
Abstract
We investigated compositions of plant communities and their abiotic determinants in the Yellow River estuary, China. Along a topographic gradient, we quantified plant compositions and abiotic factors in different vegetation zones, and examined the relationships between plant communities and abiotic factors using canonical correspondence analysis and the effects of vegetation shading using a removal experiment. The relationships between plant communities and abiotic factors differed between high elevations and low elevations. Salinity and flooding oppositely related with distributions of plant communities at low elevations, but they appeared to operate synergically at high elevations. The effects of vegetation shading were found to vary across the topographic gradient, indicating spatial variations in potential positive interactions among plants. These results suggest that spatial variations in determinants of community structure should be addressed in future studies in estuarine and coastal systems, as well as in other natural habitats.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Coastal habitats worldwide are distinguished for vegetation zones across steep environmental gradients (Chapman 1974). These conspicuous vegetation zones are generally considered to be products of mediation of both physical stress and plant competition (reviewed by Bertness and Pennings 2000). Trade-offs in plants between stress tolerance and competitive ability lead to competitively superior species monopolizing benign habitats and displacing competitively inferior species to stressful habitats. Despite of applicability in several coastal sites, this rule of salt marsh plant zonation has been criticized for generality across geographical regions (Bertness and Pennings 2000; Pennings et al. 2005; Fariña et al. 2009). Established rules of salt marsh community organization may not be directly applicable to unstudied systems (Kunza and Pennings 2008; Fariña et al. 2009).
An important hypothesis being tested currently is that rules of salt marsh community organizations vary among geographical sites on coasts (Bertness and Pennings 2000; Pennings et al. 2003; Pennings and Silliman 2005; Pennings et al. 2009). Geographic variations in climate may induce different salinity patterns across marsh elevations (Bertness and Pennings 2000; Wang et al. 2007). Taking latitude gradients in climate (which are currently repeatedly used to examine variations in patterns and processes of salt marshes) for example, in low-latitude salt marshes, the hot climate leads to elevated salinities at upper/middle elevations of the marshes compared with high-latitude salt marshes where the climate is much cooler and salinity is relatively low (Bertness and Pennings 2000). This difference in salinities between low- and high-latitude salt marshes was suggested to generate variations in mechanisms of salt marsh plant organizations (Pennings and Bertness 2000; Pennings et al. 2003). Saintilan (2009) also attributed latitudinal patterns of salt marsh plant diversity to climate-induced differences in salinity patterns between low and high latitudes in Australia. Besides differences in salinity patterns, Kunza and Pennings (2008) suggested differences in flooding regime to be the most plausible hypothesis accounting for differences in plant diversity between Georgia and Texas salt marshes.
At finer scales, mechanisms of salt marsh plant organization are suggested to vary with marsh elevation (Pennings and Callaway 1992; Lenssen et al. 1999; Fariña et al. 2009). In a Californian salt marsh, Pennings and Callaway (1992) indicated that although flooding, soil salinity, and competition all interacted to determine plant zonation patterns, the relative importance of these factors differed at different elevations. Moreover, the precise roles of the two main physical stressors, salinity and flooding, in mediating plant zonation were also suggested to vary with elevation in a Chinese salt marsh (He et al. 2009a).
Compared with salt marshes on the Atlantic coast of North America where most influential salt marsh plant zonation studies were conducted and in several other geographical regions, such as Australia (reviewed in Saintilan 2009), Western Europe (e.g., Bockelmann and Neuhaus 1999; van Wijnen and Bakker 1999; Huckle et al. 2002) and South America (e.g., Costa et al. 2003; Alberti et al. 2008), salt marshes on the western Pacific coast of Asia have been much less investigated. Chapman (1977) has classified salt marshes on the western Pacific coast of Asia as a Sino-Japanese group based on floristic and vegetation criteria. However, when Adam (1990) was doing a global review of salt marsh vegetation, he could found little information on the flora and vegetation of salt marshes in this geographic region (except for salt marshes in Japan). A few studies on Chinese salt marshes have risen in recent years (e.g., Wang et al. 2006; Wu et al. 2008; Ji et al. 2009). However, most of these studies focused on Spartina invasions rather than structures and organizations of native and characteristic species in Chinese salt marshes. Nevertheless, studies investigating structures and organizations of native and characteristic species in Chinese salt marshes are imperative facing the rapid invasion of Spartina from southern to northern salt marshes on the Chinese coast.
In this article, we investigate compositions and abiotic determinants of plant communities along a topographic gradient in the Yellow River estuary on the western Pacific coast of northern China, where most plant communities are still native but facing the invasion of Spartina alterniflora (hereafter, Spartina; He et al. 2009b). The marshes in the estuary are recently formed due to high sediment deposit and rapid land accretion (Li et al. 2009a). In such rapidly developing marshes, vegetation zonation has been suggested to be mainly controlled by abiotic factors either directly or indirectly via the alteration of competitive relationships (Wang et al. 2010). Here, we first describe the compositions of plant communities and abiotic conditions in different vegetation zones along the topographic gradient, and then test the following hypotheses specially: along the topographic gradient, (1) the relationships between plant communities and abiotic factors differ between low elevations and high elevations; (2) the effects of vegetation shading (and hence potential positive interactions) vary with elevation. Our study from a previously little-studied coastal geographic region provides new information on the hypothesis that mechanisms of coastal plant community organization vary geographically and within site.
Materials and Methods
Study Site
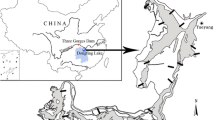
Field work was conducted at a site (Fig. 1; 37°46′ N, 119°09′ E) in the Yellow River estuary, northern China. Salt marshes and mudflats in the estuary are 1–6 km in width and are still accruing rapidly. Tides in the estuary are irregularly semidiurnal. The study site has a typical temperate monsoon climate, being dry and cool all the year round, except hot, rainy summers. Refer to Li et al. (2009a) for a detailed description of the climate condition.

Map of the study site and the topographic gradient. The arrow-headed line indicates the topographic gradient. Lines a–e, divisions between mudflat and Spartina low marsh, Spartina low marsh and Suaeda low marsh, Suaeda low marsh and high marsh, high marsh and terrestrial border, and terrestrial border and upland, respectively. Refer to He et al. (2009a) for the location of the study site on the western Pacific coast
The flora of the estuary is relatively simple. Dominant vascular plant species include Suaeda salsa (dominating low and high marshes; hereafter, Suaeda), Tamarix chinensis (dominating terrestrial borders of the salt marshes; hereafter, Tamarix), and Phragmites australis (dominating uplands near the river course, see Fig. 1; hereafter, Phragmites). In recent years, smooth cordgrass native to North America, Spartina, has invaded the mudflats in the estuary, becoming a dominant species in these habitats and is moving up rapidly. Other plant species including Salicornia europaea, Scripus spp., and Juncus spp. occur only sparsely or in small patches in the marsh habitats. Plant species in the uplands are much more diverse, including Calamagrostis pseudophragmites, Triarrhena sacchariflora, Imperata cylindrical, etc.
Parallel Transect Sampling
To quantify compositions of plant communities and their relationships with abiotic factors across topographic gradients from the lower limits of vascular plants to uplands, five zones (habitats) along the topographic gradient were delimitated based on elevation and vegetation: (1) the upland dominated by Phragmites; (2) the terrestrial border dominated by Suaeda and Tamarix; (3) the high marsh dominated by Suaeda; (4) the low marsh dominated by Suaeda (hereafter, Suaeda low marsh) with a few other species, such as Phragmites and S. europaea; and (5) the low marsh newly formed by Spartina invasions (hereafter, Spartina low marsh) (Fig. 1). In August 2009, we located two to five parallel transects of 80 m in length in each zone. The number of transects in each zone was approximately proportional to the width of that zone. We set a 2 × 2 m quadrat every 20 m along each transect (five quadrats in total for each transect) and recorded all herbaceous species and their cover values (visually estimated) in each quadrat. Cover values of shrubby and arboreal species were estimated in a 5 × 5 m quadrat concentrical with the corresponding 2 × 2 m quadrat. Most plant species including rare species were recorded. We used cover values (rather than other species variables) for analyses of vegetation compositions, since (1) species compositions in our study site were relatively simple and cover values could well describe structures of the plant communities; (2) cover values were one of the variables used frequently by previous studies in similar habitats (e.g. Vince and Snow 1984; Bertness and Ellison 1987; Traut 2005; Kunza and Pennings 2008). A soil core of topsoil (5.05 cm in diameter × 5 cm in depth) was collected at the center of each quadrat to determine soil pore water salinity, moisture content, bulk density and pH. Soil organic matter and total nitrogen were not determined, since our previous studies in the estuary suggested these factors had only minor relationships with variations in vegetation (He et al. 2009b).
Permanent Plot Sampling and Quantification of Abiotic Factors
To quantify among-zone and seasonal variations in abiotic conditions across the topographic gradient, we randomly established 8 1 × 1 m plots in each zone delimitated above and a soil core of topsoil (5.05 cm in diameter × 5 cm in depth) was collected in each plot monthly from June through August 2009 to determine soil pore water salinity, moisture content, bulk density and pH.
Soil cores were weighed, then dried at 60°C for 48 h in an oven and reweighed to determine moisture content and bulk density. Dried soil was mixed with a known volume of deionized water (5:1 aqueous suspension, 1-min vibration). The pH of the supernatant was measured after 30 min. The salinity of the supernatant was measured after 24 h using an electronic meter, and the original soil pore water salinity was calculated based on the initial moisture content of the core (Pennings et al. 2005).
Due to the harsh working conditions in the newly formed estuarine marsh (the marsh was very extensive and had many tidal creeks), daily observations of flooding at permanent locations were logistically difficult. Thus, we estimated the flooding frequency in each zone according to our field experience working in the estuary (>3 years). The estimated flooding frequencies were also verified by experienced local fishers.
Removal Experiment
As vegetation feedbacks may play an important role in mediating edaphic conditions, which meant both abiotic and biotic factors were important in determining patterns in these marshes, we further conducted a removal experiment to quantify the potential feedbacks of vegetation shading on edaphic conditions (other probable feedback effects, such as substrate stabilization and wave buffering as reviewed in Pennings and Bertness 2000, were not considered in the present paper). In early June 2009, we established 12 1 × 1 m plots (>20 m distances between plots) in each zone of the upland, the terrestrial border and the high marsh where vegetation shading was suggested to play a role in generating positive interactions in plants (Bertness and Pennings 2000). We clipped all the aboveground vegetation in half of these plots (randomly selected). The left plots (control) were maintained without treatments. Clipping treatments were repeated biweekly as needed. We did not establish shaded neighbor-removal treatments to mimic vegetation shading, since (1) such treatments were found to have similar effects on soil salinity as control treatments (although not for redox potential; see Bertness and Hacker 1994; Bertness and Yeh 1994) and (2) previous studies suggested that comparisons between control and neighbor-removal treatments should be an effective way to investigate the effects of vegetation shading on edaphic factors (Bertness and Hacker 1994; Bertness and Yeh 1994; Hacker and Bertness 1999; Pennings et al. 2003). In middle September 2009, soil cores were collected in all plots and six additional soil cores were collected in bare patches (typical in the terrestrial border but rare in other zones) which were naturally generated by disturbances and could often be permanent for years, and then edaphic factors were determined as described above.
Statistical Analysis
One-way analyses of variance (ANOVAs; followed by post hoc multiple comparisons, LSD test) were generally used to test differences in abiotic factors among zones along the topographic gradient, among months and among treatments in the removal experiment. Data were log10(x + 1) transformed if necessary to increase normality and homogeneity of variance. Those data that did not meet the assumptions of ANOVA after usual transformations were analyzed using nonparametric multiple comparisons (Steel test) or nonparametric Kruskal–Wallis tests. ANOVAs and nonparametric Kruskal–Wallis tests were performed using SAS Version 9.1 (SAS Institute, Cary, North Carolina, USA). Nonparametric multiple comparisons were performed using R Version 2.7 (R Development Core Team 2008).
The relationships between plant communities and abiotic factors were analyzed using canonical correspondence analysis (CCA). CCA is one of the most widely used gradient analyses by which species composition can be directly related to measured environmental variables (Palmer 1993). In order to investigate differences in the plant community-abiotic factor relationships between low-elevation habitats (Sparitna low marsh, Suaeda low marsh and high marsh) and high-elevation habitats (high marsh, terrestrial border and upland), two CCAs were performed for low-elevation habitats and high-elevation habitats, respectively. High marsh was considered as a transitional zone between the low-elevation and the high-elevation habitats (Traut 2005), and it was included in both low-elevation habitats and high-elevation habitats for the CCAs. An additional CCA was performed to detect the relationship between plant communities and abiotic factors across the entire topographic gradient. Species percentage cover values in all quadrats were log10(x + 1) transformed and used for CCA species matrices, and abiotic data including flooding frequency (medium values of the estimated ranges for each zone, see “Results”) were also log10(x + 1) transformed (Palmer 1993) and used for CCA environment matrices. CCA was performed using Canoco for Windows 4.5 (ter Braak and Šmilauer 2002).
Results
Plant Communities Across Topographic Gradients
Across the topographic gradient in the Yellow River estuary, compositions of plant communities differed among different zones (Table 1). In the low marsh, there were mainly two distinguishing vegetation zones: the Spartina zone seaward and the Suaeda zone landward. No other vascular plant species were found in the Spartina zone while several other species, such as Phragmites and S. europaea, occurred in relatively low coverage in the Suaeda zone (Table 1). On the lower border of the Suaeda zone, Spartina also formed some patches. In the high marsh, Suaeda was the only dominant plant species. Although there were some small patches of Phragmites, the mean coverage of Phragmites in the high marsh was much lower than in the Suaeda low marsh (Table 1). In the terrestrial border at higher elevations, a shrubby species, Tamarix, formed a conspicuous zone of shrubs, despite that Suaeda was still dense. In the upland, Phragmites replaced Suaeda and Tamarix and became the dominant species in this zone. Other main plant species that occurred in the upland included T. sacchariflora, C. pseudophragmites, and an arboreal species, Salix matsudana (Table 1).
Abiotic Factors Across Topographic Gradients
Among-zone variations
Flooding frequency generally decreased from the lowest Spartina zone to the upland. The Spartina and the Suaeda low marshes were flooded frequently while the terrestrial border and the upland were little flooded (Fig. 2a). The high marsh, as a transitional zone between low and high elevations, was flooded with a medium frequency of 5–20%. All the four edaphic factors determined differed significantly among different zones along the topographic gradient. Soil salinity reached a conspicuous peak in the high marsh, and it was much lower in the other zones both landward and seaward; in contrast, soil moisture content and pH were lowest in the high marsh but were highest in the upland and the Spartina low marsh (Fig. 2). Soil bulk density was relatively low in the upland and high in other zones (Fig. 2e).

Seasonal variations in abiotic factors from June through August 2009. a Flooding frequency; b soil salinity; c soil moisture content; d soil pH; and e soil bulk density. Data are shown as means ± SE except flooding frequency data which were shown as estimated maximal, minimum and medium values. For edaphic factors, the sample sizes are eight except those in the Spartina low marsh determined in June whose sample size are six, due to two samples’ lost during sample analysis. Significant differences among months are indicated by a star (P < 0.05) or two stars (P < 0.01) above each of the bar groups. For differences among zones, bars of each month in each subplot sharing a letter are not significantly different from one another. Statistical comparisons for the flooding frequency data which were estimated were infeasible
Seasonal variations
Soil salinity varied seasonally in each of the five zones while soil bulk density showed a strong seasonal variation only in the Sparitna low marsh (Fig. 2). Significant seasonal variations in soil moisture content were detected only in the terrestrial border, the high marsh and the Sparitna low marsh. Soil pH differed among months in each of the zones except the Suaeda low marsh. Despite of these seasonal variations, the patterns of among-zone variations along the topographic gradient for each factor were generally similar when determined in different months. For example, soil salinity was constantly highest in the high marsh zone and lowest in the upland and the Sparitna low marsh (Fig. 2b).
Relationships Between Plant Communities and Abiotic Factors
For the three zones at low elevations,
i.e., the Spartina low marsh, the Suaeda low marsh, and the high marsh: The first axis of the CCA ordination explained most of the total variance of the species-environment relation (Table 2). All the five abiotic factors significantly correlated with the first axis but showed no relationship with the second axis (Table 2). Flooding, moisture content, and pH positively correlated while salinity and bulk density negatively correlated with the first axis (Table 2). Plant communities in the Sparitna low marsh, the Suaeda low marsh and the high marsh were mainly distributed on the right, the middle and the left along the first axis, respectively (Fig. 3a).

CCA biplots showing relationships between plant communities and abiotic factors at low elevations (a), at high elevations (b), and across all elevations (c)
For the three zones at high elevations,
i.e., the high marsh, the terrestrial border, and the upland: the first axis of the CCA ordination also explained most of the total variance of the species-environment relation (Table 2). All the five abiotic factors significantly correlated with the first axis, and only flooding and moisture content showed significant relationships with the second axis (Table 2). Salinity, flooding and bulk density positively correlated while the others negatively correlated with the first axis. Plant communities in the upland were distributed on the left of the CCA biplot (Fig. 3b). In contrast, plant communities in the high marsh were mainly distributed at the lower right of the biplot. Plant communities in the terrestrial border were distributed on the upper right of the biplot (Fig. 3b).
For all the zones along the topographic gradient
The first axis and the second axis of the CCA ordination explained 58.2% and 29.3% of the total variance of the species-environment relation, respectively. All the abiotic factors significantly correlated with the first and the second axis, except bulk density which correlated only with the first axis (Table 2). Salinity, flooding and bulk density negatively correlated while pH and moisture content positively correlated with the first axis (Table 2). Flooding, moisture content, and pH positively correlated while salinity negatively correlated with the second axis (Table 2). Plant communities in the upland were distributed on the right of the CCA biplot (Fig. 3c). Plant communities in the terrestrial border and the high marsh were mainly distributed on the lower left of the biplot (Fig. 3c). The Suaeda and the Spartina low marshes were distributed on the upper left of the biplot (Fig. 3c).
Vegetation Feedbacks by Shading
Results from the removal experiment showed that vegetation removal significantly elevated soil salinities in the upland and in the terrestrial border rather than in the high marsh (Fig. 4a). Significant effects of shading on soil moisture content and bulk density were only detected in the upland (Fig. 4b, d). Significant effects of shading on soil pH were not detected in any zone (Fig. 4c).

Summary of the removal experiment investigating vegetation feedbacks by shading. Data are shown as means ± SE (n = 6). Differences between vegetation-present and vegetation-removal plots are indicated by the P values above each bar group. For bars of terrestrial border in each subplot, bars sharing a letter are not significantly different from one another
Compared with vegetation-removal plots in the terrestrial border, natural bare patches had a significantly higher level of soil salinity and bulk density (Fig. 4a, d). Differences in soil moisture content and pH between vegetation-removal plots and natural bare patches were not detected (Fig. 4b, c).
Discussion
Zonation Pattern in the Yellow River Estuary
In the Yellow River estuary, the most conspicuous vegetation zones are Suaeda-dominated low marshes and high marshes. Suaeda has wide tolerance to salt and flooding stress (Song et al. 2008; He et al. 2009a), being one of the main salt marsh plant species in salt marshes across the temperate coast of China (Yang and Chen 1995; Editorial Committee of Wetland Vegetation in China 1999). Suaeda dominates extensive marshes in the Yellow River estuary, extending from the low marsh frequently flooded to the high marsh and the inland terrestrial border where are little flooded and salinity are extremely high. In the terrestrial border at upper elevations, the shrubby species Tamarix grows relatively vigorous above Suaeda plants, forming a striking zone of shrubby plants along the estuary. Tamarix is also one of the main salt marsh plant species across the temperate coast of China (Editorial Committee of Wetland Vegetation in China 1999). Despite of the wide stress tolerance, Suaeda and Tamarix are replaced by dense Phragmites plants and patches of several other plant species in the upland.
Besides the native vegetation zones, we would like to address the emerging zone formed by Spartina invasions in the Yellow River estuary, due to the potential effects of Spartina invasions on patterns and processes of the marsh ecosystem in the estuary. The effects of Spartina invasions on the Yangtze estuarine marshes in southern China have already proved to be significant (Wang et al. 2006; Wu et al. 2008; Li et al. 2009b). In the Yellow River estuary in northern China, Spartina was only distributed in a few patches of commonly less than 10 m2 in size in the mudflat before 2007; Amazingly, in 2009, a Spartina marsh zone of several hundred meters in width was formed in the mudflat, and Spartina now is tending to spread rapidly seaward to newly formed mudflats in the estuary and landward to the native Suaeda low marshes (Qiang He, personal observation).
Determinants of Vegetation Zonation Across Elevations
In general, our study shows that abiotic factors correlate with the zonation of vegetation across the topographic gradient in the Yellow River estuary. Moreover, our results suggest that the relationships between plant communities and abiotic factors differ between low- and high-elevation habitats along the topographic gradient. Below, we would argue that flooding and salinity, the two most pronounced abiotic factors (Bertness and Ellison 1987; Bertness 1991a, b; Bertness et al. 1992; Pennings et al. 2005), play different roles in mediating vegetation zonation between low- and high-elevation habitats along topographic gradients in the estuary.
In the estuary, although flooding frequency generally decreased with increasing elevation along the topographic gradient, salinity reached a peak in the high marsh (Table 1 and Fig. 2b). At low elevations, flooding frequency decreased while salinity increased from the Sparitna low marsh through the high marsh. As revealed by the CCA ordination, flooding and salinity oppositely related with distributions of plant communities at the low elevations (Fig. 3a). The lower limits of these plant species, such as the low limit of Suaeda, at low elevations are most probably mediated by flooding rather than salinity (see experimental studies in He et al. 2009a), while their upper limits are most probably mediated by salinity. Besides, as the upper boundary of the current Spartina zone is still below the lower limit of Suaeda, the role of competition by Spartina in mediating the lower limit of Suaeda should be limited. However, with Spartina rapidly moving up towards the Suaeda low marsh, competition will be predicted to play a role in mediating the low limit of Suaeda a few years later.
At high elevations, however, both flooding frequency and salinity decreased with increasing elevation (Table 1 and Fig. 2b). Flooding and salinity appeared to synergically mediate distributions of plant communities at high elevations (Fig. 3b). Even though, the role of salinity may be more important in directly generating patterns of plant communities at high elevations. At least, salinity alone could well explain the segregation of plant communities between in the terrestrial border and in the upland (Fig. 3b). The extreme saline conditions of the terrestrial border (see Table 1 and Fig. 2b) are apparently beyond the salt tolerance of Phragmites who is only a pseudohalophyte (Nikolaevskii and Smirnova 1968) and who has limited salt tolerance (Adams and Bate 1999). However, combination of salinity and flooding rather than salinity alone would better explain the segregation of plant communities between in the high marsh and in the terrestrial border (Fig. 3b). Tamarix has a wider salt tolerance than Phragmites (He et al. 2008), yet we’d argue that the vigorous growth of Tamarix in the terrestrial border is probably mediated by positive interactions with Suaeda who often forms dense canopies and can significantly reduce soil salinities in the terrestrial border (see Fig. 4a). Thin dwarfish Suaeda plants in the high marsh had little effect on soil salinity (Fig. 4a), resulting in higher soil salinity levels in the high marsh than in the terrestrial border, despite of none statistical significance (see Table 1 and Fig. 3a). In contrast to the low limits of plants, the upper limit of Suaeda at high elevations was suggested to be controlled by competition (He et al. 2009a). Whether competition mediates the upper limits of Tamarix remains uninvestigated.
In addition to the pronounced roles of flooding and salinity in mediating the vegetation zonation, our study also indicates that soil moisture content, pH, and bulk density may play some role in mediating the vegetation zonation in the Yellow River estuary. It has been suggested that soil moisture content could affect seed germination of salt marsh plants (Noe and Zedler 2000) and that soil pH can affect performances of marsh plants by altering the form of inorganic ions available to the plants and the plant-mycorrhizal associations (Montemayor et al. 2008; Gormally and Donovan 2010). However, in our study, whether the statistically significant but limited variations in these abiotic factors do matter to the plants has to be further experimentally examined.
Taken together, our study supports the plant zonation paradigm which suggests that abiotic factors play a role in mediating coastal vegetation zonation (Bertness and Ellison 1987; Bertness 1991a, b; Bertness et al. 1992; Bertness and Pennings 2000; Pennings et al. 2005). Moreover, our study supports the current hypothesis that the precise controlling factors mediating vegetation zonation vary across elevations on coasts (Pennings and Callaway 1992; Lenssen et al. 1999; Fariña et al. 2009). As determinants of vegetation pattern can vary with elevation and differ due to the spatial scale used (see Matthews et al. 2009), spatial variations in the determinants of vegetation pattern should be more explicitly addressed in future studies.
The Role of Vegetation Feedbacks
Our study shows that the most significant effect of vegetation shading on edaphic conditions in the three zones at upper elevations is the effect of shading on soil salinity. Shading significantly reduced soil salinity in the upland and the terrestrial border but had little effect on soil salinity in the high marsh. The effect of shading was most significant in the terrestrial border where was flooded only during a very few exceptional high tides in late autumn and storm tides. In the terrestrial border, Suaeda often forms dense canopies whereas disturbance-generated bare patches where soil salinity is extremely high (see Fig. 4a) would be permanent without vegetation cover (Qiang He, personal observation). Although in the upland soils without vegetation shading still maintained a low level of salinities, vegetation shading also reduced soil salinity. The elevated soil salinity in the terrestrial border and the upland without vegetation shading may relate with changes in evaporative concentration of groundwater salts at the soil surface (Salama et al. 1999; Jobbágy and Jackson 2004; Werner and Lockington 2006). In contrast to the terrestrial border and the upland, vegetation removal had little effect on soil salinity in the high marsh where flooding frequency was about 5~20%. This is probably a result from relatively thin dwarfish Suaeda plants which have limited effects of shading in the high marsh.
These results confirm previous studies which suggested that the removal of vegetation would strongly elevate soil salinity (Bertness and Yeh 1994; Hacker and Bertness 1999; Brewer et al. 1997; Pennings et al. 2003). Moreover, our study indicates that vegetation shading can significantly influence edaphic conditions at further upper elevations, such as the upland and the terrestrial border besides the high marsh along coastal topographic gradients, and that vegetation shading may not play a role in high marshes where salt stress is extremely severe restricting plants from forming enough canopies to have a substantial feedback. Since vegetation feedbacks can meliorate edaphic conditions even in terrestrial borders and uplands where are little flooded but not in high marshes at middle tidal heights, positive interactions among plants are predictable to play a role in mediating organizations of plants at the landward end of and vary across coastal topographic gradients.
In conclusion, along the topographic gradient in the Yellow River estuary on the Pacific coast of northern China, different plant communities dominated different zones of varying abiotic conditions. The relationships between plant communities and abiotic factors differed between low- and high-elevation habitats along the topographic gradient. Salinity and flooding oppositely related with distributions of plant communities at the low elevations, but they appeared to operate synergically at the high elevations. The role of vegetation feedbacks can vary across the topographic gradients; at upper elevations beyond the high marsh, vegetation feedbacks (and hence potential positive interactions) can still play a role in mediating organization of plant communities. Spatial variations in determinants of community structure should be therefore addressed in future studies in estuarine and coastal systems, as well as in other natural habitats.
References
Adam, P. 1990. Saltmarsh ecology. Cambridge: Cambridge University Press.
Adams, J.B., and G.C. Bate. 1999. Growth and photosynthetic performance of Phragmites australis in estuarine waters: A field and experimental evaluation. Aquatic Botany 64: 359–367.
Alberti, J., M. Escapa, O. Iribarne, B. Silliman, and M. Bertness. 2008. Crab herbivory regulates plant facilitative and competitive processes in Argentinean marshes. Ecology 89: 155–164.
Bertness, M.D. 1991a. Interspecific interactions among high marsh perennials in a New England salt marsh. Ecology 72: 125–137.
Bertness, M.D. 1991b. Zonation of Spartina patens and Spartina alterniflora in a New England salt marsh. Ecology 72: 138–148.
Bertness, M.D., and A.M. Ellison. 1987. Determinants of pattern in a New England salt marsh plant community. Ecological Monographs 57: 129–147.
Bertness, M.D., and S.D. Hacker. 1994. Physical stress and positive associations among marsh plants. The American Naturalist 144: 363–372.
Bertness, M.D., and S.C. Pennings. 2000. Spatial variation in process and pattern in salt marsh plant communities in eastern North America. In Concepts and controversies in tidal marsh ecology, ed. M.P. Weinstein and D.A. Kreeger, 39–57. Dordrecht: Kluwer.
Bertness, M.D., and S.M. Yeh. 1994. Cooperative and competitive interactions in the recruitment of marsh elders. Ecology 75: 2416–2429.
Bertness, M.D., L. Gaugh, and S.W. Shumway. 1992. Salt tolerances and the distribution of plants across a New England salt marsh. Ecology 72: 1842–1851.
Bockelmann, A., and R. Neuhaus. 1999. Competitive exclusion of Elymus athericus from a high-stress habitat in a European salt marsh. Journal of Ecology 87: 503–513.
Brewer, J.S., J.M. Levine, and M.D. Bertness. 1997. Effects of biomass removal and elevation on species richness in a New England salt marsh. Oikos 80: 333–341.
Chapman, V.J. 1974. Salt marshes and salt deserts of the world. New York: Interscience.
Chapman, V.J. 1977. Introduction. In Ecosystems of the world 1: Wet coastal ecosystems, ed. V.J. Chapman. Amsterdam: Elsevier.
Costa, C.S.B., J.C. Marangoni, and A.G. Azevedo. 2003. Plant zonation in irregularly flooded salt marshes: Relative importance of stress tolerance and biological interactions. Journal of Ecology 91: 951–965.
Editorial Committee of Wetland Vegetation in China. 1999. Wetland vegetation in China. Beijing: Science.
Fariña, J.M., B.R. Silliman, and M.D. Bertness. 2009. Can conservation biologists rely on established community structure rules to manage novel systems? ... Not in salt marshes. Ecological Applications 19: 413–422.
Gormally, C.L., and L.A. Donovan. 2010. Responses of Uniola paniculata L. (Poaceae), an essential dune-building grass, to complex changing environmental gradients on the coastal dunes. Estuaries and Coasts, doi:10.1007/s12237-010-9269-2.
Hacker, S.D., and M.D. Bertness. 1999. Experimental evidence for factors maintaining plant species diversity in a New England salt marsh. Ecology 80: 2064–2073.
He, Q., B.S. Cui, X.S. Zhao, and H.L. Fu. 2008. Niches of plant species in wetlands of the Yellow River Delta under gradients of water table depth and soil salinity. Chinese Journal of Applied Ecology 19: 969–975.
He, Q., B. Cui, Y. Cai, J. Deng, T. Sun, and Z. Yang. 2009a. What confines an annual plant to two separate zones along coastal topographic gradients? Hydrobiologia 630: 327–340.
He, Q., B. Cui, X. Zhao, H. Fu, and X. Liao. 2009b. Relationships between salt marsh vegetation distribution/diversity and soil chemical factors in the Yellow River Estuary, China. Acta Ecologica Sinica 29: 676–687.
Huckle, J.M., R.H. Marrs, and J.A. Potter. 2002. Interspecific and intraspecific interactions between salt marsh plants: Integrating the effects of environmental factors and density on plant performance. Oikos 96: 307–319.
Ji, Y., G. Zhou, and T. New. 2009. Abiotic factors influencing the distribution of vegetation in coastal estuary of the Liaohe Delta, Northeast China. Estuaries and Coasts 32: 937–942.
Jobbágy, E.G., and R.B. Jackson. 2004. Groundwater use and salinization with grassland afforestation. Global Chang Biol 10: 1299–1312.
Kunza, A.E., and S.C. Pennings. 2008. Patterns of plant diversity in Georgia and Texas salt marshes. Estuaries and Coasts 31: 673–681.
Lenssen, J.P.M., F.B.J. Menting, W.H. van der Putten, and C.W.P.M. Blom. 1999. Control of plant species richness and zonation of functional groups along a freshwater flooding gradient. Oikos 86: 523–534.
Li, S., G. Wang, W. Deng, Y. Hu, and W.W. Hu. 2009a. Influence of hydrology process on wetland landscape pattern: A case study in the Yellow River Delta. Ecological Engineering 35: 1719–1726.
Li, B., C. Liao, X. Zhang, H. Chen, Q. Wang, Z. Chen, X. Gan, J. Wu, B. Zhao, Z. Ma, X. Cheng, L. Jiang, and J. Chen. 2009b. Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects. Ecological Engineering 35: 511–520.
Matthews, J.W., A.L. Peralta, D.N. Flanagan, P.M. Baldwin, A. Soni, A.D. Kent, and A.G. Endress. 2009. Relative influence of landscape vs. local factors on plant community assembly in restored wetlands. Ecological Applications 19: 2108–2123.
Montemayor, M.B., J.S. Price, L. Rochefort, and S. Boudreau. 2008. Temporal variations and spatial patterns in saline and waterlogged peat fields. 1. Survival and growth of salt marsh graminoids. Environmental and Experimental Botany 62: 333–342.
Nikolaevskii, V.G., and A.N. Smirnova. 1968. The effect of salinization of soil on the growth and yield of the common reed under conditions of a vegetation experiment. Rastit Resur 44: 552–554.
Noe, G.B., and J.B. Zedler. 2000. Differential effects of four abiotic factors on the germination of salt marsh annuals. American Journal of Botany 87: 1679–1692.
Palmer, M.W. 1993. Putting things in even better order: The advantages of canonical correspondence analysis. Ecology 74: 2215–2230.
Pennings, S.C., and M.D. Bertness. 2000. Salt marsh communities. In Marine community ecology, ed. M.D. Bertness, S.D. Gaines, and M.E. Hay, 289–316. Sunderland: Sinauer Associates.
Pennings, S.C., and R.M. Callaway. 1992. Salt marsh plant zonation: the relative importance of competition and physical factors. Ecology 73: 681–690.
Pennings, S.C., and B.R. Silliman. 2005. Linking biogeography and community ecology: Latitudinal variation in plant–herbivore interaction strength. Ecology 86: 2310–2319.
Pennings, S.C., E.R. Selig, L.T. Houser, and M.D. Bertness. 2003. Geographic variation in positive and negative interactions among salt marsh plants. Ecology 84: 1527–1538.
Pennings, S.C., M.B. Grant, and M.D. Bertness. 2005. Plant zonation in low-latitude salt marshes: disentangling the roles of flooding, salinity and competition. Journal of Ecology 93: 159–167.
Pennings, S.C., C.-K. Ho, C.S. Salgado, K. Więski, N. Davé, A.E. Kunza, and E.L. Wason. 2009. Latitudinal variation in herbivore pressure in Atlantic Coast salt marshes. Ecology 90: 183–195.
R Development Core Team. 2008. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing
Saintilan, N. 2009. Distribution of Australian saltmarsh plants. In Australian saltmarsh ecology, ed. N. Saintilan, 23–39. Collingwood: Csiro Publishing.
Salama, R.B., C.J. Otto, and R.W. Fitzpatrick. 1999. Contributions of groundwater conditions to soil and water salinization. Hydrogeology Journal 7: 46–64.
Song, J., H. Fan, Y. Zhao, Y. Jia, X. Du, and B. Wang. 2008. Effect of salinity on germination, seedling emergence, seedling growth and ion accumulation of a euhalophyte Suaeda salsa in an intertidal zone and on saline inland. Aquatic Botany 88: 331–337.
ter Braak, C.J.F., and P. Šmilauer. 2002. Canoco for Windows 4.5. Wageningen: Biometris-Plant Research International.
Traut, B.H. 2005. The role of coastal ecotones: a case study of the salt marsh/upland transition zone in California. Journal of Ecology 93: 279–290.
van Wijnen, H.J., and J.P. Bakker. 1999. Nitrogen and phosphorus limitation in a coastal barrier salt marsh: the implications for vegetation succession. Journal of Ecology 87: 265–272.
Vince, S.W., and A.A. Snow. 1984. Plant zonation in an Alaskan salt marsh: I. Distribution, abundance and environmental factors. Journal of Ecology 72: 651–667.
Wang, Q., C.H. Wang, B. Zhao, Z.J. Ma, Y.Q. Luo, J.K. Chen, and B. Li. 2006. Effects of growing conditions on the growth of and interactions between salt marsh plants: Implications for invasibility of habitats. Biological Invasions 8: 1547–1560.
Wang, H., Y.P. Hsieh, M.A. Harwell, and W. Huang. 2007. Modeling soil salinity distribution along topographic gradients in tidal salt marshes in Atlantic and Gulf coastal regions. Ecological Modelling 201: 429–439.
Wang, C.H., M. Lu, B. Yang, Q. Yang, X.D. Zhang, T. Hara, and B. Li. 2010. Effects of environmental gradients on the performances of four dominant plants in a Chinese saltmarsh: Implications for plant zonation. Ecological Research 25: 347–358.
Werner, A.D., and D.A. Lockington. 2006. Tidal impacts on riparian salinities near estuaries. Journal of Hydrology 328: 511–522.
Wu, Y.T., C.H. Wang, X.D. Zhang, B. Zhao, L.F. Jiang, J.K. Chen, and B. Li. 2008. Effects of saltmarsh invasion by Spartina alterniflora on arthropod community structure and diets. Biological Invasions 11: 635–649.
Yang, S., and J. Chen. 1995. Coastal salt marshes and mangrove swamps in China. Chinese Journal of Oceanology and Limnology 13: 318–324.
Acknowledgments
We thank Xianxian Liu, Yanyan Hua, Yanzi Cai, and Jiafeng Deng for invaluable field and/or laboratory assistance, Chenghuan Wang, Xiao Sun, two anonymous reviewers for critical comments, and Yueliang Liu and Juanzhang Lv for access to the study site. This study was funded by National Natural Science Foundation of China (U0833002), National Key Basic Research Program of China (2006CB403303), and the Fundamental Research Funds for the Central Universities (2009SD-24).
Conflicts of Interest Notification
We state that this manuscript have been read and approved by all authors listed on the title page, and that all authors have made substantial contributions to the design, execution, and reporting of this study. All financial interests held by the authors in any reported findings and all sources of financial support for the work have been described in the acknowledgments of the paper. We do not have a financial relationship with the organization that sponsored the research and no potential conflicts of interest exist to our knowledge and understanding. We maintain full control of all primary data, and we agree to allow the journal to review our data if requested.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
CUI, BS., HE, Q. & AN, Y. Community Structure and Abiotic Determinants of Salt Marsh Plant Zonation Vary Across Topographic Gradients. Estuaries and Coasts 34, 459–469 (2011). https://doi.org/10.1007/s12237-010-9364-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-010-9364-4