
Abstract
Heat shock proteins (HSPs) are molecular chaperones subdivided into several families based on their molecular weight. Due to their cytoprotective roles, these proteins may help protect cancer cells against chemotherapy-induced cell death. Investigation into the biologic activity of HSPs in a variety of cancers including primary bone tumors, such as osteosarcoma (OSA), is of great interest. Both human and canine OSA tumor samples have aberrant production of HSP70. This study assessed the response of canine OSA cells to inhibition of HSP70 and GRP78 by the ATP-mimetic VER-155008 and whether this treatment strategy could sensitize cells to doxorubicin chemotherapy. Single-agent VER-155008 treatment decreased cellular viability and clonogenic survival and increased apoptosis in canine OSA cell lines. However, combination schedules with doxorubicin after pretreatment with VER-155008 did not improve inhibition of cellular viability, apoptosis, or clonogenic survival. Treatment with VER-155008 prior to chemotherapy resulted in an upregulation of target proteins HSP70 and GRP78 in addition to the co-chaperone proteins Herp, C/EBP homologous transcription protein (CHOP), and BAG-1. The increased GRP78 was more cytoplasmic in location compared to untreated cells. Single-agent treatment also revealed a dose-dependent reduction in activated and total Akt. Based on these results, targeting GRP78 and HSP70 may have biologic activity in canine osteosarcoma. Further studies are required to determine if and how this strategy may impact the response of osteosarcoma cells to chemotherapy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Osteosarcoma (OSA) is the most common primary bone tumor of both man and dogs. In both species, OSA has a locally aggressive behavior and high rate of systemic metastasis to organs such as the lungs. Naturally occurring OSA of pet dogs is a good model of this disease in man. Both are treated with surgical removal of the primary tumor and systemic chemotherapy to prevent metastasis. Standard of care chemotherapy regimens for canine OSA often encompass the administration of doxorubicin, carboplatin, or cisplatin. Regardless of the treatment protocol or chemotherapeutic agents utilized, canine OSA patients typically have a median survival time post-surgery of approximately 300 days (Berg et al. 1997; Bergman et al. 1996; Straw et al. 1991). Despite chemotherapy treatment, pulmonary metastasis remains highly prevalent, is significantly associated with a poor prognosis, and is the cause of euthanasia in the vast majority of canine OSA cases (Khoury et al. 2014; Mauldin et al. 1988). As clinical responses to conventional chemotherapy have reached a plateau, newer approaches involving targeted therapies, alone or in combination with cytotoxic chemotherapy, are desperately needed to make improvements in outcome.
Heat shock proteins (HSPs) are molecular chaperones subdivided into several families based on their molecular weight: HSP40, HSP60, HSP70, HSP90, HSP100, and small HSPs (Hartl et al. 2011). HSPs are found in organisms ranging in complexity from yeast and bacteria, to humans, and have been associated with ubiquitous functions (Kregel 2002). HSPs, such as those from the 70 kDa family, are known to complete protein translocation, maturation, folding, and degradation duties within the cell under both stressed and non-stressed states (Feder and Hofmann 1999). Acting in this fashion, HSP70s intervene in the apoptotic cascade and prevent protein aggregation, ultimately hindering apoptosis by an ATP-dependent mechanism (Beere et al. 2000; Elmore 2007; Gabai et al. 2002; Gotoh et al. 2004; Gurbuxani et al. 2003; Li et al. 2010; Ruchalski et al. 2003; Zhuang et al. 2013).
Given the role of HSP70s in the prevention of apoptosis, their relative expression has been studied in numerous cancers. In human OSA, overexpression of HSP70 was observed in 47 % of samples (Uozaki et al. 2000). Expression levels of HSP70 were also associated with poor prognosis and larger tumor size (Uozaki et al. 2000). Adding to the notion that human and canine OSA is very similar, immunohistochemical staining of 18 canine tumor samples found that HSP70 expression was associated with the presence of a grade III tumor, and lack of HSP70 staining correlated with longer survival time (Romanucci et al. 2012).
In addition to expression and prognostic analyses, the utility of HSP70 as a therapeutic target in oncology has also been evaluated. Inhibitors of HSP70 family members have been shown to reduce cellular viability, increase apoptosis, and reduce clonogenic survival in a number of cancer cell types (Leu et al. 2009). One example of a small molecule HSP70 inhibitor is VER-155008, which targets the ATPase binding domain and has been shown to induce apoptosis in the lung, colon, and multiple myeloma cells (Chatterjee et al. 2013; Leu et al. 2009; Massey et al. 2010; Wen et al. 2014). In other studies, increased expression of HSP70 reduced the efficacy of HSP90 inhibitors at inducing apoptosis (Guo et al. 2005). Furthermore, following the use of an HSP90 inhibitor in canine OSA patients, HSP70 was overexpressed in tumor tissue (London et al. 2011), which is considered a hallmark of effective HSP90 inhibition. Despite these findings, we are unaware of studies attempting to inhibit HSP70 and/or GRP78 as a single or dual target in canine OSA. The purposes of this study were to investigate the biologic activity of an HSP70 and GRP78 inhibitor in vitro in canine OSA cell lines and evaluate this treatment strategy in combination with standard of care doxorubicin chemotherapy.
Materials and methods
Antibodies and reagents
Antibodies for western blot or immunofluorescent detection of β-actin, HSP70, GRP78, and HSP90 were purchased from Cell Signaling Technology. BAG-1 and α-tubulin antibodies were purchased from Epitomics and Sigma-Aldrich. Herp and C/EBP homologous transcription protein (CHOP) antibodies were obtained from Abcam. Alexa-Fluor 488 fluorescent secondary antibody was purchased from Life Technologies.
The HSP70 inhibitor VER155008 was obtained from Tocris Bioscience and resuspended in dimethyl sulfoxide (DMSO) at a 10 mM concentration. This stock solution was stored at −80 °C for long-term storage and kept at 4 °C during use. Doxorubicin was obtained from the Ontario Veterinary College Animal Cancer Centre Pharmacy and maintained at 4 °C, at a concentration of 3.68 mM. The annexin V and propidium iodide (PI) apoptosis reagents were obtained from BioVision. This kit included annexin V and PI antibodies, as well as the annexin V binding buffer. Premixed Clonotech WST-1 reagent was used for cell viability assays and purchased from Cedarlane.
Cell lines and culture conditions
The D17 canine OSA cell line, as described by Riggs et al. (1974), was generously provided by Sarah Wootton University of Guelph. The JL31 and Dharma canine OSA cell lines were each derived from the primary tumor of a canine OSA patient and adapted to culture by Geoffrey Wood and Anthony Mutsaers, University of Guelph, respectively. Cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10 % FBS and penicillin/streptomycin. All cell lines were maintained at 37 °C and 5 % carbon dioxide in a humidified incubator.
Viability assays
Cells were plated in 96-well dishes in triplicate and allowed to adhere for 24 h. They were then incubated with VER155008 (VER), an N-terminal HSP70 ATPase inhibitor, for 24 h before doxorubicin was added for an additional 48 h. After 48 h, WST-1 reagent was pipetted into the wells and absorbance readings were obtained at a wavelength of 450 nm.
Western blot analysis
Lysis buffer (20 mM Tris-HCL, 150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 2.5 mM sodium pyrophosphate, 1 mM β-glycerophosphate, 1 mM Na3VO4, and 1 μg/mL of leupeptin from Cell Signaling Technology) containing aprotinin, phenylmethanesulfonyl fluoride, and a phosphatase inhibitor cocktail was used to lyse cells. Total protein from the samples was quantified using a Bradford assay (Bradford 1976). Protein was separated by weight using Bio-Rad electrophoresis chambers and power systems with 7.5, 10, or 12 % sodium dodecyl sulfate polyacrylamide gels. The protein was then transferred to a polyvinyl difluoride membrane and imaged using a Bio-Rad ChemiDoc system.
Immunofluorescence
Following treatment, cells were fixed with 4 % paraformaldehyde and blocked. The primary antibodies were then diluted in antibody dilution buffer (5 % BSA, 0.33 % Triton-X 100, and PBS). Slides were incubated with the primary antibody overnight prior to DAPI being applied. The resulting slides were visualized using an Olympus IX83 confocal microscope.
Apoptosis
After 72 h of treatment, floating and adherent cells were collected from each treatment group. The cells were counted using a Countess Automated Cell Counter (Invitrogen), and an equal number of cells were taken from each group, centrifuged, and resuspended in annexin V binding buffer. Propidium iodide and annexin V were added to each sample in the dark for 15 min. The samples were then analyzed using a BD FACScan flow cytometer.
Clonogenic survival
After 72 h of treatment, floating and adherent cells were collected. The cells were centrifuged and resuspended in fresh media. The cells were counted and plated into 60-mm plates in limiting dilutions. Every 3–4 days, the media was changed. When the untreated control colonies were becoming too numerous to count, the cells were fixed with 20 % methanol and simultaneously stained with 0.5 % crystal violet. Colonies were counted manually. Successful colony formation was defined as a colony of at least 50 cells.
Statistical analysis
Minitab 17 software (Minitab Inc.) was utilized to conduct analysis of variance (ANOVA) and two sample t tests. Tukey post hoc tests were used at the 95 % confidence interval. Results are reported as mean ± standard error of the mean (SEM).
Results
Viability/apoptosis/clonogenic survival
In all three canine OSA cell lines, single-agent treatment with VER for 72 h significantly reduced cellular viability (Fig. 1). However, Dharma was the only cell line that displayed a significant reduction in cellular viability in a combination group when compared to doxorubicin alone.

Cellular viability of canine OSA cells following HSP70 inhibition in combination with doxorubicin. VER causes significant reductions in cellular viability. Combination treatment reduced cellular viability in excess of that achieved by chemotherapy alone. [Values are compared to DMSO controls. *P < 0.05, statistical significance from DMSO; # P < 0.05, statistical significance from doxorubicin, each based on three replicates and an ANOVA. Values represent mean ± SEM]
Given the inability of WST-1 assays to directly assess cell death by apoptosis, annexin V and PI flow cytometry was utilized to assess cellular death. D17, Dharma, and JL31 all showed an increase in the number of apoptotic cells, although for Dharma, it was primarily in early stages of apoptosis (lower right quadrant) following 72 h of VER treatment (Figs. 2, 3, and 4). However, no additive effects were observed in VER and doxorubicin combination treatment groups.

Apoptosis in JL31 OSA cells as determined by annexin V/PI flow cytometry. VER155008 treatment produced apoptosis. Combination treatments did not produce additive effects. Cells were incubated for 72 h with VER155008 (V2, V10, V50) alone or in combination with doxorubicin (D0.2 μM). Cells in the lower left quadrant are considered viable, lower right early apoptosis, upper left undergoing necrosis, and upper right late apoptosis

Apoptosis in Dharma OSA cells as determined by annexin V/PI flow cytometry. VER155008 treatment produced primarily early phase apoptosis at 72 h. Combination treatments did not produce additive effects. Cells were incubated for 72 h with VER155008 (V2, V10, V50) alone or in combination with doxorubicin (D0.2 μM)

Apoptosis in D17 cells as determined by annexin V/PI flow cytometry. VER155008 treatment increased apoptosis. Combination treatment did not further increase apoptosis above that achieved by either drug alone. Cells were treated for 72 h with VER (V2, V10, V50) either alone or in combination with doxorubicin (D0.2 μM)
To assess whether VER treatment would reduce cellular repopulation, clonogenic survival assays were performed in JL31 cells. VER treatment reduced colony forming ability as a single agent (Fig. 5a). In combination treatment groups, colony formation was further reduced compared to VER and doxorubicin single-agent treatments (Fig. 5b).

Clonogenic survival of JL31 cells. a VER single-agent treatment decreased colony formation. b Combination treatments of VER and doxorubicin (0.2 μM) further reduced colony formation. [Cells were exposed to VER155008 for 72 h and doxorubicin for 48 h. “Live cells” indicates the number of viable cells replated after treatment in the dilution assay. N = 3]
HSP70 response to treatment
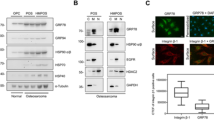
HSP70 expression levels were first assessed by western blot following VER treatment for 24 h, the time at which doxorubicin was added to combination treatment groups (Fig. 6). In all three canine OSA cell lines, increasing concentrations of VER resulted in dose-dependent increases in HSP70 expression. VER155008 is also known to bind GRP78 with an IC50 value of 2.6 μM (Massey et al. 2010). As such, protein expression levels of GRP78 were also examined via western blot. Similar to HSP70 but to a much larger degree, increased expression of GRP78 was observed following 24 h of VER treatment (Fig. 7). Furthermore, to investigate changes in cellular distribution of GRP78, which have been previously reported, including cell surface expression, GRP78 was assessed by immunofluorescence in D17 cells. A wider cytoplasmic and possibly cell surface distribution of GRP78 was noted in D17 cells following VER treatment (Fig. 8).

Western blot of HSP70 following treatment with VER. VER increased HSP70 expression in JL31 and D17 OSA cell lines. Cells were treated (V2, V10, V50) for 24 h. Values represented are the relative mean abundance of HSP70, compared to the untreated controls, ±SEM. *P < 0.05, statistical significance from both DMSO and untreated controls, as determined by an ANOVA from three replicates

Western blot of GRP78 following treatment with VER. VER increased GRP78 expression in canine OSA cell lines. Cells were treated for 24 h with VER (V2, V10, V50). *P < 0.05, statistical significance from both DMSO and the untreated control, as determined by an ANOVA from three replicates. Values represent the mean of the relative abundance of GRP78, compared to the untreated controls, ±SEM

Cellular localization of GRP78 in D17 cells after treatment with VER. GRP78 appeared increased at 24 and 48 h after VER treatment (V50) and localized throughout the cytoplasm, possibly the cell surface, and within the nucleus. GRP78 was predominantly localized within the nucleus in untreated cells. [Images were taken at ×60 magnification using oil immersion on a confocal microscope. N = 2]
Given the reported observation of increased HSP70 expression with the use of HSP90 inhibitors, we examined HSP90 expression following treatment with the HSP70 inhibitor VER. Significant alterations to HSP90 abundance following VER treatment were not appreciated, possibly apart from an increase in D17 cells (Fig. 9).

Western blot of HSP90 following VER treatment. Only D17 cells showed a significant alteration to HSP90 abundance following VER treatment. Values represent relative mean of HSP90 expression, ±SEM, after 24 h of treatment with VER155008 (V2, V10, V50). *P < 0.05, statistical significance from both DMSO and untreated controls, as determined by an ANOVA on three replicates
Unfolded protein response and Akt signaling
GRP78 is known as a marker for the unfolded protein response (UPR) (Samali et al. 2010). Thus, it is possible that the observed increased expression of GRP78 could be an indication of UPR, following VER treatment. In addition to GRP78, other markers of the UPR, including Herp and CHOP, were examined to determine the likelihood of UPR activation in canine OSA cells. JL31 and D17 cells both showed a marked upregulation of Herp and CHOP after VER treatment, suggesting that the UPR is occurring and may also be responsible for the increased expression of GRP78 (Fig. 10).

Western blot of Herp and CHOP following treatment with VER. VER increased expression of Herp and CHOP. Cells were treated for 24 h with VER (V2, V10, V50). N = 3
HSP70 expression is typically controlled through one of two major transcription factor families: heat shock factors and hypoxia-inducible factors (Akerfelt et al. 2010; Huang et al. 2009). Since oxygen levels were not altered during these experiments, the heat shock factor pathway was indirectly inspected via western blot. A major signaling pathway that activates heat shock factors is the phosphoinositol 3-kinase, Akt/protein kinase B signaling cascade (Carpenter et al. 2015; Chatterjee et al. 2013; Mustafi et al. 2010). After 24 h of treatment, VER decreased overall Akt and serine-473 phosphorylated Akt, suggesting that Akt pathway activation is not responsible for the observed increase in HSP70 after VER treatment (Fig. 11).

Western blot of Akt following treatment with VER. Total Akt and phosphorylated Akt (P-Akt) decreased following treatment with VER (V2, V10, V50). Cells were treated for 24 h with VER. N = 3
HSP70 function
Optimal HSP70 function requires coordination with co-chaperones and nucleotide exchange factors. One such nucleotide exchange factor is BAG-1 (Mayer and Bukau 2005). BAG-1 generates conformational changes to HSP70 family members, which are unfavorable to nucleotide binding and promote ADP release (Hohfeld and Jentsch 1997; Sondermann et al. 2001). Treatment with VER resulted in the degradation of BAG-1 in JL31 and D17 cell lines. As the mechanism of VER is inhibition of the ATP binding pocket, this result reveals that HSP70 function is also impacted in another way following VER treatment (Fig. 12).

Western blot of BAG-1 following HSP70 inhibition. Both human and canine cells display less BAG-1 after VER treatment. Cells were treated for 24 h with VER (V2, V10, V50). N = 3
Discussion
HSP70 is known to prevent apoptosis through a variety of molecular mechanisms and becomes activated in response to cellular stress. Thus, it was hypothesized that the inhibition of HSP70 would facilitate greater canine OSA cell death when combined with a cellular stressor such as doxorubicin. We found that OSA treatment with VER alone induced apoptosis and reduced viability and clonogenic survival. However, combination treatments with doxorubicin were unable to significantly increase apoptosis or decrease cellular viability beyond that achieved by chemotherapy alone, although improved results were observed with combination treatment in clonogenic survival assays. These findings may highlight that combination treatments are more efficient at preventing the clonogenic outgrowth/repopulation of OSA cells than in inducing apoptosis.
To investigate possible mechanisms for the lack of additive effects in cellular viability and apoptosis, target protein expression was assessed via western blot. As VER specifically interacts with the ATPase binding domain of HSP70, it is not expected to change HSP70 expression with treatment, which has been observed by others (Wen et al. 2014). However, in our canine OSA cells, HSP70 expression actually increased after 24 h of incubation with VER (the time at which doxorubicin would be added). This increased expression of HSP70 is potentially problematic, in that the protein is known to directly suppress the apoptotic cascade and protect cells from doxorubicin-mediated apoptosis in a variety of model systems (Gabai et al. 1997, 1995; Karlseder et al. 1996; Samali and Cotter 1996). Although we did not confirm a cytoprotective role for the increased HSP70 observed in canine OSA cell lines, induced overexpression of HSP70 in human OSA cell lines partially prevented apoptosis induced by the flavenoid baicalein (Ding et al. 2014).
Another potentially problematic and possibly compensatory response to VER treatment was increased GRP78 expression, which is also a target of this compound. Several associations between GRP78 expression and chemotherapy resistance have been identified. For instance, GRP78 hindered etoposide and cisplatin-induced apoptosis in glioblastoma cells (Lee et al. 2008). Several possible mechanisms for the chemoresistance induced by GRP78 have been suggested. These mechanisms include activation of pro-survival kinase pathways such as Akt and ERK or inhibition of caspase-7, bik, and bax (Fu et al. 2007; Misra et al. 2006; Ranganathan et al. 2006; Reddy et al. 2003). Based on this evidence, the upregulation of GRP78 after VER treatment, prior to the addition of doxorubicin, is also potentially problematic and could explain the lack of improvement observed with combination treatments.
Recognized as an indicator for the unfolded protein response, GRP78 is typically localized to the endoplasmic reticulum (Samali et al. 2010). However, various studies demonstrate that GRP78 is localized to the cell surface, nucleus, and cytoplasm under differing cellular environments (Ni et al. 2011). At the cell surface, GRP78 has been associated with various receptors, notably binding kringle 5, microplasminogen, and α2 M macroglobulin (Gonzalez-Gronow et al. 2007; Misra et al. 2002). Multiple studies have taken advantage of this cell surface location as a method of specifically targeting cancer cells, due to the lack of GRP78 on the surface of non-malignant cells (Arap et al. 2004; de Ridder et al. 2012; Liu et al. 2007; Ni et al. 2011). Ligands that bind GRP78 become internalized into the cell in a GRP78-dependent mechanism (Liu et al. 2007). A significantly lower abundance of these ligands is found in the liver, spleen, brain, and pancreas when compared to tumor tissue (Liu et al. 2007). The present study demonstrated that VER treatment induces ER stress in canine OSA cells. Following ER stress, significantly higher levels of GRP78 travel to the cell surface (Zhang et al. 2010). Although further work is required in these canine cell lines to determine if GRP78 is indeed expressed directly on the cell surface, our initial immunofluorescence results are promising. Following overexpression, GRP78 was not localized to a single location. Furthermore, some cells appeared to have GRP78 localized close to the cell surface. Thus, this overexpression of GRP78 could be capitalized upon to possibly provide cancer cell-specific targeting following VER treatment. As GRP78 moves to the cell surface following VER administration, OSA cells could be targeted with GRP78-bound peptide conjugates, such as Pep42 which can be linked to a cytotoxic payload like doxorubicin (Liu et al. 2007).
To function optimally in response to stress, HSP70 operates with several co-chaperones. The canonical model of protein folding posits that one of the initial stages in protein refolding is the delivery of the client to HSP70 by co-chaperones (Kampinga and Craig 2010). By binding and delivering substrates to HSP70, these chaperone proteins are inherently preventing their aggregation. Heat-denatured luciferase experiments confirm that the co-chaperone carboxyl terminus of Hsc70-interacting protein (CHIP) possesses chaperone-like activity independent of HSP70 interactions (Rosser et al. 2007). Thus, it is possible that inhibition of a single heat shock protein may be aided by the lack of functional co-chaperones. Accordingly, this study found a direct correlation between BAG-1 expression and apoptosis in D17 and JL31 canine OSA cell lines. BAG-1 is known as a nucleotide exchange factor for HSP70 (Mayer and Bukau 2005). Binding of the BAG domain to HSP70 increases nucleotide exchange rates up to 900-fold, allowing for client substrates to be released after proper protein folding is completed (Gassler et al. 2001). At the highest dose of VER, which was the most effective in all cell lines, the lowest abundance of BAG-1 was evident. This decreased BAG-1 should facilitate longer interactions of HSP70 with client proteins, ultimately slowing their release and promoting the aggregation of misfolded proteins.
Given HSP70’s role in cellular survival, this protein could be an important therapeutic target in many malignancies. However, redundancy of chaperones highlights one of the challenges that exist in HSP targeting, so simultaneous inhibition of multiple family members may be an important strategy moving forward. VER treatment was sufficient to decrease cellular viability, induce apoptosis, and reduce clonogenic survival of canine OSA cells in our study. Unfortunately, combination treatments with doxorubicin chemotherapy showed no meaningful increase in apoptosis. This result could be due to the upregulation of HSP70, GRP78, or other proteins prior to the administration of chemotherapy. Further determination of the pathways involved in the potentially compensatory increases in HSP70 and GRP78 expression could yield useful information for optimizing combination schedules.
References
Akerfelt M, Morimoto RI, Sistonen L (2010) Heat shock factors: integrators of cell stress, development and lifespan. Nat Rev Mol Cell Biol 11(8):545–555
Arap MA, Lahdenranta J, Mintz PJ, Hajitou A, Sarkis ÁS, Arap W, et al. (2004) Cell surface expression of the stress response chaperone GRP78 enables tumor targeting by circulating ligands. Cancer Cell 6(3):275–284
Beere HM, Wolf BB, Cain K, Mosser DD, Mahboubi A, Kuwana T, et al. (2000) Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the apaf-1 apoptosome. Nat Cell Biol 2(8):469–475
Berg J, Gebhardt MC, Rand WM (1997) Effect of timing of postoperative chemotherapy on survival of dogs with osteosarcoma. Cancer 79(7):1343–1350
Bergman PJ, MacEwen EG, Kurzman ID, Henry CJ, Hammer AS, Knapp DW, et al. (1996) Amputation and carboplatin for treatment of dogs with osteosarcoma: 48 cases (1991 to 1993). J Vet Intern Med 10(2):76–81
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1):248–254
Carpenter RL, Paw I, Dewhirst MW, Lo H-W (2015) Akt phosphorylates and activates HSF-1 independent of heat shock, leading to slug overexpression and epithelial–mesenchymal transition (EMT) of HER2-overexpressing breast cancer cells. Oncogene 34:546–557
Chatterjee M, Andrulis M, Stuhmer T, Muller E, Hofmann C, Steinbrunn T, et al. (2013) The PI3K/akt signaling pathway regulates the expression of Hsp70 which critically contributes to Hsp90-chaperone function and tumor cell survival in multiple myeloma. Hematologica 98(7):1132–1141
Ding L, He S, Sun X (2014) HSP70 desensitizes osteosarcoma cells to baicalein and protects cells from undergoing apoptosis. Apoptosis 19(8):1269–1280
Elmore S (2007) Apoptosis: a review of programmed cell death. Toxicol Pathol 35(4):495–516
Feder ME, Hofmann GE (1999) Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and ecological physiology. Ann Rev Physiol 61(1):243–282
Fu Y, Li J, Lee AS (2007) GRP78/BiP inhibits endoplasmic reticulum BIK and protects human breast cancer cells against estrogen starvation-induced apoptosis. Cancer Res 67(8):3734–3740
Gabai VL, Meriin AB, Mosser DD, Caron AW, Rits S, Shifrin VI, et al. (1997) Hsp70 prevents activation of stress kinases. A novel pathway of cellular thermotolerance. J Biol Chem 272(29):18033–18037
Gabai VL, Zamulaeva IV, Mosin AF, Makarova YM, Mosina VA, Budagova KR, et al. (1995) Resistance of ehrlich tumor cells to apoptosis can be due to accumulation of heat shock proteins. FEBS Lett 375(1):21–26
Gabai VL, Mabuchi K, Mosser DD, Sherman MY (2002) Hsp72 and stress kinase c-Jun N-terminal kinase regulate the bid-dependent pathway in tumor necrosis factor-induced apoptosis. Mol Cell Biol 22(10):3415–3424
Gassler CS, Wiederkehr T, Brehmer D, Bukau B, Mayer MP (2001) Bag-1M accelerates nucleotide release for human Hsc70 and Hsp70 and can act concentration-dependent as positive and negative cofactor. J Biol Chem 276(35):32538–32544
Gonzalez-Gronow M, Kaczowka SJ, Payne S, Wang F, Gawdi G, Pizzo SV (2007) Plasminogen structural domains exhibit different functions when associated with cell surface GRP78 or the voltage-dependent anion channel. J Biol Chem 282(45):32811–32820
Gotoh T, Terada K, Oyadomari S, Mori M (2004) Hsp70-DnaJ chaperone pair prevents nitric oxide-and CHOP-induced apoptosis by inhibiting translocation of bax to mitochondria. Cell Death Differ 11(4):390–402
Guo F, Rocha K, Bali P, Pranpat M, Fiskus W, Boyapalle S, et al. (2005) Abrogation of heat shock protein 70 induction as a strategy to increase antileukemia activity of heat shock protein 90 inhibitor 17-allylamino-demethoxy geldanamycin. Cancer Res 65(22):10536–10544
Gurbuxani S, Schmitt E, Cande C, Parcellier A, Hammann A, Daugas E, et al. (2003) Heat shock protein 70 binding inhibits the nuclear import of apoptosis-inducing factor. Oncogene 22(43):6669–6678
Hartl FU, Bracher A, Hayer-Hartl M (2011) Molecular chaperones in protein folding and proteostasis. Nature 475(7356):324–332
Hohfeld J, Jentsch S (1997) GrpE-like regulation of the hsc70 chaperone by the anti-apoptotic protein BAG-1. EMBO J 16(20):6209–6216
Huang WJ, Xia LM, Zhu F, Huang B, Zhou C, Zhu HF, et al. (2009) Transcriptional upregulation of HSP70-2 by HIF-1 in cancer cells in response to hypoxia. Int J Cancer 124(2):298–305
Kampinga HH, Craig EA (2010) The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat Rev Mol Cell Biol 11(8):579–592
Karlseder J, Wissing D, Holzer G, Orel L, Sliutz G, Auer H, et al. (1996) HSP70 overexpression mediates the escape of a doxorubicin-induced G2 cell cycle arrest. Biochem Biophys Res Commun 220(1):153–159
Khoury JF, Ben-Arush MW, Weintraub M, Waldman E, Futerman B, Vlodavsky E, et al. (2014) Alkaline phosphatase level change in patients with osteosarcoma: its role as a predictive factor of tumor necrosis and clinical outcome. Isr Med Assoc J 16(1):26–32
Kregel KC (2002) Heat shock proteins: modifying factors in physiological stress responses and acquired thermotolerance. J Appl Physiol (1985) 92(5):2177–2186
Lee HK, Xiang C, Cazacu S, Finniss S, Kazimirsky G, Lemke N, et al. (2008) GRP78 is overexpressed in glioblastomas and regulates glioma cell growth and apoptosis. Neuro-Oncology 10(3):236–243
Leu J, Pimkina J, Frank A, Murphy ME, George DL (2009) A small molecule inhibitor of inducible heat shock protein 70. Mol Cell 36(1):15–27
Li H, Liu L, Xing D, Chen WR (2010) Inhibition of the JNK/Bim pathway by Hsp70 prevents bax activation in UV-induced apoptosis. FEBS Lett 584(22):4672–4678
Liu Y, Steiniger SC, Kim Y, Kaufmann GF, Felding-Habermann B, Janda KD (2007) Mechanistic studies of a peptidic GRP78 ligand for cancer cell-specific drug delivery. Mol Pharm 4(3):435–447
London CA, Bear MD, McCleese J, Foley KP, Paalangara R, Inoue T, et al. (2011) Phase I evaluation of STA-1474, a prodrug of the novel HSP90 inhibitor ganetespib in dogs with spontaneous cancer. PLoS One 6(11):e27018
Massey AJ, Williamson DS, Browne H, Murray JB, Dokurno P, Shaw T, et al. (2010) A novel, small molecule inhibitor of Hsc70/Hsp70 potentiates Hsp90 inhibitor induced apoptosis in HCT116 colon carcinoma cells. Cancer Chemother Pharmacol 66(3):535–545
Mauldin GN, Matus RE, Withrow SJ, Patnaik AK (1988) Canine osteosarcoma treatment by amputation versus amputation and adjuvant chemotherapy using doxorubicin and cisplatin. J Vet Intern Med 2:177–180
Mayer M, Bukau B (2005) Hsp70 chaperones: cellular functions and molecular mechanism. Cell Mol Life Sci 62(6):670–684
Misra UK, Deedwania R, Pizzo SV (2006) Activation and cross-talk between akt, NF-kappaB, and unfolded protein response signaling in 1-LN prostate cancer cells consequent to ligation of cell surface-associated GRP78. J Biol Chem 281(19):13694–13707
Misra UK, Gonzalez-Gronow M, Gawdi G, Hart JP, Johnson CE, Pizzo SV (2002) The role of grp 78 in alpha 2-macroglobulin-induced signal transduction. Evidence from RNA interference that the low density lipoprotein receptor-related protein is associated with, but not necessary for, GRP 78-mediated signal transduction. J Biol Chem 277(44):42082–42087
Mustafi SB, Chakraborty PK, Raha S (2010) Modulation of akt and ERK1/2 pathways by resveratrol in chronic mylogenous leukemia (CML) cells results in the downregulation of Hsp70. PLoS One 5(1):e8719
Ni M, Zhang Y, Lee A (2011) Beyond the endoplasmic reticulum: atypical GRP78 in cell viability, signaling and therapeutic targeting. Biochem J 434:181–188
Ranganathan AC, Zhang L, Adam AP, Aguirre-Ghiso JA (2006) Functional coupling of p38-induced up-regulation of BiP and activation of RNA-dependent protein kinase-like endoplasmic reticulum kinase to drug resistance of dormant carcinoma cells. Cancer Res 66(3):1702–1711
Reddy RK, Mao C, Baumeister P, Austin RC, Kaufman RJ, Lee AS (2003) Endoplasmic reticulum chaperone protein GRP78 protects cells from apoptosis induced by topoisomerase inhibitors: role of ATP binding site in suppression of caspase-7 activation. J Biol Chem 278(23):20915–20924
de Ridder GG, Ray R, Pizzo SV (2012) A murine monoclonal antibody directed against the carboxyl-terminal domain of GRP78 suppresses melanoma growth in mice. Melanoma Res 22(3):225–235
Riggs JL, McAllister RM, Lennette EH (1974) Immunofluorescent studies of RD-114 virus replication in cell culture. J Gen Virol 25(1):21–29
Romanucci M, D’Amato G, Malatesta D, Bongiovanni L, Palmieri C, Ciccarelli A, et al. (2012) Heat shock protein expression in canine osteosarcoma. Cell Stress Chaperones 17(1):131–138
Rosser MF, Washburn E, Muchowski PJ, Patterson C, Cyr DM (2007) Chaperone functions of the E3 ubiquitin ligase CHIP. J Biol Chem 282(31):22267–22277
Ruchalski K, Mao H, Singh SK, Wang Y, Mosser DD, Li F, et al. (2003) HSP72 inhibits apoptosis-inducing factor release in ATP-depleted renal epithelial cells. Am J Physiol Cell Physiol 285(6):C1483–C1493
Samali A, Cotter TG (1996) Heat shock proteins increase resistance to apoptosis. Exp Cell Res 223(1):163–170
Samali A, Fitzgerald U, Deegan S, Gupta S (2010) Methods for monitoring endoplasmic reticulum stress and the unfolded protein response. Int J Cell Biol 2010:830307
Sondermann H, Scheufler C, Schneider C, Hohfeld J, Hartl FU, Moarefi I (2001) Structure of a bag/Hsc70 complex: convergent functional evolution of Hsp70 nucleotide exchange factors. Science (New York, NY) 291(5508):1553–1557
Straw RC, Withrow SJ, Richter SL, Powers BE, Klein MK, Postorino NC, et al. (1991) Amputation and cisplatin for treatment of canine osteosarcoma. J Vet Intern Med 5(4):205–210
Uozaki H, Ishida T, Kakiuchi C, Horiuchi H, Gotoh T, Iijima T, et al. (2000) Expression of heat shock proteins in osteosarcoma and its relationship to prognosis. Pathol Res Pract 196(10):665–673
Wen W, Liu W, Shao Y, Chen L (2014) VER-155008, a small molecule inhibitor of HSP70 with potent anti-cancer activity on lung cancer cell lines. Exp Biol Med (Maywood) 239:638–645
Zhang Y, Liu R, Ni M, Gill P, Lee AS (2010) Cell surface relocalization of the endoplasmic reticulum chaperone and unfolded protein response regulator GRP78/BiP. J Biol Chem 285(20):15065–15075
Zhuang H, Jiang W, Zhang X, Qiu F, Gan Z, Cheng W, et al. (2013) Suppression of HSP70 expression sensitizes NSCLC cell lines to TRAIL-induced apoptosis by upregulating DR4 and DR5 and downregulating c-FLIP-L expressions. J Mol Med (Berl) 91(2):219–235
Acknowledgments
This project was supported by the OVC Department of Biomedical Sciences, OVC Dean’s office, and a memorial donation through the University of Guelph Alumni Affairs and Development office. J. Asling was supported by a graduate stipend from the Art Rouse Cancer Biology fund at OVC.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Asling, J., Morrison, J. & Mutsaers, A.J. Targeting HSP70 and GRP78 in canine osteosarcoma cells in combination with doxorubicin chemotherapy. Cell Stress and Chaperones 21, 1065–1076 (2016). https://doi.org/10.1007/s12192-016-0730-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12192-016-0730-4