Southern Florida Flood Control Project Comprehensive Review Study Final Integrated Feasibility Report and Programmatic Environmental Impact Statement (PEIS), West Palm Beach, FL
van der Valk AG, Volin JC, Wetzel PR (2015) Predicted changes in interannual water-level fluctuations due to climate change and its implications for the vegetation of the Florida Everglades. Environ Manag 55:799–806
Venice_System (1959) The Venice System for the Classification of Marine Waters According to Salinity. Arch Oceanogr Limnol 9:243–245
Wanless HR, Parkinson RW, Tedesco LP (1994) Sea level control on stability of Everglades wetlands Everglades. the Ecosystem and Its Restoration St Lucie Press, Delray Beach, FL, p 199–223
Wdowinski S, Bray R, Kirtman BP, Wu Z (2016) Increasing flooding hazard in coastal communities due to rising sea level. Case Study Miami Beach, Fla Ocean & Coast Manag 126:1–8
West JM et al. (2009) US natural resources and climate change: concepts and approaches for management adaptation. Environ Manag 44:1001–1021
Zapata‐Rios X, Rivero RG, Naja GM, Goovaerts P (2012) Spatial and temporal phosphorus distribution changes in a large wetland ecosystem Water Resour Res 48, Article number: W09512
Zhang K (2011) Analysis of non-linear inundation from sea-level rise using LIDAR data: a case study for South Florida. Clim Change 106:537–565
Acknowledgements
This material is based upon work supported by the National Science Foundation through the Florida Coastal Everglades Long-Term Ecological Research program under Cooperative Agreements No. DEB-1237517, Grant No. DBI-0620409, and Grant No. DEB-9910514. This is contribution number 840 from the Southeast Environmental Research Center at Florida International University. We would like to thank Dr. Jayantha Obeysekera and Jenifer Barnes of Hydrologic & Environmental Systems Modeling, South Florida Water Management District, for providing results of simulation runs of the South Florida Water Management Model. The performance assessments of our Everglades Landscape Model would not have been possible without the long-term monitoring data sets provided by the Florida Coastal Everglades Long Term Everglades Research Program. In particular we appreciate Dr. Evelyn Gaiser and Dr. Daniel Childers for sharing their extensive long term water quality data sets from Shark River Slough (Gaiser and Childers 2016). We also appreciate Dr. Tiffany Troxler for sharing her extensive long term water quality data sets from Taylor Slough (Troxler 2017a; Troxler 2017b; Troxler and Childers 2008).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no competing interests.
Electronic Supplementary Material
Rights and permissions
About this article

Cite this article
Flower, H., Rains, M. & Fitz, C. Visioning the Future: Scenarios Modeling of the Florida Coastal Everglades. Environmental Management 60, 989–1009 (2017). https://doi.org/10.1007/s00267-017-0916-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-017-0916-2