
Abstract
Ecotones, the narrow transition zones between extensive ecological systems, may serve as sensitive indicators of climate change because they harbor species that are often near the limit of their physical and competitive tolerances. We investigated the ecotone between salt marsh and adjacent upland at Elkhorn Slough, an estuary in California, USA. Over a period of 10 years, we monitored movement of the ecotone–upland boundary, plant community structure, and physical factors likely to drive ecotone response. At three undiked sites, the ecotone boundary migrated about 1 m landward, representing a substantial shift for a transition zone that is only a few meters wide. Analysis of potential correlates of this upward migration suggests that it was driven by increased tidal inundation. Mean sea level did not increase during our study, but inundation at high elevations did. While the ecotone boundary responded dynamically to interannual changes in inundation at these undiked sites, the plant community structure of the ecotone remained stable. At two diked sites, we observed contrasting patterns. At one site, the ecotone boundary migrated seaward, while at the other, it showed no consistent trend. Diking appears to eliminate natural sensitivity of the ecotone boundary to interannual variation in oceanic and atmospheric drivers, with local factors (management of water control structures) outweighing regional ones. Our study shows that the marsh–upland ecotone migrated rapidly in response to environmental change while maintaining stable plant community structure. Such resilience, stability, and rapid response time suggest that the marsh–upland ecotone can serve as a sensitive indicator of climate change.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Ecotones are transition zones between adjacent ecological systems (Risser 1995). Typically, they are narrow zones with steep environmental gradients located between extensive systems with more consistent environmental conditions. Ecotone boundaries may be particularly responsive to environmental changes because species are living near the edge of their tolerances (Peters et al. 2006, Yarrow and Martin 2007). Thus, ecotones may serve as sensitive indicators of global climate change (Risser 1995). Historical ecology analyses have linked past climate change to changes in boundaries of ecosystems (e.g., Crumley 1993). Ecotones could likewise serve as sensitive monitoring indicators of contemporary and future, accelerated climate change.
Modeling suggests that only ecotones with high resiliency and stability, and with quick response rates will track rapid environmental change. For instance, upward creep of tree lines along altitudinal and latitudinal gradients in response to global warming may occur too slowly or inconsistently, as a result of increased fire or flood frequency, to rapidly track changes in climatic conditions (Noble 1993, Kupfer and Cairns 1996). It has been hypothesized that wetland margins, in contrast, may prove to be more sensitive indicators of climate change (Noble 1993). Coastal estuaries migrate landward or seaward in response to changes in sea level over long time periods. Much recent attention has focused on ability of estuarine salt marshes to track predicted sea level increases (e.g., Morris et al. 2002; Kirwan et al. 2010). Estuaries can encompass multiple ecotones (Elliot and Whitfield 2011). We focus on one estuarine ecotone: the landward boundary of the salt marsh. The goal of our study was to determine whether the salt marsh–upland ecotone has potential to serve as a sensitive indicator of climate change, specifically sea level rise.
The salt marsh–upland transition zone in California (Fig. 1) is a narrow ecotone that harbors a unique plant community, supports wildlife, and plays an important role in nutrient uptake (James and Zedler 2000; Traut 2005; Wasson and Woolfolk 2011). The boundaries of this ecotone are probably set largely by edaphic factors such as soil salinity and moisture (Callaway et al. 1990; James and Zedler 2000), which directly affect distribution of marsh and upland species through physical tolerance limits and indirectly affect them through altered strength of competition (Cui et al. 2011). Soil salinity and moisture in the ecotone can be affected by tidal inundation and by precipitation. Both inundation and precipitation can be influenced by interannual variation in regional atmospheric conditions. Human management of tidal exchange through water control structures can also affect inundation time and influence ecotone structure (Wasson and Woolfolk 2011).

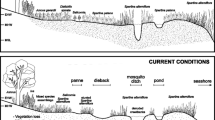
Salt marsh–upland ecotone. This transition zone is a narrow band between 100 % vegetated cover by pickleweed (reddish vegetation to the right in this photo) and 100 % cover by upland vegetation (larger shrubs and grasses to the left in this photo)
The objectives of our study were to determine (1) whether the boundaries of the salt marsh–upland ecotone are temporally dynamic on short timescales, (2) whether artificial tidal restriction affects temporal dynamics, (3) whether temporal dynamics of the ecotone are correlated to trends in climatic or oceanographic factors, and (4) whether similar plant communities persist over time, demonstrating resilience of the ecotonal plant community to climate change. Movement of the ecotone boundary in response to short-term variation in local or regional drivers, while maintaining stable plant community structure, would demonstrate the potential of this ecotone to serve as a useful indicator for climate change.
Methods
Study System and Sampling Sites
We conducted this study in the marsh–upland ecotone at Elkhorn Slough, a 1,200-ha estuary in central California, USA. This estuary has one of the most extensive salt marshes in the region, after San Francisco Bay. The estuary receives only limited freshwater inputs, and water salinity in undiked regions is usually close to salinity in adjacent marine habitats. Tides are semidiurnal and ebb-dominated, with a maximum tidal range of about 2.5 m. The region has a Mediterranean climate, with all significant rainfall occurring between October and May. More background on the estuary can be found in Caffrey (2002).
Salt marsh occurs between about Mean High Water (MHW) and Mean Higher High Water (MHHW) at Elkhorn Slough. One marsh dominant, pickeweed (Salicornia pacifica), accounts for almost all the vegetated cover in this zone; there is no cord grass (Spartina spp.) at this estuary. Between MHHW and the reach of the very highest tides lies a transition zone between the marsh and the adjacent uplands, the salt marsh–upland ecotone (Fig. 1). This is a narrow band, typically only a few meters wide (Wasson and Woolfolk 2011), while the marsh plain seaward of it stretches for hundreds of meters. The vertical elevational range of the ecotone is from about 1.6 to 2.2 m NAVD88 at this estuary. Various native marsh plants are limited to this zone; these are “ecotone specialists.” The most common ecotone specialists at Elkhorn Slough are alkali heath (Frankenia salina), salt grass (Distichlis spicata), fleshy jaumea (Jaumea carnosa), and spearscale (Atriplex triangularis) (Wasson and Woolfolk 2011). Both the marsh-dominant and upland plants are also represented in this transition zone. At Elkhorn Slough, grasslands and scrub typically occur above the salt marshes. These habitats are highly invaded by non-native species, many of which also occur in the ecotone (Wasson and Woolfolk 2011).
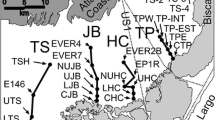
We established a long-term monitoring program to track the movement of the ecotone boundary, selecting five sites at which to establish permanent transects (Fig. 2). Replicate vegetation transects across environmental gradients may provide a particularly powerful means to monitor species migrations at landscape scales (Stohlgren et al. 2000). The sites were chosen for ease of access and to span representative conditions. Three sites were chosen in undiked portions of the estuary, exposed to the full 2.5 m maximum tidal range. These were located in the mid- to upper estuary because the ecotone in the lower estuary has been highly altered (with a harbor, salt ponds, or berms adjacent to the marsh rather than natural hillsides). Two of the sites are along the main channel in sites that have never been diked (U1–2). One of the sites (U3) was diked for many decades and then restored to full tidal exchange in 1982. During the diked period, the area formerly occupied by marsh subsided, and when tidal exchange was returned in the 1980s, mudflats replaced former marshes because these areas were now 50–150 cm below MHW on average (unpublished LIDAR data). The marsh dominant and the ecotone are now compressed into a narrow zone of fringing marsh along the steep hillsides at this site.

Location of monitoring sites. Ecotone monitoring occurred at three undiked sites (U1–3) and two diked sites (D1–2). Water level and precipitation monitoring occurred at a paired water quality and weather station (WS)
The two remaining sites were selected in diked portions of the estuary. Site D1 is an extensive marsh at the head of the estuary, which receives seasonal freshwater input from two creeks and agricultural runoff. Tidal exchange is excluded with one-way tide gates under a road that serves as a dike; the tide gates allow for freshwater to drain out of the wetland but prevent entry of saltwater. The tide gates periodically leak when they are propped open by debris and were broken continuously for about 5 years (from a 1989 earthquake until they were repaired in 1995). Thus, while the formerly brackish wetland has been managed for many decades as a freshwater impoundment to protect upstream agriculture and residences, it occasionally receives tidal inundation. Site D2 was also managed to exclude tidal exchange. It receives little fresh water and is separated from the tidal portions of the estuary by berms. However, during our study period, in 2003, a breach occurred in one of the berms and gradually increased in size over subsequent years. This increased the extent of standing water in the wetland, and while no surface inundation of the ecotone resulted, the marsh soils appear to have become more waterlogged due to greater proximity to standing water.
Field Monitoring Design
At each site, we established five permanent belt transects in 2001. The transects were 50 cm wide and located haphazardly at each site, at least 10 m apart, but all within a 500-m stretch of shoreline. Each transect was run perpendicular to the shoreline (Fig. 3) and marked with a PVC stake placed near the landward boundary of the ecotone. Measurements of the ecotone boundary locations in this and all subsequent years were taken relative to this PVC marker. By our definition, the ecotone encompasses the zone between 100 % of vegetated cover by the marsh dominant and 100 % cover by upland plants. We located the seaward boundary of the ecotone within each transect, defined as the most landward location where 100 % of the vegetative cover consists of the marsh plain dominant, S. pacifica. We located the landward boundary of the ecotone within each transect, defined as the most seaward location where 100 % of the vegetative cover consists of upland plants. We noted the identity of the marsh plant and upland plant at the landward boundary of the ecotone and surveyed the transects to generate a species list of all plants with >10 % cover within the transects (between the landward and seaward boundaries of the ecotone).

Ecotone sampling design. The ecotone is the zone between 100 % pickleweed cover and 100 % upland vegetation cover, where ecotone specialists occur along with pickleweed and upland plants. A PVC marker was placed in the ecotone in 2001. X In every monitoring survey, the horizontal distance from this marker to the current landward boundary of the ecotone was measured with a transect tape (in this example, the distance is to the 2008 boundary). Y The vertical elevational difference between the 2001 and 2008 boundaries was measured once (in 2010) by relocating these boundaries relative to the PVC marker
We revisited these transects in 2002, 2003, 2008, 2010, and 2011, repeating the above assessments. These regular measurements were taken with a transect tape running from the permanent PVC marker and thus represent horizontal positions in the landscape, not elevations (X in Fig. 3). Sites were visited in September in every year, towards the end of the marsh plant growing season.
Elevational Surveys
To complement our regular transect tape measurements of distance of the ecotone boundary from the permanent marker, in 2010, we also did a one-time assessment of vertical elevational change over time. We relocated the position where the ecotone boundary had been in 2001 and 2008 using transect tape measurements from the permanent marker. These years were chosen because they represent the two most contrasting years at the undiked sites, with the greatest change in horizontal distance in boundary location. We measured the elevation of each of these boundaries relative to a known benchmark using a surveyor's autolevel. The orthometric height of each benchmark was established using GPS observations with a Trimble 5800 RTK, with vertical elevational precision of 2.5 cm. Survey data was postprocessed using the National Oceanic and Atmospheric Administration's Online Positioning User Service (OPUS) and the GEOID09 model. We then calculated the vertical elevation difference between the landward boundary of the ecotone in 2001 and 2008 (Y in Fig. 3).
Analysis of Monitoring and Elevational Data
To characterize temporal changes in the location of the landward boundary of the ecotone at each long-term monitoring site, we conducted repeated measures ANOVA separately for each of the five sites, using the five transects per site as replicates. We used Fisher's PLSD (alpha = 0.05) to detect significant differences between pairs of individual years for the horizontal location of the boundary, for which we had data from six separate years. For changes in vertical elevation, we only had data from 2 years (2001 and 2008) and thus, no post hoc tests were necessary.
To determine whether the ecotone plant community composition was stable in the face of temporal changes in location, we examined plant communities in the ecotone transect at the undiked sites, comparing data from 2001 vs. 2008, where the largest observed divergence in ecotone boundaries occurred. We analyzed presence/absence data from the species lists obtained from the 15 undiked transects using the program Primer v. 6 (Clarke and Gorley 2006). To graphically compare patterns of species composition between 2001 and 2008, we created nonmetric multidimensional scaling (nMDS) plots based on a Bray–Curtis resemblance matrix. In such plots, samples with similar community structure cluster together, while dissimilar samples are located far from each other. Analysis of similarities (ANOSIM) was used to determine whether there was significant dissimilarity in community structure between years.
Examination of Potential Causes of Landward Ecotone Migration at Undiked Sites
The analysis of the location of the ecotone boundary revealed a very consistent pattern at all three undiked sites: the landward boundary of the ecotone migrated landward in the latter three monitoring years. To enable comparisons with potential causal factors, we conducted a t test on the horizontal location of the landward boundary of the ecotone, averaging across all 15 undiked transects, using year as replicate, and monitoring period (early = 2001–2003; late = 2008–2011) as factor. We then conducted similar t tests by period on three physical factors: mean water level, inundation time at the elevation of the most landward portion of the ecotone, and precipitation. Since early vs. late monitoring years differed for the ecotone boundary, any causal factor should display a similar difference between periods. In addition to these univariate analyses of individual factors, we conducted simple linear regressions between location of the landward boundary of the ecotone in each year vs. each of the physical factors.
Data on water levels were obtained from a long-term water monitoring station (WS in Fig. 2) that is part of the National Estuarine Research Reserve System-wide Monitoring Program (NERR SWMP). A sonde (Yellow Springs Instruments) was continuously deployed at a fixed location and logged depth data every 15 minutes using a pressure sensor. The depth data were converted to water levels using NAVD88 vertical datum after surveying a local bench mark, the orthometric height of which was established with static GPS observation using the GEOID09 model. MLLW at this water monitoring station is about 0.04 m higher than 0.0 NAVD88, a scale difference we used for converting between the two measurement types. We analyzed the data to determine the mean water level and the total hours of inundation at an elevation of 2.1 m NAVD88, which corresponds to location of the most landward portion of the ecotone in 2001. We averaged the water level data and summed the 15-min intervals with water levels higher than 2.1 to obtain total inundation time for the period for each elevation. (Inundation duration was highly correlated to inundation frequency because inundation events at this elevation were 15–135 min in duration, occurring right around the higher high tide of the day on a spring tide week. So number of hours of inundation per year and number of days with such events per year were similar, but the former showed somewhat more variation and thus was used for our analysis.) We also examined precipitation data from a meteorological station at the same site, which was also part of the NERR SWMP network (more information on data collection is available at http://cdmo.baruch.sc.edu). For all of these physical variables, we summed or averaged values for the period 2 years prior to each field monitoring survey. This was chosen as a relevant ecological time frame for changes in soil salinity or moisture to translate into changes in plant distribution; changes might not be immediate for the marsh and ecotonal plants, which include various perennial species.
In addition to examining potential physical correlates, we explored the possibility that changes in a dominant plant species might be responsible for boundary changes, for instance with a particular marsh plant undergoing a landward expansion or a particular upland plant undergoing retreat. We examined our data on identity of the marsh plant and dominant upland plant located at the landward ecotone boundary, conducting ANOVA to determine whether plant identity affected rate of movement of the ecotone boundary. We also compared marsh species present at the boundary in 2001 vs. 2008 to determine whether colonization of the zone above the 2001 ecotone occurred by the same species originally present or by different species.
Results
Temporal Dynamics from Field Surveys
Change in Location of Ecotone Boundary
Analysis of the field measurements of horizontal locations of ecotone boundaries revealed that at the three undiked sites, temporal dynamics at the landward boundary of the ecotone were remarkably consistent (Fig. 4a–c). The most pronounced difference, significant at all three sites in repeated measures ANOVA, was the landward migration of the boundary in the later three monitoring years vs. the early three years. The patterns at the three undiked sites are also similar in their details: the boundary was furthest seaward at all sites in 2001, moved gradually landward in 2002 and 2003, migrated dramatically landward between 2003 and 2008 and then remained in a similar location in 2010 and 2011.

Location of the landward ecotone boundary over time. P values listed refer to the overall significance of the repeated measures ANOVA conducted. Small letters indicate which years were significantly different in Fisher's PLSD post hoc tests. Bars show standard errors. Each graph represents a different site (abbreviations as in Fig. 1): a U1, b U2, c U3, d D1, e D2. At sites U2 and U3, the ecotone boundary was located at the marker in 2001; thus, a value of zero is shown. Positive values on the y-axis indicate landward migration of the upland boundary
The above data on horizontal locations obtained by transect tape measurements from the permanent marker made in each monitoring period revealed an average landward migration of about 1 m at these three undiked sites over the monitoring period. Vertical elevation data obtained in 2010, relocating the boundaries from the years of greatest contrast (2001 vs. 2008), revealed that the landward boundary migrated upward on average 19 cm across the three undiked sites between 2001 and 2008, from an average of 2.20 to 2.39 m NAVD88. This increase in elevation over time was statistically significant at all three undiked sites in repeated measures ANOVA (U1: p = 0.02, U2: p = <0.0001, U3: p = 0.01).
The two diked sites displayed contrasting patterns, as assessed by the horizontal field measurements. At D1, repeated measures ANOVA revealed that the ecotone boundary was significantly lower in most of the later years than the earlier years (Fig. 4d). At this site with gradual slope, the ecotone boundary had migrated seaward about 4 m in our horizontal transect tape measurements. The vertical elevation data revealed that this corresponded to an 11-cm decrease in elevation. Repeated measures ANOVA showed that the boundary had moved down significantly (p = 0.04) between 2001 and 2008, from 0.88 to 0.77 m NAVD88.
At site D2, repeated measures ANOVA of the horizontal ecotone boundary showed no significant temporal patterns (Fig. 4e). While there was a gradual seaward migration in the first years, this was reversed by 2008, the first measurement after the breach of the dike occurred. Vertical elevation also showed no significant change in repeated measures ANOVA.
Stability of Plant Community Structure
Nonmetric multidimensional scaling plots of plant community composition at undiked sites over the monitoring period revealed considerable overlap between plant communities in 2001 vs. 2008, the years of greatest contrast in ecotone boundary location. The associated statistical procedure, ANOSIM, confirmed that plant communities in 2001 vs. 2008 were not significantly different and, indeed, were virtually indistinguishable (global R = −0.01, p = 0.54).
Examination of Potential Correlates of Ecotone Boundary Movement
Our examination of the correlation of tidal inundation, water level, precipitation, and boundary plant species with ecotone boundary movement at undiked sites revealed that only tidal inundation showed patterns consistent with boundary movement.
Physical Factors: Inundation Time, Water Level, and Precipitation
Figure 5 summarized the results of t tests comparing early vs. late monitoring periods for the location of the ecotone boundary and three potential physical drivers. The ecotone boundary migrated landward significantly between periods (Fig. 5a), so a physical driver causing this pattern should also show significant differences between these periods.

Comparison of ecotone boundary location and three physical factors in early vs. late monitoring years. P values listed refer to the significance of the comparison in a t test. Bars show standard errors. The replicate was sampling year (2001, 2002, and 2003 for the early period, and 2008, 2010, and 2011 for the late period). For b–d, environmental data from the 24 months prior to the September ecotone field sampling in each year was used
The number of hours of tidal inundation at 2.1 m NAVD88 (corresponding an elevation near the most landward portion of the ecotone) in the 2 years prior to our field sampling showed the most similar temporal pattern to the top of the ecotone of the factors we examined (Fig. 5b). A t test indicated that there were significantly fewer hours of tidal inundation at this elevation in 2001–2003 (29 h on average) than in 2008–2011 (68 h on average). Mean water level increased slightly, from an average of 97.2 to 99.2 cm above MLLW between the periods, a difference that was not statistically significant (Fig. 5c) and seems insufficient to account for the nearly 20-cm vertical elevation change in the ecotone boundary. Precipitation also increased between periods, from an average of 65 to 88 cm (per 24 months), but this difference was not statistically significant (Fig. 5d).
A simple regression of location of ecotone boundary vs. inundation time at 2.1 m NAVD88 revealed a strong relationship that was significant despite the small sample size (r 2 = 0.81, p = 0.015). In contrast, simple regressions with water level or precipitation did not reveal any strong relationships (r 2 < 0.5, p > 0.1 in both cases).
Biological Factors: Identity of Boundary Plant Species
The identity of the marsh plant at the ecotone–upland boundary had no effect on rate of movement of the boundary between 2001 and 2008, the most extreme contrast. Five marsh plants were found at the top of the ecotone in the transects: the marsh dominant S. pacifica and four ecotone specialists (A. triangularis, D. spicata, F. salina, and J. carnosa). Analysis of amount of boundary movement for each of these five species showed no significant difference, nor was there a difference when all ecotone specialists were lumped into one category and compared to the marsh dominant. At 14 out of 15 of the transects in undiked areas, the species present at the boundary in 2001 was still present in 2008, so the boundary shift was not linked to colonization by different species.
The identity of the dominant upland plant above the ecotone also had no effect on movement of the boundary. A variety of upland plant species were found to be dominant at the top of the ecotone; the most common were the native shrub Baccharis pilularis and grass Leymus triticoides, and various non-native forbs (Brassica nigra/rapa, Carduus pycnocephalus, Conium maculatum, and Raphanus raphanistrum/sativus). Neither a comparison of rate of boundary movement by individual species nor by lumped category (native vs. non-native) had any significant effect. These results indicate that the boundary movement occurred as a general landward migration of the marsh plant community and retreat of the upland plant community, rather than resulting from changes in particular marsh or upland species.
Discussion
Ecotone Boundaries are Very Dynamic but Ecotone Plant Communities are Stable
The results of this 10-year monitoring study indicate that the boundary of the high marsh–upland ecotone is capable of rapid movement. Specifically, the marsh–upland boundary at undiked sites in Elkhorn Slough showed rapid upward migration of this boundary by almost 20 cm in vertical elevation over just a few years, corresponding to a landward migration of about 100 cm as measured across the marsh landscape. Given that at Elkhorn Slough, the total elevational range of the ecotone is about 60 cm and the landscape width of the ecotone zone is only 200–500 cm, this migration represents a very significant shift in location. Our finding supports the hypothesis that ecotones may be particularly responsive to environmental change because ecotonal species are typically at the limits of their tolerance (Peters et al. 2006).
While the location of the narrow transitional band of ecotone vegetation was very dynamic between years, the plant community composition within the zone was stable. The rate of movement did not correlate with the identity of the marsh plant at the ecotone boundary or the dominant upland plant just above it or the dominant marsh plant just above it. This suggests that the boundary movement was due to wholesale landward migration of the marsh community, not the result of a particular marsh or ecotonal species rapidly colonizing landward or a specific upland plant retreating. This stability of the plant community composition in the face of environmental change indicates that the marsh–upland ecotone community is highly resilient. Ecologically, the resilience of the salt marsh–upland ecotone plant community is important because ecotonal plant species can play an important role in nitrogen uptake (Page 1995; Traut 2005) and wildlife support (Zedler et al. 2001). This California high marsh ecotone appears to be much more resilient than the high marsh community of Cape Cod, Massachusetts, which has also undergone landward migration but suffered significant loss of habitat extent (Smith 2009).
Increased Inundation Correlates with Ecotone Migration
In a classic paper in landscape ecology, Wiens et al. (1985) stressed that “in order to begin to understand landscape-level patterns and processes, we must know the details of ‘boundary dynamics’—what determines why a boundary is located where it is.” The interannual movements of the marsh–upland boundary occurred with remarkable consistency across three separate undiked sites at Elkhorn Slough (Fig. 4a–c), indicating that movement was a response to changes in estuarywide drivers, not site-specific factors. The strongest correlation we found with the landward migration of the marsh–upland boundary was an increase in inundation time at the elevation corresponding to this boundary in the late vs. early years of our monitoring (Fig. 5b). Since the ecotone–upland boundary occurs within the reach of the very highest tides, an upward shift in the highest tides could shift the boundary of this zone upward. Inundation of the zone just above the boundary likely led to mortality of upland plants, and marsh plants were subsequently able to colonize this newly available space landward of the former boundary through vegetative growth of existing individuals near the boundary or germination of seeds.
The mechanism behind the increased inundation at high elevations is unclear but could be related to climate change. To obtain a longer time series, we examined all years for which we had a complete record from our water level monitoring station (1996–2010). Hours of inundation at 2.1 m NAVD88 increased significantly over this period when examined as a simple regression (Fig. 6). However, mean water level did not increase in the estuary over this period in a simple regression (r 2 < 0.001, P = 0.94). This lack of change in mean sea level is not unusual for California. Compared to rates of global mean sea level rise, rates of relative sea level rise along the US West Coast are low, averaging only 1.4 mm/year south of Cape Mendocino for the past six to ten decades (Committee on Sea Level Rise in California, Oregon, and Washington 2012). Likewise, contrasts between inundation patterns at different elevations are common in tidal records; for instance, MHHW in nearby San Francisco Bay has increased about 19 % faster than mean sea level in past decades (Flick et al. 2003). So, the increased inundation at high elevations observed at Elkhorn Slough may be related to eustatic sea level rise or to some more local hydrodynamic phenomenon. In any case, the ecotone has migrated rapidly in response to the increased inundation. This migration may increase in the future. Although the West Coast of the US has not generally experienced increased mean sea levels in recent decades, changing wind stress patterns may result in accelerated sea level rise rates in the North Pacific Ocean (Bromirski et al. 2011).

Temporal patterns of inundation at elevation at landward margin of the estuary. Hours of inundation at 2.1 m NAVD88 have increased over the monitoring period (1996–2010)
Effects of Diking
Ecotones and ecological boundaries are increasingly being recognized as playing important functional roles in human-modified landscapes (Yarrow and Marin 2007). Diking appears to eliminate the natural sensitivity of the ecotone boundary to interannual variation in oceanic and atmospheric drivers. While the upper ecotone boundary at the three undiked sites showed very consistent interannual patterns that could be related to regional oceanic or atmospheric drivers, the boundary dynamics at two diked sites appeared to be largely driven by the conditions imposed by the water control structures at these sites. Contrasts between these two sites illustrate the strong effect that changes in local water management can have on the direction and magnitude of the ecotone boundary movement.
Artificial tidal restriction can result in seaward migration of the upland and ecotone communities, displacing the marsh community and leading to subsequent net loss of wetland habitat (Wasson and Woolfolk 2011). At one diked site (D1), tidal water has largely been excluded with tide gates since the 1950s. Saline water flooded this marsh in the early 1990s (during a period of tide gate failure), but since then, most tides have been excluded and soil salinity has decreased relative to tidal areas (Wasson and Woolfolk, unpublished data). At this site, the top of the ecotone migrated seaward very significantly over the period of our monitoring, with the boundary in 2010 almost 5 m seaward from its location in 2001 (Fig. 4d). Upland weeds are now occupying an area that was dominated by marsh plants when the monitoring began. It is not clear whether this extremely rapid seaward migration is temporary, perhaps an adjustment to the repair of the tide gates that decreased inundation after a period of greater flooding, or whether this is a consistent trajectory continuing from the initial diking in the 1950s. In terms of vertical elevation, the top of the ecotone at D1 was at 0.77 m NAVD88 in 2008, much lower than the average of 2.39 m for undiked sites. Our elevation data suggest that the ecotone is currently occupying the historic marsh plain at this site (Wasson and Woolfolk 2011). Tidal restriction thus can lead to opposite patterns of marsh migration than observed at natural sites, preventing diked marshes from keeping pace with sea level rise. At some future point, the dikes may be breached by rising sea levels, but the marsh plain and ecotone will be occupying too low an elevation to sustain vegetation. It has been established that in many estuaries, a return of tidal flooding and sediment supply can lead to increased trapping and organic matter storage in tidal wetlands, allowing for accretion and marsh building (Burdick and Roman 2012). However, this feedback model has its limits. Low elevations, limited estuarine-suspended sediment supply, and erosion of deposited estuarine mud can limit marsh reestablishment in formerly diked areas (Williams and Orr 2002). In such cases, resilience to sea level rise has been lost. Indeed, one of the greatest challenges of tidal marsh restoration already in the past decades is the elevation deficit relative to sea level, when dikes are removed (Van Dyke and Wasson 2005); this problem will only become exacerbated with accelerated sea level rise in the future.
Sufficient tidal inundation at sites with water control structures can prevent seaward migration of upland plants and consequent loss of wetland habitat. At the second diked site we examined (D2), the ecotone was initially moving seaward (Fig. 4e). However, midway through our monitoring period, in 2003, a berm broke, increasing tidal inundation of this wetland. This change appeared to reverse the trends, with the ecotone boundary shifting landward again. Our results are consistent with landward migration of salt marsh species into areas previously dominated by pasture weeds, as a result of restoration of more natural tidal exchange (Streever and Genders 1997).
Salt Marsh Upland Ecotone as an Indicator of Global Climate Change
The salt marsh–upland ecotone may be much more responsive on short timescales to environmental changes than is the rest of the salt marsh. Much attention has been focused on response of the salt marsh plain to predicted relative sea level rise. Modeling suggests that, given sufficient sediment loads, salt marshes may be able to maintain their current distribution in estuaries due to increased sediment accretion rates with higher inundation times (Morris et al. 2002; Kirwan et al. 2010). However, at the landward margin, inundation times are so brief that this sort of equilibrium may not be possible. Furthermore, upland species are not able to respond because they do not accrete by means of the same processes as marsh plants. The marsh–upland ecotone may be more likely to respond rapidly by migration rather than sediment accretion to track sea level. Empirical data on rapid changes in location of the marsh-upland boundary can thus complement modeling of broader salt marsh response to increased relative sea level rise and can inform understanding of migration potential of salt marsh habitat. Our work supports the suggestion (Stohlgren et al.2000) that replicate transects along environmental gradients can prove a powerful approach for detecting plant migrations.
It has been proposed that “a significant research challenge for the future involves testing the idea of whether ecotones, or characteristics of ecotones, are valuable as indicators of change in the global environment” (Risser 1990). In order for ecotones to serve as sensitive indicators of climate change, they should show high resilience, stability, and rapid movement in response to environmental changes (Noble 1993). Our results confirm the hypothesis (Noble 1993) that wetland margins meet these conditions and serve as valuable indicators for climate change. The responsiveness of the marsh–upland boundary to increased tidal inundation detected in our study suggests that this ecotone will be a sensitive indicator of ecological responses to increased eustatic sea level rise predicted as a part of global climate change.
References
Bromirski, P.D., A.J. Miller, R.E. Flick, and G. Auad. 2011. Dynamical suppression of sea level rise along the Pacific coast of North America: indications for imminent acceleration. J Geophys Res 116: 1–13.
Burdick, D.M., and C.T. Roman. 2012. Salt marsh responses to tidal restriction and restoration: a summary of experiences. In Tidal marsh restoration, ed. C.T. Roman and D.M. Burdick, 373–382. Washington, DC: Island Press.
Caffrey, J., M. Brown, W.B. Tyler, and M. Silberstein. 2002. Changes in a California estuary: a profile of Elkhorn Slough. Moss Landing, CA: Elkhorn Slough Foundation.
Callaway, R.M., S. Jones, W.R. Ferren, and A. Parikh. 1990. Ecology of a Mediterranean-climate estuarine wetland at Carpinteria, California—plant distributions and soil-salinity in the upper marsh. Can J Bot 68(5): 1139–1146.
Clarke, K.R. and Gorley R.N. 2006. PRIMER (Plymouth Routines In Multivariate Ecological Research) v. 6. Plymouth: PRIMER-E Ltd.
Committee on Sea Level Rise in California, Oregon, and Washington, Board on Earth Sciences and Resources, Ocean Studies Board, Division on Earth and Life Studies, National Research Council. 2012. Sea-level rise for the coasts of California, Oregon, and Washington: past, present, and future. Washington, DC: The National Academies Press.
Crumley, C.L. 1993. Analyzing historic ecotonal shifts. Ecol Appl 3(3): 377–384.
Cui, B.S., Q.A. He, and Y.A. An. 2011. Community structure and abiotic determinants of salt marsh plant zonation vary across topographic gradients. Estuaries and Coasts 34(3): 459–469.
Elliott, M., and A.K. Whitfield. 2011. Challenging paradigms in estuarine ecology and management. Estuarine Coastal Shelf Sci 94: 306–314.
Flick, R.E., J.F. Murray, and L.C. Ewing. 2003. Trends in United States tidal datum statistics and tide range. J Waterw Port Coast Ocean Eng 129(4): 155–164.
James, M.L., and J. Zedler. 2000. Dynamics of wetland and upland subshrubs at the salt marsh–coastal sage scrub ecotone. Am Midl Nat 143(2): 298–311.
Kirwan, M.L., G.R. Guntenspergen, A. D'Alpaos, J.T. Morris, S.M. Mudd, and S. Temmerman. 2010. Limits on the adaptability of coastal marshes to rising sea level. Geophys Res Lett 37: 1–5.
Kupfer, J.A., and D.M. Cairns. 1996. The suitability of montane ecotones as indicators of global climatic change. Prog Phys Geogr 20(3): 253–272.
Morris, J.T., P.V. Sundareshwar, C.T. Nietch, B. Kjerfve, and D.R. Cahoon. 2002. Responses of coastal wetlands to rising sea level. Ecology 83(10): 2869–2877.
Noble, I.R. 1993. A model of the responses of ecotones to climate change. Ecol Appl 3(3): 396–403.
Page, H.M. 1995. Variation in the natural abundance of N-15 in the halophyte, Salicornia virginica, associated with groundwater subsidies of nitrogen in a southern California salt-marsh. Oecologia 104(2): 181–188.
Peters, D.P.C., J.R. Gosz, W.T. Pockman, E.E. Small, R.R. Parmenter, S.L. Collins, and E. Muldavin. 2006. Integrating patch and boundary dynamics to understand and predict biotic transitions at multiple scales. Landsc Ecol 21(1): 19–33.
Risser, P.G. 1990. The ecological importance of land–water ecotones. In The ecology and management of aquatic–terrestrial ecotones, ed. R.J. Haiman and H. Decamps, 7–21. Paris: UNESCO.
Risser, P.G. 1995. The status of the science examining ecotones—a dynamic aspect of landscape is the area of steep gradients between more homogeneous vegetation associations. BioScience 45(5): 318–325.
Smith, S.M. 2009. Multi-decadal changes in salt marshes of Cape Cod, MA: photographic analyses of vegetation loss, species shifts, and geomorphic change. Northeast Nat 16(2): 183–208.
Stohlgren, T.J., A.J. Owen, and M. Lee. 2000. Monitoring shifts in plant diversity in response to climate change: a method for landscapes. Biodivers Conserv 9(1): 65–86.
Streever, W.J., and A.J. Genders. 1997. Effect of improved tidal flushing and competitive interactions at the boundary between salt marsh and pasture. Estuaries 20(4): 807–818.
Traut, B.H. 2005. The role of coastal ecotones: a case study of the salt marsh/upland transition zone in California. J Ecol 93(2): 279–290.
Van Dyke, E., and K. Wasson. 2005. Historical ecology of a central California estuary: 150 years of habitat change. Estuaries 28(2): 173–189.
Wasson, K., and A. Woolfolk. 2011. Salt marsh–upland ecotones in central California: vulnerability to invasions and anthropogenic stressors. Wetlands 31(2): 1–14.
Wiens, J.A., C.S. Crawford, and J.R. Gosz. 1985. Boundary dynamics—a conceptual framework for studying landscape ecosystems. Oikos 45(3): 421–427.
Williams, P.B., and M.K. Orr. 2002. Physical evolution of restored breached levee salt marshes in the San Francisco Bay Estuary. Restor Ecol 10(3): 527–542.
Yarrow, M.M., and V.H. Marin. 2007. Toward conceptual cohesiveness: a historical analysis of the theory and utility of ecological boundaries and transition zones. Ecosystems 10: 462–476.
Zedler, J.B., J.C. Callaway, and G. Sullivan. 2001. Declining biodiversity: why species matter and how their functions might be restored in Californian tidal marshes. BioScience 51(12): 1005–1017.
Acknowledgments
We are grateful to the many people who helped with monitoring fieldwork over these 10 years: M. Bakker, C. Chabre, N. D'Amore, S. Fork, K. Madden, R. Martone, A. Morrow, J. Nicholson, and R. Preisler. J. Haskins and G. Lessa collected and helped to analyze the water level data that were instrumental to our inundation analyses. J. Haskins provided the precipitation data. K. Hayes provided access to a site managed by the Elkhorn Slough Foundation and R. de Bree and R. Stephens allowed us to approach two sites via Elkhorn Ranch. Two anonymous reviewers provided remarkably extensive and constructive feedback that improved the paper. This research was supported by a grant from NOAA's Estuarine Reserve Division to the California Coastal Conservancy and the Elkhorn Slough Foundation on behalf of the Elkhorn Slough National Estuarine Research Reserve, a partnership between NOAA and the California Department of Fish and Game.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wasson, K., Woolfolk, A. & Fresquez, C. Ecotones as Indicators of Changing Environmental Conditions: Rapid Migration of Salt Marsh–Upland Boundaries. Estuaries and Coasts 36, 654–664 (2013). https://doi.org/10.1007/s12237-013-9601-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-013-9601-8