
Abstract
We investigated whether within wetland environmental conditions or surrounding land cover measured at multiple scales were more influential in structuring regional vegetation patterns in estuarine tidal wetlands in the Pacific Northwest, USA. Surrounding land cover was characterized at the 100, 250, and 1,000 m, and watershed buffer scales. Vegetation communities were characterized by high species richness, lack of monotypic zonation, and paucity of invasive species. The number of species per site ranged between 4 and 20 (mean ± standard deviation = 10.2 ± 3.1). Sites supported a high richness (mean richness of native species 8.7 ± 2.8) and abundance of native macrophytes (mean relative abundance 85 % ± 19 %). Vegetation assemblages were dominated by a mix of grasses, sedges, and herbs with Sarcocornia pacifica and Distichlis spicata being common at sites in the oceanic zone of the estuary and Carex lyngbyei and Agrostis stolonifera being common at the fresher sites throughout the study area. The vegetation community was most strongly correlated with salinity and land cover within close proximity to the study site and less so with land cover variables at the watershed scale. Total species richness and richness of native species were negatively correlated with the amount of wetland in the buffer at all scales, while abundance of invasive species was significantly correlated to within wetland factors, including salinity and dissolved phosphorus concentrations. Landscape factors related to anthropogenic disturbances were only important at the 100-m buffer scale, with anthropogenic disturbances further from the wetland not being influential in shaping the vegetation assemblage. Our research suggests that the traditional paradigms of tidal wetland vegetation structure and environmental determinants developed in east coast US tidal wetlands might not hold true for Pacific Northwest wetlands due to their unique chemical and physical factors, necessitating further detailed study of these systems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Ecologists have long been interested in the environmental controls of vegetation patterns in natural communities (Harper 1977; Tilman 1988). Historically, tidal wetlands have been ideal systems to study these questions due to their conspicuous zonation of both abiotic and biotic factors. Early studies led to the belief that species distributional patterns were governed by abiotic factors (e.g., Valiela and Teal 1974; Howes et al. 1981; Mendelssohn et al. 1981; Armstrong et al. 1985). However, continued study has resulted in the understanding that both abiotic and biotic factors play a role in structuring tidal marsh wetland communities. The current paradigm recognizes that harsh physical factors in areas of tidal marshes subject to frequent inundation result in occupation by stress-tolerant species while the more benign upper borders are occupied by competitively superior species. For example, the transplant studies of Bertness and Ellison (1987) in New England salt marshes demonstrated that the perennials Spartina patens and Juncus gerardii are confined to the high marsh due to low levels of oxygen and high salinities in the low marsh, while the low marsh dominant Spartina alterniflora is prevented from invading the high marsh due to interspecific competition. Through removal and transplant studies, Bertness (1991) determined that interspecific facilitation is an important mechanism structuring the high marsh community in disturbance driven bare patches. This pattern of physical factors and competition driving vegetation zonation patterns appears to hold true along the east coast of the USA (see also Pennings et al. 2005) and England (Castellanos et al. 1994; Castillo et al. 2000). However, the recent studies of Silliman and Zieman (2001) and Silliman and Bertness (2002) have pointed to the importance of grazers in structuring the vegetation community in these systems.
As useful as these paradigms are for understanding the processes governing the patterns of vegetation in tidal wetlands, it is unclear if they are generalizable across unstudied systems. In fact, Fariña et al. (2009) demonstrated that the mechanisms controlling plant zonation in Chilean salt marshes were different than those in New England and California salt marshes, with interspecific competition being responsible for vertical zonation of the plant community throughout the wetland. Bertness and Ewanchuk (2002) found that latitudinal variations in salinity and climate regimes will lead to different factors being important in structuring the vegetation community. In addition, Pennings et al. (2003) found that competitive interactions are more important than facilitation in southern marshes where floras are more salt tolerant.
The vegetation communities of Pacific Northwest tidal wetlands have been largely unstudied (but see Frenkel et al. 1981; Adamus et al. 2005). Pacific Northwest tidal wetlands exhibit several profound differences from the more studied Atlantic coast and California tidal wetlands. Pacific coast wetlands have mixed semidiurnal tides (Hickey and Banas 2003), resulting in diurnal fluctuations in the extent of vegetation exposed to tidal influence. They have strongly seasonal precipitation patterns, receiving most of their precipitation in winter months. This, in combination, with strong winter tidal forcing can lead to vast expanses of the marsh plain being inundated by lower saline water during the winter months. These wetlands tend to be small in size (Mitsch and Gosselink 2007) and often occur on accumulated sediment near river mouths (USGS 2000) or as small pockets fringing the bases of hillsides that reach the estuary edge. Often, the watersheds of these wetlands are largely undeveloped. However, these wetlands tend to be high in nutrients due to coastal upwelling off the Pacific Northwest coast (Hickey and Banas 2003). Finally, the vegetation community tends to be diverse, exhibiting less striking patterns of zonation with several species growing intermixed (Seliskar and Gallagher 1983; Adamus et al. 2005). Consequently, the likelihood that different mechanisms govern the vegetation community in these wetlands is high.
While examining within wetland factors is instrumental in understanding tidal wetland vegetation dynamics, examining larger-scale factors, e.g., surrounding land cover, becomes germane when one is interested in wetland conservation and management. Studies have demonstrated the influence of landscape factors on taxonomic groups other than vegetation (e.g., Lerberg et al. 2000; DeLuca et al. 2004; Bilkovic et al. 2006). Yet for the vegetation community, few studies have bridged the gap between the influence of localized, within wetland conditions and landscape level factors. Within Narragansett Bay, Wigand et al. (2003) demonstrated the importance of watershed nitrogen inputs. Bertness et al. (2002) related species composition to percent of the wetland border that was developed within this same estuary. On the west coast of the USA, Fetscher et al. (2010) examined vegetation communities in light of surrounding land cover and human population density, finding that tidal muting was more important in shaping vegetation than anthropogenic stress in the surrounding landscape. Thus, the question remains as to how environmental factors at multiple scales above the wetland level influence vegetation patterns.
While the importance of tidal wetlands has been well documented (e.g., Sather and Stuber 1984; MEA 2005), they occupy one of the most desirable locations for human development. Forty-four percent of the world’s population lives within 150 km of the coast (UN data), with the population in US coastal counties increasing by 28 % between 1980 and 2003 (Crossett et al. 2004). Subsequently, throughout history, tidal wetlands in the USA have been subjected to anthropogenic stresses of hydrologic modifications (e.g., filling, diking, ditching, drainage), fragmentation, and nutrient loading from surrounding urban areas. While many historic Pacific Northwest tidal wetlands have been lost due to conversion to pastureland and hydrologic modifications (Frenkel et al. 1981; Frenkel and Morlan 1991), present day anthropogenic stresses tend to be localized to a few estuaries (e.g., Columbia River, Puget Sound, WA, Coos Bay, OR, Tillamook Bay, OR). Many tidal wetlands in smaller estuaries are relatively unimpacted by human activities. However, development along the coast is projected to rise. This gradient of anthropogenic disturbance makes the Pacific Northwest an ideal location to study the scale at which these environmental factors regulate tidal marsh vegetation.
The purpose of this study was to examine vegetation patterns in Pacific Northwest tidal wetlands and the scale at which environmental factors influence vegetation. While environmental controls have been thoroughly examined in US Atlantic coast wetlands and wetlands along the southern US Pacific coast, wetlands of the Pacific Northwest remain largely unstudied. While previous work in other regions has focused on how within wetland conditions shape the vegetation community, this study examined the influence of environmental factors at multiple scales, ranging from within wetland to the upland contributing watershed, in shaping regional vegetation patterns. It is our aim to determine if Pacific Northwest tidal wetlands follow the paradigms of vegetation patterns and determinants developed for other geographic regions.
Methods
Study Area
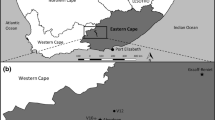
Fifty emergent tidal wetlands along the Oregon coast were sampled for this study (Fig. 1). Forty wetlands were sampled between July and September of 2007 and an additional ten wetlands were sampled during the same period of 2008. Sites were selected using a nonrandom design to cover geographic, salinity, and anthropogenic influence gradients. To ensure inclusion of sites along a gradient of condition, we selected sites that had previously been studied and rated for condition based on best professional judgment (Adamus et al. 2005). The sampling frame included all tidal wetlands along the Oregon Coast in two hydrogeomorphic (HGM) classes: marine-sourced low marsh and marine-sourced high marsh. Approximately equal numbers of sites in each HGM class were selected for study. Sampling locations within the wetland were chosen randomly. Data were collected within a 25-m radius circular area within each wetland at low tide.

Map showing the location of the study sites in coastal Oregon, USA estuaries
Vegetation
Emergent macrophytes were quantified along three transects within the wetland. Transects were oriented perpendicular to the water’s edge. One transect ran through the center of the wetland, and the other two transects were located 15 m on either side. Vegetation cover was quantified in 1 × 1 m quadrats at 10 m intervals along each transect for a total of 15 sampling points per site. The amount of bare ground, standing water, and litter material in each quadrat was also quantified. Percent cover of each species was estimated using a modified Daubenmire cover class system using a six-point scale. Cover of each species at the 15 sampling points per site was averaged to produce a single estimate of the cover of each species at each site.
Environmental Variables
Environmental variables were measured at fives scales: within wetland, in 100, 250, and 1,000 m buffers surrounding the wetland, and in the contributing watershed surrounding the wetland. Within wetland environmental variables included those characterizing channel water chemistry and soil chemistry. Variables within the buffers and the watershed characterized land cover, impervious surface area, road density, population density, and number of point source pollution discharges.
Water chemistry was characterized in the main tidal channel within the site or closest tidal channel to the study site. Measurements of dissolved oxygen, pH, temperature, and salinity were quantified at the surface and near the bottom within the channel using an YSI-6000 Multiparameter water quality meter. Since these measurements did not differ with depth, they were averaged. A 1-L water sample was collected at mid-depth within the tidal channel for the analysis of water column nutrients. The water sample was shaken thoroughly to ensure that no particulate matter had settled, and split into three subsamples for the analysis of total nitrogen and phosphorus, the sum of dissolved inorganic and organic nutrients, and dissolved inorganic nutrients.
Sediment samples were collected at five locations within the study site using modified 60 cm3 syringes: from the center and at each of the cardinal directions along the perimeter of the study site. Sediment samples were analyzed for total organic carbon, total nitrogen, and total phosphorus content. The sediment contents of the five samples from each study area were averaged for data analysis.
The landscape conditions (e.g., % forest cover, % impervious surface) surrounding the wetlands were evaluated at four scales using GIS: 100 m buffer, 250 m buffer, and 1,000 m buffer surrounding each study site, and the upland contributing watershed to the wetland (for those wetlands not located on islands). The direct upland watershed was calculated for the entire wetland within which the study site was located using ArcGIS spatial analyst and ArcHydro. Land cover was calculated using the following GIS data: Coastal Landscape Analysis and Modeling Study 1996 gradient nearest neighbor vegetation classes, Multi-Resolution Landscape Characteristics 2001 land cover data, Scranton (2004) tidal wetland classification for Oregon coastal wetlands, and USGS 10 m digital elevation models.
Data Analysis
Sites were classified as to whether they were on the mainland or on an isolated island. To examine if vegetation patterns differed among these groups, all analyses were run for all sites together, and for mainland (n = 37) and island sites (n = 13) separately, to account for the fact that watershed level data could not be generated for island sites. To account for the strong effect of salinity on vegetation composition, sites were classified into two groups of salinity influence, oceanic (n = 25) and mesohaline (n = 25). Classification was accomplished combining information from three sources: Lee and Brown (2009), NOAA (1998), and measured salinity. The Lee and Brown (2009) study divides estuaries into two zones based on detailed measurements of dry season dissolved inorganic nitrogen concentrations and salinity. The NOAA (1998) study divides Pacific Coast estuaries into three zones based on measured salinities (tidal fresh 0–0.5, mixing 0.5–25, and seawater >25).
Statistical analyses were performed using R (2.10.1/2009, R Foundation for Statistical Computing, Vienna, Austria). Student’s t tests were used to examine differences in environmental variables between mainland and island sites and oceanic and mixing sites. To examine vegetation patterns among sites, sites were ordered based on % cover of vegetation species composition using non-metric multidimensional scaling techniques (NMDS; metaMDS in the Vegan package of R). Only species present at more than one site were included in the NMDS analysis. The NMDS was performed using the Bray–Curtis distance measure. Environmental data were fit to the ordination using the ENVFIT function of the Vegan package. The ENVFIT function provides information on the strength of the correlation of the environmental vectors to the NMDS axes.
Regression tree change point analysis was used to explore the relationships between measures of vegetation community structure (NMDS axis 1, native species abundance), measures of species richness (total richness and native species richness), and environmental variables measured at five scales for all sites. Regression tree analysis can identify a set of important predictors, automatically handle interactions between predictor variables, and illustrate hierarchical relationships among predictor variables (Quinlan 1986; Venables and Ripley 2002). In this study, the various environmental parameters are the predictor variables. More details of regression tree analysis are presented in Weilhoefer and Pan (2008). Impurity within each node was measured as deviance by the Gini Index (Therneau and Atkinson 1997). A cross-validation procedure was used to determine when to stop partitioning the data. For this study, the 1-SE stopping rule was used (Therneau and Atkinson 1997).
Results
Vegetation
Species composition and relative abundance varied among sites. Sixty-five emergent plant species and three seagrass species were found throughout the study area (Table 1). The number of species at a site ranged between 4 and 20, with 10.2 species per site on average. Sampling plots within a site were often highly intermixed, with 4.5 ± 2.0 species on average (range 1–11) found within a 1 × 1 m area. Sites supported a high richness (mean richness of native species 8.7 ± 2.8) and abundance of native macrophytes (mean relative abundance 85 % ± 19 %). Common native species included grasses, sedges, and herbs, such as Carex lyngbyei (20 %), Sarcocornia pacifica (14 %), and Jaumea carnosa (11 %). Common invasive species included Agrostis stolonifera (13 %) and Phalaris arundinacea (1 %). Most sites had some amount of bare area (9 %) and water (7 %) present within the sampling area. While there were slight differences in % cover of individual species between mainland and island sites, none of these differences were significant.
Vegetation at sites classified as oceanic was dominated by succulent and salt tolerant species. The most abundant species at these sites included S. pacifica (23 %), J. carnosa (19 %), and Distichlis spicata (14 %). Grasses and sedges were dominant at sites in the mixing zone. Common species included C. lyngbyei (29 %), A. stolonifera (20 %), Juncus balticus (14 %), and Descampsia caespitosa (8 %). Species richness and diversity (Shannon’s Index) were not different between oceanic and mixing zone sites. However, invasive species were more abundant at sites in the mixing zone. Mean relative abundance of invasive species in the mixing zone was 22 % compared with 7 % in oceanic sites. A. stolonifera was an abundant invasive species at both oceanic and mixing sites, while the invasive species P. arundinacea was only present at sites in the mixing zone.
Physical and Chemical Data
Wetland sites displayed great variability in within wetland environmental conditions (Table 2). Sites ranged in salinity from nearly fresh conditions (1.4) to saline (32.8). On average, sites were mesohaline (mean = 24.5 ± 6.8). Oceanic zone sites had higher measured salinities, on average, than mixing zone sites. Tidal channel water was well-oxygenated (9.2 ± 2.4 mg/L) and of neutral pH (7.6 ± 0.3). These environmental parameters did not vary between mainland and island sites or between salinity classes. Concentrations of dissolved nutrients in tidal channels varied among wetlands, with some sites exhibiting high levels of both nitrogen and phosphorus. Tidal channel nutrient concentrations did not differ between mainland and island sites. However, oceanic zone sites had slightly elevated levels of total dissolved phosphorus, total phosphorus, and nitrate compared with mixing zone sites. Soil nitrogen, phosphorus, and carbon content were highly variable among sites. These environmental parameters did not vary between mainland and island sites or between oceanic and mixing zone sites.
Land Cover Data
Land cover surrounding the site varied depending on the scale at which it was quantified (Table 3). Land cover in the 100-m buffer surrounding the sites was dominated by wetlands (mean = 66 % ± 29 %) and secondarily by mudflats (14 % ± 20 %). This trend applied to all sites, regardless of whether they were islands or on the mainland and regardless of salinity regime. While some sites had high amounts of developed land and impervious surface area in their 100 m buffer, overall the percent of land occupied by these land covers was low (6 % ± 13 % and 2 % ± 4 %, respectively). Similar to the 100-m buffer, wetlands were the dominant land cover in the 250-m buffer surrounding the sites (52 % ± 28 %). However, cover by woodland became more prevalent at this scale. Developed land cover also was more abundant at this scale. Mudflat was more abundant and developed land was less abundant surrounding islands as compared to mainland sites. Mudflat was also more abundant surrounding oceanic zone sites.
At the 1,000-m scale, wetland was still the most abundant land cover (29 % ± 17 %), but it was less dominant than at smaller scales. Open water (14 % ± 13 %), mudflat (15 % ± 13 %), and developed (11 % ± 14 %) land covers were more abundant at this larger scale. Developed land is more common surrounding mainland sites than islands; however, its abundance did not differ between mixing zone and oceanic sites. Contributing watersheds to the sites were highly variable in size, ranging from 0.011 to 5.8 km2. At the watershed scale, forested land cover became the most abundant (45 % ± 33 %), followed by wetland (24 % ± 27 %) and woodland (22 % ± 19 %) land covers.
Vegetation Community Response to Environmental Conditions
To explore patterns in vegetation community structure, NMDS ordinations were developed for all sites, mainland sites, island sites, oceanic sites, and mixing sites. For all sites, NMDS analysis resulted in a two-dimensional solution (final stress = 0.18; Fig. 2). The first axis separated oceanic from mixing sites, with oceanic sites clustering on the negative end of axis 1 and mixing sites clustering on the positive end of axis 1. Island and mainland sites were not separated along the ordination axes. Axis 1 was highly negatively correlated with the salt tolerant succulents J. carnosa and S. pacifica and highly positively correlated with more freshwater species C. lyngbyei and Argentina egedii. In addition, the abundance of the majority of invasive species, including P. arundinacea and A. stolonifera, was positively correlated with axis 1. Axis 2 was positively correlated with Grindelia stricta and Atriplex patula and negatively correlated with the freshwater tolerant Schoenoplectus americanus and C. lyngbyei. NMDS ordinations for mainland, island, oceanic, and mixing stratifications showed similar relationships to species as the ordination for all sites.

Non-metric multidimensional scaling ordination plot of the 50 sites based on vegetation assemblages (sites in species space). Circles represent ocean sites and squares represent mixing sites. Closed circles and squares are mainland sites (n = 37) and open circles and squares are island sites (n = 13)
The vegetation community was highly correlated with wetland environmental conditions as well as with land cover variables at several scales (Table 4). When all sites were examined, axis 1 represented a salinity and phosphorus gradient, with both of these variables exhibiting significant negative correlations to this axis. Land cover variables at all scales that were most strongly correlated to vegetation community were those variables characterizing the amount of wetland, the amount of mudflat, and the amount of open water surrounding the site. In addition, developed land and impervious surface area with the buffer immediately surrounding the wetland (100 m) were strongly correlated to vegetation community, while no significant correlations were found for land cover variables related to human development in the 250- and 1,000-m buffers. When only mainland sites were examined, similar environmental variables were strongly related with vegetation structure. In contrast to the vegetation community for all sites, when only mainland sites were examined, no land cover variables related to human development were significantly correlated to the vegetation community. Land cover variables at the watershed scale were less strongly correlated to the vegetation community than those at smaller scales. Vegetation community was not related to site latitude.
When sites were classified by salinity, vegetation communities were correlated with different environmental variables. At the within wetland scale, the vegetation community structure in oceanic sites was significantly correlated with nitrate and phosphate in the tidal channels. In contrast, vegetation community structure at sites located within the mixing zone of the estuaries was most significantly correlated with channel water salinity. The vegetation community in oceanic sites was significantly correlated with land cover variables related to the amount of wetland surrounding the wetlands at the 100-, 250-, and 1,000-m scales, as well as the amount of woodland in the 250- and 1,000-m buffers. In contrast, for mixing sites, only the amount of wetland immediately surrounding the site (100 m buffer) was significantly correlated to the vegetation assemblages.
Species richness parameters displayed similar relationships with environmental variables as did the vegetation community. For all sites, total species richness and richness of native species were most significantly correlated with variables characterizing the amount of wetland and mudflat surrounding the site, with high total richness and high richness of native species being correlated with lower amounts of wetlands surrounding the site (total richness: r = −0.34 for % wetlands in 100 m buffer, −0.40 for 250 m buffer, and −0.32 for 1,000 m buffer; native richness: r = −0.37 for % wetlands in 100 m buffer, −0.49 for 250 m buffer, and −0.49 for 1,000 m buffer). Native species abundance was negatively correlated with the amount of broadleaf vegetation in all buffers surrounding the wetland. In contrast, abundance of invasive species was significantly correlated with water quality variables, including salinity (r = −0.56) and dissolved phosphorus concentrations (r = −0.32), indicating higher amounts of invasive species at sites with lower salinities and nutrients. Patterns in richness and invasive species metrics were similar when sites were stratified into island and mainland sites and ocean and mixing sites (data not presented).
Regression trees were developed for overall vegetation community composition (NMDS1), native species richness, and native species abundance using all sites and the subset of sites on the mainland, with predictive powers (r 2) ranging between 0.25 and 0.68 (Fig. 3). Based on cross validation, no regression tree could be developed for total species richness. Vegetation community structure (NMDS1, native species abundance) was determined primarily by salinity and secondarily by amount of wetland surrounding the study site. Salinity less than 22 predicted high positive axis scores, corresponding to freshwater taxa and invasive taxa, and abundance of native species greater than 90 %. In contrast, native species richness for all sites was determined by amount of wetland surrounding the site or the amount of broadleaf vegetation in the 1,000-m buffer for mainland sites. Models developed for native species richness had only one split and had lower predictive power. For all sites, amount of wetland in the 250-m buffer less than 60 % predicted greater richness. For mainland sites, broadleaf vegetation in the 1,000-m buffer less than 15 % predicted higher native richness.

Regression tree structure for NMDS1 axis scores, native species abundance, and native species richness for all sites (n = 50) and mainland sites (n = 37). Terminal nodes give the predicted NMDS score (a, b), relative abundance of native species (c, d), and native richness (e, f) and the number of sites in that group (in parenthesis). The values beside each split represent the critical threshold of given variables which provide the basis for that split (1,000.wet = % wetland in 1,000 m buffer, 1,000.MSH = % marine-sourced high marsh wetland in 1,000 m buffer, 250.wet = % wetland in 250 m buffer, 1,000.BL = % broadleaf vegetation in 1,000 m buffer)
Discussion
This study represents the first large-scale study of vegetation patterns and relationships with environmental variables for Pacific Northwest tidal wetlands. Our results point to four key differences between vegetation in these wetlands and those in California and the north east coast of the USA, where much of the knowledge of tidal wetland functioning has been developed. The first striking difference is the lack of strong vegetation zonation, with on average 10.2 species within a 25-m radius area. In addition, when the entire marsh plain is considered, the species richness is comparable with or slightly higher than marshes along the US southeastern and Gulf coasts, which average 10–12 species (Richards et al. 2005; Kunza and Pennings 2008). Secondly, plots were highly intermixed, with 4.5 species on average observed within the sampling quadrat. This contrasts with US Atlantic and Gulf coast tidal wetlands where monotypic zones are observed (e.g., Miller and Egler 1950; Chapman 1974; Niering and Warren 1980). In addition, in contrast with the dense vegetation cover (>95 %) observed in New England salt marshes (Bertness and Ellison 1987) most plots had some amount of bare area. In these regards, Pacific Northwest tidal wetlands appear to be more similar to wetlands of California and the southern USA. Zedler et al. (1999) characterized southern California salt marshes as species-rich, with an average of 4.38 species found within 0.25-m2 quadrats. While southern US marshes often contain large unvegetated salt pans (e.g., Pennings and Richards 1998). A third difference is that the dominant species in Pacific Northwest wetlands differed from east coast tidal wetlands, with the sedge C. lyngbyei, and two succulent forbs, S. pacifica and J. carnosa, being the most abundant species found throughout the study area. While S. pacifica is a common species found in tidal wetlands in southern and central California (Zedler 1982; Zedler et al. 1999; Fetscher et al. 2010), other dominant species in the Pacific Northwest were not as abundant in California tidal wetlands. Earlier work on vegetation communities in Oregon tidal wetlands noted that the disjunction between Oregon and California flora occurs at the Rogue and Chetco Rivers, south of our current study area (Jefferson 1975). We found no latitudinal patterns in vegetation composition or richness metrics, indicating that the tidal vegetation, while diverse, is relatively uniform in this area.
A final difference between vegetation patterns in Pacific Northwest tidal wetlands was the relative lack of invasive species. In this regard, Pacific Northwest tidal wetlands are similar to those in the US southeast where no invasive species are found. While several invasive species, including A. stolonifera and P. arundinacea, were encountered in this study, overall invasive species richness and abundances were low at most sites. Invasive species were more common at mixing sites, with most invasive species being salt water intolerant. Along the northeast coast, Phragmites australis is a common invasive species in tidal wetlands with altered hydrology, often forming vast monotypic expanses (King et al. 2007) and Fetscher et al. (2010) found 8.5 times more invasive species cover in tidally muted than fully tidal systems. The majority of wetlands in our study had intact hydrologic regimes. We hypothesize that this high diversity, low invasive species abundance, different vegetation composition, and lack of zonation in Pacific Northwest tidal wetlands results from natural differences in their physical (e.g., size, tidal regime) and chemical (e.g., coastal upwelling) nature. In addition, many tidal wetlands in Oregon are located in pockets fringing hillsides and are relatively unimpacted by human development, also contributing to the lack of invasive species in this area.
Vegetation composition in tidal wetlands is controlled by the interaction of environmental factors acting at multiple scales. In this study, patterns of vegetation in these wetlands were correlated with conditions within the wetland or immediately surrounding the wetland. Vegetation community composition was most strongly correlated to site salinity and metrics of the amount of wetland in close proximity to the site, and not anthropogenic land covers. Experimental and survey work have demonstrated that plants in tidal wetlands have varying tolerances to salinity (e.g., Barbour and Davis 1970; Crain et al. 2004). At the within wetland scale, several studies have demonstrated that the physical stresses of salinity and water-logging structure the vegetation community within the physically stressful, low marsh area of tidal wetlands and competition structures the community in the more hospitable upper marsh area (Bertness and Ellison 1987; Bertness 1991). In contrast, in hypersaline southern California salt marshes, interspecific competition becomes less important in the high marsh (Pennings and Callaway 1992). How salinity works to structure vegetation patterns on a landscape scale has been less well studied. In a transplant study of ten common species, Crain et al. (2004) demonstrated that the distribution of vegetation along estuarine gradients is driven by differential tolerance to salt stress and plant competition, with fresh-water wetland species being restricted from salt marshes by physical factors and salt tolerant species being restricted from freshwater wetlands by interspecific competition. Our study covered a wide salinity gradient, with site salinities ranging from oligohaline to euhaline, and it appears that distinct vegetation assemblages occupy different salinity zones within the estuary.
While salinity was instrumental in shaping the overall vegetation community, species richness was only weakly, negatively correlated to site salinity. Two of the three highest species richness were found at two sites with oligohaline (<4.6) salinities, while the highest species richness was found at a site with elevated salinity (30.9). This finding contrasts with several studies that have demonstrated vegetation richness increasing along decreasing salinity gradients (Anderson et al. 1988; Odum 1988; Greenberg et al. 2006). While a negative relationship between salinity and richness has been demonstrated for many estuaries, Sharpe and Baldwin (2009) did observe an interaction between salinity and urbanization for tidal wetlands covering a 0.5–18 salinity gradient. They found that in undeveloped estuaries, richness was higher at transitional sites than freshwater and brackish sites, while in urbanized estuaries, richness decreased linearly with salinity. The vegetation at sites in this study tends to be much more diverse than the aforementioned studies, possibly leading to a different relationship between richness and salinity. In addition, this study covers a large salinity gradient and the impacts of urbanization are restricted to a few sites.
Of secondary importance in structuring tidal wetland vegetation communities in the Pacific Northwest is the amount of wetland and mudflat in close proximity to the study site. Richness of native species was negatively correlated with amount of wetland in the buffer at all scales and positively correlated with amount of developed land in the 100-m buffer. Multiple land covers surrounding the wetland may serve as a source of terrestrial and upland colonizers, contributing to a more diverse wetland community. For saline sites (>22), the amount of wetland in the 1,000-m buffer was positively correlated with the amount of freshwater and upland species at the site. Freshwater wetlands surrounding the saline sites may lead to higher rates of colonization by freshwater wetland species due to the ecological connectivity of these systems (MacArthur and Wilson 1967). In addition, invasive species abundance was negatively correlated with the amount of mudflat surrounding the sites. Mudflat bordering the wetland can be considered a wetland edge in a natural state and would not result in the negative consequences produced by a human impacted edge.
Possibly of even greater interest than the importance of buffer wetlands in shaping vegetation communities in Pacific Northwest tidal wetlands is the fact that anthropogenic activities surrounding the wetlands seem to be of much less importance in determining vegetation patterns. The amount of anthropogenic influence (developed and impervious land covers) surrounding the sites was highly variable, with some sites exhibiting high levels of influence. Other authors have demonstrated that anthropogenic land covers may alter wetland vegetation composition and richness by serving as a source of exotic species and physical disruption of soil and hydrology (Callaway and Zedler 2004; Sharpe and Baldwin 2009). In this study, the amount of developed and impervious land cover in the 100-m buffer was the only anthropogenic land cover variables associated with vegetation assemblages (NMDS axis scores). Upon closer inspection, these land covers are highly correlated to sites with high abundances of invasive species, particularly, Elymus repens, Holcus lanatus, and Lotus corniculatus, indicating that anthropogenic land covers might provide a corridor enabling exotic species to invade wetlands. Bertness et al. (2002) and King et al. (2007) found a positive relationship between the amount of the invasive species P. australis in the wetland border and the amount of development in the wetland border. In contrast, in California tidal wetlands, Fetscher et al. (2010) observed no significant relationships between watershed level population densities and developed land cover in the surrounding habitat patch and invasive species cover. The differences in relationships between vegetation and developed land cover among studies may be due to the different scales at which land cover was characterized in the studies. For freshwater wetlands, Houlahan et al. (2006) observed that land cover in the 250–300-m buffer was most strongly correlated with vegetation richness, with higher overall richness and lower richness of exotic species in sites with more forested land cover. Brown and Vivas (2004) recommend 100 m from wetland edge as the ideal scale at which to characterize land cover impacts to wetlands. In our study, anthropogenic impacts appear to only be important at the scale immediately surrounding the wetland.
Urban land cover may also influence tidal wetland vegetation by serving as a source of nutrients. Most of the work on landscape level patterns of tidal wetland vegetation has focused on the importance of nitrogen availability from urban land covers, with several authors demonstrating positive relationships between urban land cover and nitrogen loading to surrounding wetlands and subsequent decreases in species richness and shifts in species distributions with increased nitrogen loading (e.g., Bertness et al. 2002; Wigand et al. 2003). Other studies have noted an increase in the cover of the invasive species P. australis with increasing nitrogen inputs from urbanization (Silliman and Bertness 2002; King et al. 2007). In this study, water column nitrate levels showed a weak positive correlation with developed land in the 100- and 250-m buffers surrounding the wetland, indicating that anthropogenic land covers may serve as a source of nitrogen to these wetlands.
While anthropogenic land covers may serve as a source of nitrogen to wetlands, it is important to consider that Pacific Northwest estuaries may be high in nitrogen due to several natural sources. A coastal upwelling zone occurs off the coast of Oregon, serving as a source of nutrients to coastal waters. In addition, many coastal watersheds that underwent logging historically are dominated by red alder, Alnus rubra, a species with a symbiotic nitrogen fixing association. Naymik et al. (2005) noted elevated levels of nitrogen in Oregon streams with watersheds dominated by red alder. In addition, nitrate levels in excess of 100 μM have been recorded in Yaquina Bay, OR, tidal wetlands with watersheds dominated by red alder (EPA unpublished data). In this study, the highest sediment total nitrogen levels were found at sites with a high abundance of J. balticus, another species with a nitrogen fixing symbiosis. The findings of this study raise questions as to whether the traditional paradigm of nitrogen poorness of salt marshes (e.g., Valiela and Teal 1974) and nitrogen limitation of primary production holds true for Pacific Northwest tidal wetlands.
Finally, the results of this study point to several implications for tidal wetland management and assessment. First, the vegetation community varies widely across salinity gradients. Therefore, it will be necessary to conduct protection and restoration efforts across this salinity gradient in order to conserve all vegetation and habitat types. Secondly, salinity is such a driving factor behind the vegetation community that it might be necessary to stratify assessments by site salinity to accurately evaluate the vegetation community. Thirdly, both within wetland and near-wetland land cover conditions are important in structuring the vegetation community and therefore stressors at both of these scales should be measured to accurately assess impacts to these systems. Finally, developed land cover in close proximity to the wetland seems to be a stressor to vegetation and therefore should be included in assessments of these systems. Future studies on these systems should focus on the role that increasing urbanization may play in shaping the vegetation communities.
References
Adamus, P.R., J. Larsen, and R. Scranton. 2005. Wetland profiles of Oregon’s coastal watersheds and estuaries. Part 3 of a Hydrogeomorphic Guidebook, Report to Coos Watershed Association, US EPA, and Oregon Department of State Lands, Salem, OR.
Anderson, R.R., R.G. Brown, and R.D. Rappleye. 1988. Water quality and plant distribution along the upper Patuxent River, MD. Chesapeake Science 9: 145–156.
Armstrong, W., E.J. Wright, S. Lythe, and T.J. Gaynard. 1985. Plant zonation and the effects of the spring-neap tidal cycle on soil aeration in a Lumber salt marsh. Journal of Ecology 73: 323–339.
Barbour, M.G., and C.B. Davis. 1970. Salt tolerance of five California salt marsh plants. American Midland Naturalist 84: 262–265.
Bertness, M.D. 1991. Interspecific interactions among high marsh perennials in a New England salt marsh. Ecology 72: 125–137.
Bertness, M.D., and A.M. Ellison. 1987. Determinants of pattern in a New England salt marsh plant community. Ecological Monographs 57: 129–147.
Bertness, M.D., and P.J. Ewanchuk. 2002. Latitudinal and climate-drive variation in the strength and nature of biological interactions in New England salt marshes. Oecologia 132: 392–401.
Bertness, M.D., P.J. Ewanchuk, and B.R. Silliman. 2002. Anthropogenic modification of New England salt marsh landscapes. Proceedings of the National Academy of Sciences 99: 1395–1398.
Bilkovic, D.M., M. Roggero, C.H. Hershner, and K.H. Haves. 2006. Influence of land use on macrobenthic communities in nearshore estuarine habitats. Estuaries and Coasts 29: 1185–1195.
Brown, M.T., and M.B. Vivas. 2004. Landscape development intensity index. Environmental Monitoring and Assessment 101: 1–21.
Callaway, J.C., and J.B. Zedler. 2004. Restoration of urban salt marshes: lessons from southern California. Urban Ecosystems 7: 107–124.
Castellanos, E.M., M.E. Figueroa, and A.J. Davy. 1994. Nucleation and facilitation in salt marsh succession: interactions between Spartina maritima and Arthrocnemum perenne. Journal of Ecology 82: 239–248.
Castillo, J.M., L. Fernández-Baco, E.M. Castellanos, C.J. Luque, M.E. Figueroa, and A.J. Davy. 2000. Lower limits of Spartina densiflora and S. maritima in a Mediterranean salt marsh determined by different ecophysiological tolerances. Journal of Ecology 88: 801–812.
Chapman, V.J. 1974. Salt marshes and salt deserts of the world, 392. Germany: J. Cramer.
Crain, C.M., B.R. Silliman, S.L. Bertness, and M.D. Bertness. 2004. Physical and biotic drivers of plant distribution across estuarine salinity gradients. Ecology 85: 2539–2549.
Crossett, K.M., T.J. Culliton, P.C. Wiley, and T.R. Goodspeed. 2004. Population trends along the Coastal US: 1980–2008. NOAA Report, Washington D.C.
DeLuca, W.V., C.E. Studds, L.L. Rockwood, and P.P. Marra. 2004. Influence of land use on the integrity of marsh bird communities of the Chesapeake Bay, USA. Wetlands 24: 837–847.
Fariña, J.M., B.R. Silliman, and M.D. Bertness. 2009. Can conservation biologist rely on established community structure rules to manage novel systems?… Not in salt marshes. Ecological Applications 19: 413–422.
Fetscher, A.E., M.A. Sutula, J.C. Callaway, V.T. Parker, M. Vasey, J.N. Collins, and W.G. Nelson. 2010. Patterns of vegetation communities of estuarine wetlands in two California regions: insights from a probabilistic survey. Wetlands 30: 833–846.
Frenkel, R.E., and J.C. Morlan. 1991. Can we restore our salt marshes? Lessons from the Salmon River, Oregon. Northwest Environmental Journal 7: 119–135.
Frenkel, R.E., H.P. Eilers, and C.A. Jefferson. 1981. Oregon coastal salt marsh upper limits and tidal datums. Estuaries 4: 195–205.
Greenberg, R., J. Maldonado, and M.V. McDonald. 2006. Tidal marshes: a global perspective on the evolution and conservation of their terrestrial vertebrates. BioScience 56: 675–685.
Harper, J.L. 1977. Population biology of plants, 992. London: Academic.
Hickey, B.M., and N.S. Banas. 2003. Oceanography of the U.S. Pacific Northwest coastal ocean and estuaries with application to coastal ecology. Estuaries 26: 1010–1031.
Houlahan, J.E., P.A. Keddy, K. Makkay, and C.S. Findlay. 2006. The effects of adjacent land use on wetland species richness and community composition. Wetlands 26: 79–96.
Howes, B.L., R.W. Howard, J.M. Teal, and I. Valiela. 1981. Oxidation-reduction potentials in salt marshes: spatial patterns and interactions with primary production. Limnology and Oceanography 26: 350–360.
Jefferson, C.A. 1975. Plant communities and succession in Oregon coastal salt marshes. Ph.D. Dissertation, Oregon State Univ. Corvallis, OR, unpublished.
King, R.S., W.V. DeLuca, D.F. Whigham, and P.P. Marra. 2007. Threshold effects of coastal urbanization on Phragmites australis (common reed) abundance and foliar nitrogen in Chesapeake Bay. Estuaries and Coasts 30: 469–481.
Kunza, A.E., and S.C. Pennings. 2008. Patterns of plant diversity in Georgia and Texas salt marshes. Estuaries and Coasts 31: 673–681.
Lee II, H. and Brown, C.A. (2009) Classification of regional patterns of environmental drivers and benthic habitats in Pacific Northwest Estuaries. US EPA, Office of Research and Development, National Health and Environmental Effects Research Laboratory, Western Ecology Division, Newport, OR.
Lerberg, S.B., A.F. Holland, and D.M. Sanger. 2000. Responses of tidal creek macrobenthic communities to the effects of watershed development. Estuaries 23: 838–853.
MacArthur, R.H., and E.O. Wilson. 1967. The theory of island biogeography, 224. Princeton: Princeton University Press.
MEA. 2005. Ecosystems and human well-being: general synthesis. Millennium ecosystems assessment report, 160. Washington: Island Press.
Mendelssohn, I.A., K.L. McKee, and W.H. Patrick. 1981. Oxygen deficiency in Spartina alterniflora roots: metabolic adaptation to anoxia. Science 214: 439–441.
Miller, W.B., and F.E. Egler. 1950. Vegetation of the Wequetequock-Pawcatuck tidal marshes, Connecticut. Ecological Monographs 20: 143–172.
Mitsch, W.J., and J.G. Gosselink. 2007. Wetlands, 600. Hoboken: Wiley.
Naymik, J., Y. Pan, and J. Ford. 2005. Diatom assemblages as indicators of timber harvest effects in coastal Oregon streams. Journal of the North American Benthological Society 24: 569–584.
Niering, W.A., and R.S. Warren. 1980. Vegetation patterns and processes in New England salt marshes. BioScience 30: 301–307.
NOAA. 1998. NOAA’s Estuarine Eutrophication Survey, Volume 5: Pacific Coast Region. Silver Spring: Office of Ocean Resources Conservation and Assessment.
Odum, W.E. 1988. Comparative ecology of tidal freshwater and salt marshes. Annual Review of Ecology and Systematics 19: 147–176.
Pennings, S.C., and R.M. Callaway. 1992. Salt marsh plant zonation: the relative importance of competition and physical factors. Ecology 73: 681–690.
Pennings, S.C., and C.L. Richards. 1998. Effects of wrack burial in salt-stressed habitats: Batis maritima in a southwest Atlantic salt marsh. Ecography 21: 630–638.
Pennings, S.C., E.R. Selig, L.T. Houser, and M.D. Bertness. 2003. Geographic variation in positive and negative interactions among salt marsh plants. Ecology 84: 1527–1538.
Pennings, S.C., M. Bestor-Grant, and M.D. Bertness. 2005. Plant zonation in low-latitude salt marshes: disentangling the roles of flooding, salinity and competition. Journal of Ecology 93: 159–167.
Quinlan, J.R. 1986. Induction of decision trees. Machine Learning 1: 81–106.
Richards, C.L., S.C. Pennings, and L.A. Donovan. 2005. Habitat range and phenotypic variation in salt marsh plants. Plant Ecology 176: 263–273.
Sather, J.H., and P.J.R. Stuber. 1984. Proceedings of the national wetlands values assessment workshop. US Fish and Wildlife Service, Western Energy and Land Use Team, Washington, D.C.
Scranton, R. 2004. The application of Geographic Information Systems for delineation and classification of tidal wetlands for resources management of Oregon’s coastal watersheds. Master’s Thesis, Oregon State Univ, Corvallis, OR.
Seliskar, D.M., and J.L. Gallagher. 1983. The ecology of tidal marshes of the Pacific Northwest coast: a community profile. Washington: U.S. Fish and Wildlife Service, Division of Biological Services.
Sharpe, P.J., and A.H. Baldwin. 2009. Patterns of wetland plant species richness across estuarine gradients of the Chesapeake Bay. Wetlands 29: 225–235.
Silliman, B.R., and M.D. Bertness. 2002. A trophic cascade regulates salt marsh primary production. Proceedings of the National Academy of Sciences of the United States ofAmerica 99: 10500–10505.
Silliman, B.R., and J.C. Zieman. 2001. Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh. Ecology 82: 2830–2845.
Therneau, T.M., and E.J. Atkinson. 1997. An introduction to recursive partitioning using the RPart routines. Rochester: Mayo Foundation.
Tilman, D. 1988. Plant strategies and the dynamics and structure of plant communities, 360. Princeton: Princeton University Press.
USGS. 2000. Oregon Wetland Resources. http://or.water.usgs.gov/pubs/Online/Html/WSP2425/. Accessed 2/17/2011.
Valiela, I., and J.M. Teal. 1974. Nutrient limitation of salt marsh vegetation. In Ecology of Halophytes, ed. R.J. Reimold and W.H. Queen, 547–563. New York: Academic.
Venables, W.N., and B.D. Ripley. 2002. Modern and applied statistics with S-Plus, 520. New York: Springer.
Weilhoefer, C.L., and Y. Pan. 2008. Using change-point analysis and weighted averaging approaches to explore the relationships between common benthic diatoms and in-stream environmental variables in Mid-Atlantic Highlands streams, USA. Hydrobiologia 614: 259–274.
Wigand, C., R.A. McKinney, M.A. Charpentier, M.M. Chintala, and G.B. Thursby. 2003. Relationships of nitrogen loadings, residential development, and physical characteristics with plant structure in New England salt marshes. Estuaries 26: 1494–1504.
Zedler, J.B. 1982. The ecology of southern California coastal salt marshes. US Fish and Wildlife Service Biological Report. Washington, D.C.
Zedler, J.B., J.C. Callaway, J.S. Desmond, G. Vivian-Smith, G.D. Williams, G. Sullivan, A.E. Brewster, and B.K. Bradshaw. 1999. Californian salt-marsh vegetation: an improved model of spatial pattern. Ecosystems 2: 19–35.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Weilhoefer, C.L., Nelson, W.G., Clinton, P. et al. Environmental Determinants of Emergent Macrophyte Vegetation in Pacific Northwest Estuarine Tidal Wetlands. Estuaries and Coasts 36, 377–389 (2013). https://doi.org/10.1007/s12237-012-9569-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-012-9569-9