
Relationship between Use Value and Ecological Importance of Floristic Resources of Seasonally Dry Tropical Forest in the Balsas River Basin, México. The use of seasonally dry tropical forest in the Balsas River Basin, Mexico was utilized as a model to study the relationship between the use value and the ecological importance of wild flora in nine study sites that have an indigenous-Mestizo cultural gradient. Ethnobotanical data from the area were used to obtain the use value per species and correlate it with the ecological importance value of the species found in 0.9 ha. We test the hypothesis that the most used species are the most ecologically important and that this relationship is affected by the changing cultural process that occurs in Mexican rural communities. We registered 320 species, 56.25 % of which are useful and fall into twelve use categories. The study sites showed similar percentages of useful plants. We found relationships between the use value of the species and of the main categories (medicinal, construction, firewood, and food) with the ecological importance value. The frequency was the ecological attribute that determined the use of species in the main categories of use, while the density and dominance attributes were related respectively to the medicinal and food categories. There were differences regarding cultural groups. Indigenous groups mentioned the use of medicinal, edible, and firewood plants more frequently, whereas the Mestizo group pointed out more species used for construction.
Relación entre valor de uso e importancia ecológica de los recursos vegetales del bosque estacional seco en la cuenca del Rio Balsas, Mexico. El uso de la bosque tropical caducifolio de la Cuenca del Río Balsas, México, se utilizó como modelo para estudiar la relación entre el valor de uso y la importancia ecológica de la flora silvestre de nueve sitios de estudio que tienen un gradiente cultural indígena-mestizo. Se consideró información etnobotánica del área para estimar el valor de uso de las especies y se relacionó con el valor de importancia ecológica de las especies encontradas en 0.9 ha. Ponemos a prueba la hipótesis de que las especies más utilizadas son las de mayor importancia ecológica y que esta relación se ve afectada por el proceso de cambio cultural que se presenta en las comunidades rurales mexicanas. Se registraron 320 especies, 56.25 % de ellas son útiles y corresponden a 12 categorías de uso. Los sitios de estudio mostraron el mismo porcentaje de plantas útiles. Encontramos relación entre el valor de uso de las especies y de las principales categorías (medicinal, construcción, leña y alimentos) con el valor de importancia ecológica de las diferentes especies. La frecuencia fue el atributo ecológico que determinó el uso de las especies en las principales categorías, mientras que los atributos de densidad y dominancia se relacionan con las categorías de medicina y alimentos, respectivamente. Hubo diferencias con respecto a los grupos culturales, los grupos indígenas mencionan más especies con usos medicinal, alimenticio y para leña, mientras que el grupo mestizo señaló más especies usadas para construcción. En términos generales, las especies más notables son las más frecuentemente mencionadas como útiles.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
What makes some plant species more important to people than others? Are there patterns of useful plants selection by humans? These are central questions in ethnobotany that have not been thoroughly answered. A number of scholars have addressed these questions from either biological or cultural perspectives. The influence of factors such as culture, palatability, smell, visibility, or quality of the plant have been widely stressed (Hanazaki et al. 2010; Holbrook et al. 1995; Pieroni 2001; Ramos et al. 2008; Turner 1988). Cultural consensus based mostly on the number of uses recorded for a plant species independently of its frequency of utilization have also been investigated (Phillips and Gentry 1993a, 1993b).
The importance of ecological variables related to the use of plants has been pointed out by other authors, including Phillips and Gentry (1993b), Galeano (2000), Cunha and Albuquerque (2006), and Thomas et al. (2009), and it has been often presumed that plant density or frequency determines plant accessibility to people. It has also been suggested that ecological salience of a species—for example, abundance, height, and diameter at breast height—have also influenced its recognition by humans (Hanazaki et al. 2010). A recent approach suggests the existence of a close relationship between the use value of a species and its ecological importance value (Cunha and Albuquerque 2006).
Studies about knowledge, use, and management of plant diversity in Mexico and other world regions show the existence of common patterns that local populations employ to select useful plants, and to classify and rank these plant resources (Berlin and Berlin 2005; Caballero et al. 2001; Moerman et al. 1999). However, Albuquerque et al. (2005) and Lucena et al. (2007) reported variations in the characteristics that affect patterns of use between communities, even in areas of relatively close proximity. These differences in use patterns appear to be due to the heterogeneity of the flora of tropical ecosystems, and so are only valid within specific biocultural contexts. These patterns may be affected by the nature of the cultural processes of knowledge acquisition and transference, as well as by the local characteristics of the flora and the surrounding environment.
Based on the ecological apparency hypothesis for explaining herbivore behavior (Feeny 1976; Rhoades and Cates 1976), and its adaptation to ethnobotany by Phillips and Gentry (1993a, 1993b), various scholars have recorded human uses and collected the most abundant and conspicuous useful species (Albuquerque and Lucena 2005; Kristensen and Balslev 2003; Lawrence et al. 2005; Lucena et al. 2007; Thomas et al. 2009). The ecological apparency hypothesis has been tested in rainforests of South America and Mexico in different contexts (Cunha and Albuquerque 2006; Galeano 2000; Kristensen and Balslev 2003; Torre-Cuadros and Islebe 2003), but dry forest plants and their uses have received little attention (Albuquerque et al. 2005; Lucena et al. 2007). Albuquerque et al. (2005) and Lucena et al. (2007) suggested that people may follow different strategies for using plant resources in arid and semiarid regions in comparison to those in rainforests, since in dry environments they depend on water availability. This has been recently shown by Albuquerque (2010) in the case of medicinal plants. However, more data are needed in other seasonally dry tropical forests to determine whether this is a common pattern.
Mexico is a culturally and biologically rich country where human-plant interactions have developed intricately over time and space (Caballero et al. 2001). The accelerating loss of ethnobiological knowledge by culture change raises the need for studies of specific ecological factors that relate to the uses of plants on the local, regional, and national scale.
Seasonally dry tropical forests (SDTF) are the most representative of Mexico’s ecosystems (Dirzo and Ceballos 2010). These forests have been strongly affected by deforestation (Trejo and Dirzo 2000), and Murphy and Lugo (1986) consider the affinity of humans for these areas as a possible cause of extensive damage to their communities. For these authors, low canopy height facilitates clearings for conversion to agricultural land. Fertile soils and a relatively benign climate have made dry forest more suitable for livestock activities than those in tropical rain forests.
The interaction of human groups with SDTF in Mexico dates back to 6000–8000 B.C.E. During the Classical and Postclassical periods, several pre-Hispanic groups settled in the SDTF (Balvanera and Maass 2010). Currently, of the 54 ethnic groups known for Mexico, 23 live near or in the seasonally dry tropical forests, and those groups have shown a deep understanding of their environment (Bye 1995). More than 600 plant species have been reported to be used for medicine, food, construction, and many other purposes (Bye 1995; Maldonado 1997; Soto 2010). The SDTF is the vegetation type in which the population utilizes the largest proportion of its plant species (Maldonado 1997; Soto 2010).
The role of ecological factors in Mexican dry forest plant use has been investigated by several authors (Casas et al. 1996; Maldonado 1997; Monroy-Ortiz and Monroy 2004), but only in a limited capacity. This paper analyzes the ethnobotanical relationship between use value (UV) and ecological importance value (EIV) by means of a case study of seasonally dry tropical forests in Mexico’s Balsas River Basin. We test the hypothesis that the most commonly used species are the most conspicuous regardless of the floristic heterogeneity of these forests. We also propose that this relationship is affected by the changing cultural situation that is occurring in Mexican rural communities.
Methods
Study Area
The study was conducted in the Balsas River Basin (BRB), a depression with an east-west direction, in the southern central part of Mexico that includes parts of the states of Tlaxcala, Puebla, Oaxaca, Estado de Mexico, Guerrero, Michoacán, Jalisco, and the entire state of Morelos. Just over 20 % of the country´s forest cover is found on the BRB, of which seasonally dry tropical forests is the most common (Rzedowski 1978).
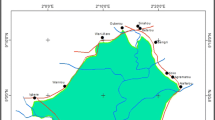
Nine sites that belong to the states of Guerrero, Morelos, and Puebla were selected mainly for their demographic composition and proximity to less disturbed SDTFs: Teocalcingo, Mezquitlan, and Temaxcalapa in Guerrero; El Limón de Cuauchichinola, Santa Catarina, and Cuentepec in Morelos; and Quetzotla, Xochitepec, and Huehuepiaxtla in Puebla. All sites are situated in the upper and middle regions of the BRB (Fig. 1). Formerly inhabited by Olmecs, at present the region is occupied by Mestizos and indigenous Nahuatl speakers. These groups, in combination with the SDTF, comprise a biocultural matrix whose primary productive activities are rain-fed agriculture, cattle ranching, and extraction of timber and non-timber products.

Geographical location of the study area in the Balsas River Basin, México; the BRB has traditionally been divided as upper, middle, and lower regions, our study sites are located in the upper and middle regions, comprising localities in the states of Guerrero, Morelos, and Puebla.
Selection of Sampling Sites
We chose seasonally dry tropical forest sites that were perceived by the researchers and the local residents to be the least disturbed, so as to eliminate as much as possible the effect of ongoing human vegetation management. Human populations were selected that were in close proximity to the vegetated study sites. Selected sites represented a gradient of cultural change with varying proportions of indigenous and Mestizo individuals. Three main groups were recognized. The first group comprises indigenous villages where over 50 % of the population speaks Nahuatl and whose settlement dated from pre-Hispanic times. Less than 50 % of the second group speaks the language, but the settlement has existed for over 100 years. The third group includes towns less than 100 years old and whose inhabitants speak only Spanish. Nine localities, three sites in each group, were studied (Table 1).
Approval from the local authorities was obtained before the study began. Once the participation of the community was approved, the sampling area was jointly defined by researchers, local authorities, and owners of the chosen properties. The active participation of experienced informants, chosen by the community, was also agreed upon.
Estimating the Ecological Importance Value of the Species
Ten 50 × 2.0 m plots (0.1 ha each), distributed within the area occupied by the seasonally dry tropical forest, were established in the nine sites (0.9 ha in total). The information gathered in these plots was the diameter at breast height (DBH) (>1.0 cm of all woody individuals) and the basal diameter (BD) of all lianas rooted within the plot (Gentry 1982, 1988, modified by Trejo and Dirzo 2002). Data on the basic structural parameters, density, dominance, and frequency were also recorded. Using the relative value of these parameters, we estimated the ecological importance value (EIV) according to Mueller-Dumbois and Ellenberg (1974), by using the following formula,
A matrix of presence and absence and a similarity analysis (Jaccard index) were developed for each of the species using the floristic richness from each site. Botanical specimens collected in each of the sites were taxonomically identified and deposited in the HUMO Herbarium (Universidad Autónoma del Estado de Morelos) and in MEXU Herbarium (Universidad Nacional Autónoma de Mexico).
Estimating the Use Value
In each of the study sites we selected six experienced informants (54 in total), all native to the study area. All were men over 40 years old who, according to the community, had experience in the use of plants and were engaged in primary productive activities.
The informants were asked about Spanish and/or Nahuatl common names of each plant, as well as their use(s) from a random sample of plant individuals in the forest plots. With this information and the use of the algorithm proposed by Phillips and Gentry (1993a, 1993b), as modified by Rossato et al. (1999), we calculated the use value (UV) of the species used. This index was estimated as the sum of the uses recognized by each informant for each species divided by the total number of informants according to the following formula: \( \mathrm{UVs}={{{\left( {\varSigma\,\mathrm{U}is} \right)}} \left/ {\mathrm{N}} \right.} \), where Uis equals the number of uses mentioned by informant i, for species s. N is the total number of informants interviewed for each species. Thus, the use value is the average of uses reported for each species by the 54 informants. For the purpose of the analysis, these species then were grouped according to their main use categories.
A non-parametric Kruskal-Wallis test was used to assess the differences between the number of reported species and the percentage of useful species, and a Mann-Whitney test was used to calculate the differences between number of species per use category among the most contrasting cultural groups (indigenous and Mestizo).
Use Value and Ecological Importance Value
The relationship between use value and ecological importance value was estimated by first doing a simple regression analysis using all the UV data as the responsive variable and the EIV of the species as the explanatory variable. This same comparison was done after separating the data by site. A multiple regression analysis was carried out to estimate the relationship between UV and EIV, but using the four use categories listed as the most important according to the number of useful species. A multiple regression analysis was used to assess the contribution of each ecological attribute (frequency, density, and dominance) to the UV by category.
A linear model of mixed effects was used to determine if the UV is affected by the EIV in the different communities studied. The fixed components of the model were the cultural groups; the random components were the EIV variables for each site. The UV and the EIV were transformed to their natural logarithm to normalize the residuals. The lme procedure (Pinheiro et al. 2009) of the R program version 2.8.1 (R Development Core Team 2009) with the maximum verisimilitude option was used to adjust the mixed model. The statistical significance of the terms of the fixed model was tested with the χ2 values, and the differences in degree of freedom were obtained by comparing the adjustment between the more complex models with the simplest one.
Results
Floristic Richness and Similarity between Sites
There were a total of 320 species belonging to 187 genera and 61 botanical families in the nine sites. According to the number of species, the most representative families were: Fabaceae (51), Burseraceae (20), Asteraceae (14), Euphorbiaceae (10), Malpighiaceae (9), Cactaceae (8), Convolvulaceae (8), Anacardiaceae (7), Apocynaceae (7), and Rubiaceae (6). The nine sites were floristically very heterogeneous, and the similarity analyses revealed that only sites I and VII were similar, sharing 35 % of species. Additionally, all sites shared only five species, three trees—Bursera copallifera (DC.) Bullock, Comocladia engleriana Loes, Lysiloma divaricatum (Jacq.) J.F. Macbr., and two lianas—Cissus sicyoides L. and Serjania triquetra Radlk.
Of the species recorded, 180 (56.25 %) were useful plants corresponding to twelve use categories. Of these, 94 fell into a single use category and 86 had multiple uses. The most important use categories from the number of species reported for all sites were as follows: medicinal (48.8 %), construction (34.4 %), firewood (25 %), and food (17.7 %).
The Kruskal-Wallis test showed no significant differences between sites (p < 0.63) compared to the number of species reported per site and to the number of species listed as useful per cultural group. Each case involved the use of about 60 % of the species found. The same was true when comparing the number of categories present per site using this same test. However, when considering use categories and the two contrasting groups, the Mann-Whitney test showed significant differences between groups (p < 0.05) in the medicinal, construction, firewood, and food use categories. Indigenous peoples mentioned more species for medicine, food, and firewood than the Mestizo informants, whereas the latter reported more species for construction than the indigenous informants (Table 2).
Use Value and Ecological Importance Value of Species
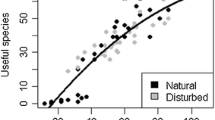
The regression of UV related to the EIV of each species per site showed a significant relationship (p < 0.05) between both factors in only four of the nine sites, three belonging to the indigenous group and one to the Mestizo group (Table 3). In addition, when all sites were analyzed together, a significant relationship was found between the UV and the EIV of all useful species and the four main use categories (Fig. 2).

Linear regression between use values (UV) and ecological importance values (EIV) of useful species in seasonally dry tropical forest in the Balsas River Basin.
The multiple regressions between the UV per category and the relative values of frequency, density, and dominance of the species showed a significant relationship for species frequency in all categories, while the attributes of density and dominance were only significant for the medicinal plants and edible plants, respectively (Table 4).
The ten species with the highest EIV were Lysiloma divaricatum, Bursera copallifera, Conzattia multiflora (B.L. Rob.) Standl., Euphorbia schlechtendalii Boiss., Heliocarpus terebinthinaceus (DC.) Hochr., Serjania triquetra, Pseudosmodingium perniciosum (Kunth) Engl., Amphipterygium adstringens (Schltdl.) Standl., Ceiba aesculifolia (Kunth) Britten & Baker, and Cyrtocarpa procera Kunth. The species with the highest UV were Bursera copallifera, Lysiloma divaricatum, Haematoxylum brasiletto H. Karst., Lysiloma acapulcense (Kunth) Benth., Leucaena esculenta (Moc. & Sessé ex DC.) Benth., Plumeria rubra L., Senna skinneri (Benth.) H.S. Irwin & Barneby, Eysenhardtia polystachya (Ortega) Sarg., Bursera bipinnata (DC.) Engl., and Guazuma ulmifolia Lam.. The five species common to all sites were reported as utilized for the same purposes. The list of 180 species recorded in the nine sites and their respective density, frequency, and dominance values, as well as their ecological importance and use values are listed in Appendix 1. (Electronic Supplementary Material).
The mixed model results showed no significant interaction between the degrees of cultural change (indigenous, Mestizo, and heterogeneous groups) and the EIV (χ2 = 1.81, g.1 = 1, p = 0.40), so they were removed from the model. The simplified model indicated a highly significant positive relationship between the EIV and the UV (χ 2 = 22.00, d.f. = 1. p < 0.001), as well as a marginally significant difference between cultural groups and UV (χ2 = 5.37, d.f. = 1, p = 0.058).
Discussion and Conclusion
Our results support the hypothesis that the seasonally dry tropical forest species most used by people in the Balsas River Basin are those that are ecologically most important. This is true despite the high β diversity of these forests and the fact that there are few species with high ecological importance (Trejo 2005). No differences were observed in the number of species mentioned by indigenous and Mestizo peoples. There were differences between the two groups, however, in terms of use categories. Indigenous people reported more species used as medicine, food, and firewood than Mestizos, whereas the latter reported more species used for construction. This is similar to the results reported by Lawrence et al. (2005) for a tropical rain forest use in Peru. In dry and humid forests, Albuquerque et al. (2005) and Cunha and Albuquerque (2006) reported that more than half of the species (52.25 %) were useful, and that the most important use categories were medicinal, construction, firewood, and food. The results of the present study are also consistent with those reported by Bye (1995) for this forest type in Mexico. Other authors (Galeano 2000; Torre-Cuadros and Islebe 2003) found that in humid forests the larger number of useful plants were those used for construction. Similarly, species with multiple uses were fewer than those with a single use in both humid and dry forests of Mexico (Caballero et al. 2001; Toledo et al. 2003; Torre-Cuadros and Islebe 2003). Moreover, other studies demonstrated that multiple-use species are culturally more important (Caballero et al. 2001; Casas et al. 1996; Phillips and Gentry 1993a, 1993b; Pieroni 2001; Turner 1988), and our results show that multiple-use species also possess higher EIV values.
The most important families (in terms of number of species) in this study are the same ones shown by floristic studies in other Mexican SDTFs—Fabaceae, Burseraceae, Asteraceae, Euphorbiaceae, Cactaceae, and Anacardiaceae (Lott and Atkinson 2006; Trejo and Dirzo 2002). These have also been reported as most frequently used species in the habitat (Bye 1995). The Fabaceae family, for example, is the most speciose family in this type of vegetation and it has the highest number of useful species. These species are mainly used in traditional house construction, manufacture of farm tools, and as firewood, as noted elsewhere by Lucena et al. (2007) in Brazilian Caatinga, and by Casas et al. (1996) and Maldonado (1997) in the Balsas River Basin. The Burseraceae family, represented by the genus Bursera, is also widely used, with 13 of its 20 reported species having one or more uses. This is in agreement with Lucena et al. (2007), who reported that plant use value (UV) is strongly related to the botanical family.
The results of this study also suggest that the use of a given species may be an important factor in the UV-EIV relationship. The species with the highest UV and EIV were Lysiloma divaricata and Bursera copallifera, which were also the most widely distributed species in the study region. As suggested by Benz et al. (1994), their abundance is a result of human disturbance. During our fieldwork we observed regenerating stumps of L. divaricata in sites that have been managed, thus favoring the presence of the species. Moreover, resin extraction from B. copallifera does not damage the trees, so their density does not decrease. The only exception to this is Conzattia multiflora, which, despite exhibiting high EIV, presents a very low UV. Although this species is abundant in the SDTF, it is said to have very brittle wood and therefore is not often used.
When the main use categories on a regional level were analyzed, we found a significant relationship between the UV and the EIV in the three sites with indigenous peoples and only one site with Mestizo people. This suggests that culture has a differential impact on selection patterns and resource use according to use categories. Indigenous groups use more species as medicine, food, and firewood, whereas Mestizo groups tend to use more species for construction (c.f. Berlin and Berlin 2005; Caballero et al. 2001; Lawrence et al. 2005; Toledo et al. 2003), which could be due to the latter group’s knowledge still being “general.” This possibly was the case in the single Mestizo group with a significant relationship between UV and EIV, which may be going through a specialization process. It could also reflect the strong interest in environmental conservation that these people have developed during the last decades. In other Mexican Mestizo groups, change in the primary productive activity toward secondary and tertiary activities, plus a higher degree in formal education, could be eroding their traditional ecological knowledge (see also González-Insuasti and Caballero 2007 and Martínez-Ballesté et al. 2006).
The species’ frequency was the attribute that best predicted UV in the four main categories—medicine, construction, firewood, and food. Density and dominance were respectively significant only for medicinal and food species. This is in agreement with the findings in humid forests by Thomas et al. (2009) and Torre-Cuadros and Islebe (2003), which showed that density, frequency, and DBH are significantly correlated with the same use categories. In contrast, our results were not consistent with those of Lucena et al. (2007) in the Brazilian Caatinga, where frequency is associated only with the medicinal category, while dominance is related to the construction category. Furthermore, Albuquerque et al. (2005) found that in dry forest the most important useful plants to local people are not necessarily the most abundant or ecologically important.
In this study, frequency is the best predictor of the use value of plants. This could be because of the heterogeneity of these forests and because related species of the same taxon have a high likelihood of co-occurring in the same narrow geographical environment (Lavin 2006). This translates to increased availability, reducing the investment in time and effort when harvesting, especially in regards to timber extraction. Because species used as food are mostly trees (75 %) whose fruits are gathered, they need to be easily found, which could explain why dominance is also a significant factor in this use category. The strong relationship between food use and dominance may also be explained by the fact that larger trees produce more edible fruits and are thus more highly valued by people (Thomas et al. 2009). In the case of medicinal plants in which the relationship between density and the UV was also significant, we suggest that a higher frequency together with a greater density of individuals of these species could facilitate harvesting time and consequently increase the total amount of harvested product. Other studies in the SDTF have already suggested that the use of plants depends on their relative density (Benz et al. 1994), but this had not been tested for each of the use categories.
This study shows that regardless of sociocultural differences, the most important useful plants for local populations are also the most ecologically important, especially those with the highest frequency. The highest ecological importance value of these species seems to be the end result of their evolutionary history, making them well adapted to colonize areas where seasonal drought is long and pronounced (Holbrook et al. 1995; Murphy and Lugo 1986; Trejo 2005). This favors a wide geographical distribution and allows these species to respond positively to human disturbance (Benz et al. 1994).
Based on current evidence from different regions of the Americas (Balée 1998; Clement 1999; Fedick 1996; Peters 2000), we suggest that the highest ecological importance of useful plants in the SDTF may not be only an ecological attribute of usefulness, but may also be a result of forest management and landscape transformation by humans over long periods of time. Although Murphy and Lugo (1986) and Trejo and Dirzo (2000) have stressed the negative impact of humans on these forests and that the long history of plant management of this vegetation may have induced significant modifications in forest composition, this remains to be investigated.
Literature Cited
Albuquerque, U. P. 2010. Implications of ethnobotanical studies on bioprospecting strategies of new drugs in semi-arid regions. The Open Complementary Medicine Journal 2:21–23.
——— and R. F. P. Lucena. 2005. Can apparency affect the use of plants by local people in tropical forest? Interciencia 30(8):506–511.
———, L. H. C. Andrade, and A. C. O. Silva. 2005. Use of plant resources in a seasonal dry forest (Northeastern Brazil). Acta Botanica Brasilica 19:27–38.
Balée, W. 1998. Historical ecology: Premises and postulates. In: Advances in historical ecology, ed. W. Balée, 13–29. New York: Columbia University Press.
Balvanera, P. and M. Maass. 2010. Los servicios ecosistémicos que proveen las selvas secas. In: Diversidad, amenazas y áreas prioritarias para la conservación de las selvas del Pacífico de México, eds. G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury Creel, and R. Dirzo, 251–269. CONABIO (National Commission for Knowledge and Use of Biodiversity) / FCE, México.
Benz, B. F., F. Santana, R. Pineda, J. Cevallos, L. Robles, and D. DeNiz. 1994. Characterization of Mestizo plant use in the Sierra de Manantlán, Jalisco-Colima. Mexico. Journal of Ethnobiology 14(1):23–41.
Berlin, B., and E. A. Berlin. 2005. Conocimiento indígena popular: La flora común, herbolaria y salud en los altos de Chiapas. In: Diversidad biológica en Chiapas, eds. N. Ramírez-Marcial and L. Ruiz-Montoya, 371–418. ECOSUR (El Colegio de la Frontera Sur), COCYTECH (Consejo de Ciencia y Tecnología de Chiapas), Plaza y Valdés, S.A. de C.V., México City.
Bye, R. 1995. Ethnobotany of the Mexican dry tropical forests. In: Seasonally dry tropical forests, eds. S. H Bullock, H. A. Mooney, and E. Medina, 423–438. Cambridge, U.K.: Cambridge University Press.
Caballero, J., A. Casas, L. Cortés, and C. Mapes. 2001. Patrones en el conocimiento, uso y manejo de las plantas en pueblos indígenas de México. Estudios Atacameños 16:181–196.
Casas, A., M. C. Vázquez, J. L. Viveros, and J. Caballero. 1996. Plant management among the Nahua and the Mixtec in the Balsas River Basin, Mexico: An ethnobotanical approach to the study of plant domestication. Human Ecology 24:455–478.
Clement, C. R. 1999. 1492 and the loss of Amazonian crop genetic resources. II. Crop biogeography at contact. Economic Botany 53:203–216.
Cunha, L. V. F. and U. P. Albuquerque. 2006. Quantitative ethnobotany in an Atlantic forest fragment of northeastern Brazil: Implications to conservation. Environmental Monitoring and Assessment 114:1–25.
Dirzo, R. and G. Ceballos. 2010. Las selvas secas de México: Un reservorio de biodiversidad y laboratorio viviente. In: Diversidad, amenazas y áreas prioritarias para la conservación de las selvas del Pacífico de México, eds. G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury Creel, and R. Dirzo, 13–20. CONABIO (National Commission for Knowledge and Use of Biodiversity) / FCE, México.
Fedick, L. S. 1996. New perspectives on ancient Maya agriculture and resource use. In: The managed mosaic: Ancient Maya agriculture and resource use, ed. L. S. Fedick, 1–14. Salt Lake City: University of Utah Press.
Feeny, P. 1976. Plant apparency and chemical defense. In: Biological interactions between plants and insects. Recent advances in phytochemistry 10, eds. J. W. Wallace and R. L. Nansel, 1–40. New York: Plenum Press.
Galeano, G. 2000. Forest use at the Pacific coast of Chocó, Colombia: A quantitative approach. Economic Botany 54:358–376.
Gentry, A. H. 1982. Patterns of neotropical plant species diversity. Evolutionary Biology 15:1–54.
———. 1988. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden 75:1–34.
González-Insuasti, M. S. and J. Caballero. 2007. Managing plant resources: How intensive can it be? Human Ecology 35:303–314.
Hanazaki, N., R. Mazzeo, A. R. Duarte, V. C. Souza, and R. R. Rodrigues. 2010. Ecologic salience and agreement on the identification of tree species from Brazilian Atlantic Forest. Biota Neotropica 10(1):77–84.
Holbrook, N. M., J. L. Whitbeck, and H. A. Mooney. 1995. Drought responses of neotropical deciduous forest trees. In: Seasonally dry tropical forests, eds. S. H. Bullock, H. A. Mooney, and E. Medina, 243–276. Cambridge, U.K: Cambridge University Press.
Kristensen, M. and H. Balslev. 2003. Perceptions, use and availability of woody plants among the Gourounsi in Burkina Faso. Biodiversity and Conservation 12:1715–1739.
Lavin, M. 2006. Floristic and geographical stability of discontinuous seasonally dry tropical forests explain patterns of plant phylogeny and endemism. In: Neotropical savannas and seasonally dry tropical forests. Plant diversity, biogeography, and conservation, eds. R. T. Pennington, G. P. Lewis, and J. A. Ratter, 433–448. The Systematics Association Special Volume Series 69. Boca Raton, Florida: CRC Press, Taylor and Francis Group.
Lawrence, A., O. L. Phillips, A. Reategui, M. Lopez, S. Rose, D. Wood, and A. J. Farfan. 2005. Local values for harvested forest plants in Madre de Dios, Peru: Towards a more contextualized interpretation of quantitative ethnobotanical data. Biodiversity and Conservation 14:45–79.
Lott, E. J. and T. H. Atkinson. 2006. Mexican and Central American seasonally dry tropical forests: Chamela-Cuixmala, Jalisco, as a focal point for comparison. In: Neotropical savannas and seasonally dry tropical forests. Plant diversity, biogeography, and conservation, eds. R. T. Pennington, G. P. Lewis, and J. A. Ratter, 315–342. The Systematics Association Special Volume Series 69. Boca Raton, Florida: CRC Press, Taylor and Francis Group.
Lucena, R. F. P., E. L. Araújo, and U. P. Alburquerque. 2007. Does the local availability of woody Caatinga plants (Northeastern Brazil) explain their use value? Economic Botany 61(4):347–361.
Maldonado, B. 1997. Aprovechamiento de los recursos florísticos en la Sierra de Huautla, Morelos. Tesis de Maestría. Ciudad de México, México: Facultad de Ciencias, Universidad Nacional Autónoma de México.
Martínez-Ballesté, A., C. Martorell, and J. Caballero. 2006. Cultural or ecological sustainability? The effect of cultural change on Sabal palm management among the lowland Maya of Mexico. http://www.ecologyandsociety.org/vol11/iss2/art27/.
Moerman, D. E., R. W. Pemberton., D. Kiefer, and B. Berlin. 1999. A comparative analysis of five medicinal floras. Journal of Ethnobiology 19:49–67.
Monroy-Ortiz, C. and R. Monroy. 2004. Análisis preliminar de la dominancia cultural de las plantas útiles en el estado de Morelos. Boletín de la Sociedad Botánica de México 74:77–95.
Mueller-Dumbois, D. and H. Ellenberg. 1974. Aims and methods of vegetation ecology. New York: John Wiley and Sons.
Murphy, P. G. and A. Lugo. 1986. Ecology of tropical dry forest. Annual Review of Ecology and Systematics 17:67–88.
Peters, M. C. 2000. Precolumbian silviculture and indigenous management of neotropical forests. In: Imperfect balance: Landscape transformations in the pre-Columbian Americas, ed. L. D. Lentz, 203–223. New York: Columbia University Press.
Phillips, O. and A. H. Gentry. 1993a. The useful plants of Tambopata, Peru: I. Statistical hypothesis test with a new quantitative technique. Economic Botany 47:15–32.
——— and ———. 1993b. The useful plants of Tambopata, Peru: II. Additional hypothesis-testing in quantitative ethnobotany. Economic Botany 47(1):33–43.
Pieroni, A. 2001. Evaluation of the cultural significance of wild food botanicals traditionally consumed in northwestern Tuscany, Italy. Journal of Ethnobiology 21:89–104.
Pinheiro, J., D. Bates, S. DebRoy., D. Sarkar, and the R Core Team. 2009. Nlme: Linear and nonlinear mixed effects models. R package version 3.1–91.
Ramos, M. A., P. M. Medeiros, A. L. S. Almeida, A. L. P. Feliciano, and U. P. Albuquerque. 2008. Can wood quality justify local preferences for firewood in an area of Caatinga (dryland) vegetation? Biomass and Bioenergy 32:503–509.
Rhoades, D. F. and R. G. Cates. 1976. Towards a general theory of plant antiherbivore chemistry. In: Biological interactions between plants and insects. Recent advances in phytochemistry 10, eds. J. W. Wallace, and R. L. Nansel, 169–213. New York: Plenum Press.
Rossato, S., H. F. Leitáo-Filho, and A. Begossi. 1999. Ethnobotany of Caicaras of the Atlantic Forest Coast (Brazil). Economic Botany 53:387–395.
Rzedowski, J. 1978. Vegetación de México. México: Editorial Limusa.
Soto, J. C. 2010. Plantas útiles de la cuenca del Río Balsas. In: Diversidad, amenazas y áreas prioritarias para la conservación de las selvas del Pacífico de México, eds. G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury Creel, and R. Dirzo, 285–320. CONABIO (National Commission for Knowledge and Use of Biodiversity) / FCE, México.
Thomas, E., I. Vandebroek, and P. Van Damme. 2009. Valuation of forest and plant species in indigenous territory and National Park Isiboro-Sécure, Bolivia. Economic Botany 63:229–241.
Toledo, V. M., B. Ortíz-Espejel., L. Cortés, P. Moguer, and M. de J. Ordoñez. 2003. The multiple use of tropical forest by indigenous peoples in Mexico: A case of adaptive management. Conservation Ecology 7(3):9.
Torre-Cuadros, M. D. and G. A. Islebe. 2003. Traditional ecological knowledge and use of vegetation in southeastern Mexico: A case study from Solferino, Quintana Roo. Biodiversity and Conservation 2(12):2455–2476.
Trejo, I. 2005. Análisis de la diversidad de la selva baja caducifolia en México. In: Sobre diversidad biológica: El significado de las Diversidades Alfa, Beta y Gamma, eds. G. Halffter, J. Soberón, P. Koleff and A. Melic, 111–122. SEA (Sociedad Entomológica Aragonesa), CONABIO (National Comission for Knowledge and Use of Biodiversity, CONACYT (Consejo Nacional de Ciencia y Tecnología) y Grupo Diversitas. Zaragoza, España.
——— and R. Dirzo. 2000. Deforestation in seasonally dry tropical forests: A national and local analysis in Mexico. Biological Conservation 94:133–142.
——— and ———. 2002. Floristic diversity of Mexican seasonally dry tropical forests. Biodiversity and Conservation 11:2063–2084.
Turner, N. J. 1988. The importance of a rose: Evaluating the cultural significance of plants in Thompson and Lillooet Interior Salish. American Anthropologist 90:272–290.
Acknowledgments
This article fulfills a partial requirement for the first author for obtaining a Ph.D. degree. The first author thanks the Posgrado en Ciencias Biológicas of the Universidad Nacional Autónoma de México and the Programa de Mejoramiento del Profesorado of the Universidad Autónoma del Estado de Morelos for their support with the scholarship to pursue postgraduate studies. Thanks to Dr. Nestor Mariano, Dr. Andrea Martínez-Ballesté, and Alfredo Saynes for his support in the statistical analysis and manuscript reviewing. Thanks to the editor and two anonymous reviewers for suggestions that greatly improved this article. To our colleagues at CIByC-UAEM for their support in the field-work. To the technicians and specialists at the Herbarium MEXU-UNAM and HUMO-UAEM for their support in the identification of botanical material. To the authorities, inhabitants, and informants in the different localities studied for their collaboration in the field work and for their hospitality.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 120 kb)
Rights and permissions
About this article
Cite this article
Maldonado, B., Caballero, J., Delgado-Salinas, A. et al. Relationship between Use Value and Ecological Importance of Floristic Resources of Seasonally Dry Tropical Forest in the Balsas River Basin, México. Econ Bot 67, 17–29 (2013). https://doi.org/10.1007/s12231-013-9222-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12231-013-9222-y