
Abstract
The microenvironment of oral cancer is highly dynamic and has been proved to affect tumor progression. Pericytes are blood vessels surrounding cells that have recently gained attention for their roles in vascular and cancer biology. The objective of the present study was to survey the scientific literature for conclusive evidence about whether pericytes are part of blood vessels in oral squamous cell carcinoma (OSCC) and their roles in the tumor microenvironment and clinical outcomes. A systematic electronic search was undertaken in Medline Ovid, PubMed, Web of Science, and Scopus. Eligibility criteria were: publications adopting in vivo models of OSCC that included pericyte detection and assessment by pericyte markers (e.g., α-smooth muscle actin, neuron-glial antigen 2 and platelet-derived growth factor receptor-β). The search yielded seven eligible studies (from 2008 to 2018). The markers most commonly used for pericyte detection were α-smooth muscle actin and neuron-glial antigen 2. The studies reviewed showed the presence of immature vessels exhibiting a reduction of pericyte coverage in OSCC and indicated that anti-cancer therapies could contribute to vessel normalization and pericyte regain. The pericyte population is significantly affected during OSCC development and cancer therapy. While these findings might suggest a role for pericytes in OSCC progression, the limited data available do not allow us to conclude whether they modify the tumor microenvironment and clinical outcome.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Pericytes were first identified in the nineteenth century by Rouget as a group of contractile cells surrounding the endothelial cells (ECs) of small blood vessels [1, 2]. They are a diverse group of cells of heterogeneous origin and functionally associated with vessel diameter, permeability and endothelial cell proliferation [3,4,5]. Other physiological roles of pericytes are leukocyte recruitment [6], the regulation of blood–brain barrier homeostasis [7] and kidney blood pressure [8].
Pericyte coverage (PC), i.e., the proportion of pericytes relative to endothelial cells, pericyte morphology and pericyte contact with ECs vary in different organs [9]. The most pericyte-covered area is observed in the blood–brain barrier with a ratio of 1:1 to 3:1 ECs for each pericyte [10]. Conversely, a PC of 1:100 has been observed in human skeletal striated muscles [11, 12]. Changes in PC have been associated with different pathological process such as diabetic retinopathy [13], Alzheimer’s disease [14], glomerulosclerosis [8], and several types of neoplasms [15,16,17,18,19,20].
PC exhibits significant changes in tumors of different origins. In glioblastomas and renal cell carcinomas, PC varies between 10 and 20%, while in mammary and colon carcinomas, the PC rate was 70% [15]. Tumors with fewer pericytes are generally characterized by dense neovascularization with active EC proliferation [15, 19].
In the neoplastic environment, pericytes interplay with a variety of extracellular matrix components and cell types [20]. Due to their multipotency, pericytes detached from the tumor microvasculature may operate as mesenchymal stem cell precursors, generating other stromal cells such as fibroblasts, which are known to contribute to tumor invasion and metastasis [21,22,23,24]. Because tumoral vessels are in a constant state of remodeling, pericytes in tumoral beds differ from habitual pericytes. Overall, they are more loosely attached to the ECs, and their cytoplasmic processes can extend into the tumor tissue [25]. Recently, it has been proposed that pericytes may provide a cellular target for pharmacological interventions based on angiogenesis inhibition [16, 26,27,28,29].
PC has been correlated with clinical parameters and its increase in melanoma, renal cell and ovarian carcinomas has been demonstrated to result in an unfavorable prognosis [16, 17]. In contrast, in colorectal cancer, the absence of PC was correlated with poor prognosis and metastasis [18]. In OSCC, PC distribution and its association with clinical outcomes have been less defined [30,31,32,33,34,35,36]. Therefore, the objective of this systematic review was to integrate the available data published in the literature in order to obtain conclusive evidence about whether pericytes are part of tumor vessels in OSCC and about their potential roles in neoplastic behavior.
Materials and Methods
Protocol and Registration
The reporting of the present systematic review conforms to the Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) Statement [37]. A protocol was drafted and registered with the International Prospective Register of Systematic Reviews (PROSPERO, No. CRD42019128572).
Eligibility Criteria
Eligibility criteria were: publications adopting in vivo models of OSCC that included pericyte assessment or pericyte identification by pericyte markers [e.g., α-smooth muscle actin (α-SMA), neuron-glial antigen 2 (NG2) and platelet-derived growth factor receptor-β (PDGFR-β)]. Letters to the editor, review articles, in vitro studies, studies in which some of the main pericyte markers were not used for pericyte detection and studies that did not include OSCC were excluded.
The PECO elements were as follows:
-
P (Patients): humans or animals;
-
E (Exposure): oral squamous cell carcinoma;
-
C (Comparison): no comparison;
-
O (Outcome): pericytes/pericyte markers.
Information Sources and Search Strategies
Computerized searches without publication date restrictions, but limited to publications in the English language, were undertaken in June 2019 and updated in May 2020 in the following electronic databases: Medline Ovid, PubMed, Web of Science and Scopus. The keywords used are reported in Appendix S1.
Hand searches were also conducted by cross-checking the reference lists of the included articles for the identification of publications that might have been missed during the searches in the electronic databases. Finally, a search using Google Scholar limiting the search to the first 100 hits was conducted. The retrieved references were exported to the EndNote software (Clarivate Analytics, Toronto, Canada). Duplicates were removed upon identification.
Study Selection
The titles/abstracts of all references retrieved were assessed independently by two review authors (I.B.V. and L.F.S.) and references whose title/abstract met the eligibility criteria were included. The full texts of the articles with titles/abstracts providing insufficient information for a decision were assessed and the references that met the eligibility criteria were included. Disagreements between review authors were resolved upon discussion with a third review author (T.A.S.).
Data Extraction
The following data (when available) were extracted on a standard form from each study included: first author’s last name, publication year, country where the study was conducted, study design and number of case(s) in the sample, methodology used, antibody assessed, and results with respect to pericyte identification.
Quality Assessment
For animal studies, the included articles were critically appraised using the SYRCLE’s risk of bias tool [38], and for studies with humans, they were appraised using the University of Adelaide critical appraisal tool for cross-sectional [39], as shown in Appendix S2.
Results
Search Results
A total of 1002 records were retrieved across the electronic databases. Thirty-eight references were selected for full text evaluation and seven articles were included [30,31,32,33,34,35,36]. Figure 1 illustrates the flowchart of this systematic review.

Flow diagram of the literature search and paper selection
A list of the 31 references excluded after full text assessment is provided in Appendix S3. The major reason for study exclusion was the use of pericyte markers for purposes other than pericyte identification. Another critical reason for exclusion referred to articles unrelated to OSCC. Several studies involved head and neck carcinomas with focus on laryngeal and pharyngeal anatomical sites. Moreover, some articles concerned mouse xenografts models in which tumors were generated from cell lines other than oral squamous cells.
Characteristics of the Included Studies
The seven included articles were published between 2008 and 2018. The main method of pericyte detection was immunofluorescence [31,32,33,34,35]. The most commonly used markers for pericyte detection were α-SMA [30,31,32, 35] and NG2 [34, 36]. Table 1 summarizes the main information about the included articles.
Quality Assessment of the Included Studies
Appendix S4 displays the results of the quality assessment of human studies using the the University of Adelaide tool and Appendix S5 presents the quality assessment of animal studies using the SYRCLE’s risk of bias tool.
Results of Individual Studies
In some of the included studies, the presence of PC in capillaries was consistent with vessel normalization or considered to be vessel maturation [32, 34, 36].
Regarding PC, Liu et al. [30] reported that the percentage of pericytes surrounding ECs in OSCC tumor vessels ranged from 42 to 78%, with a mean of 57%. Individuals with lymph node metastasis showed a loss of PC (p = 0.030) as well as a reduced count of tumor cell-lined vessels (p = 0.012). Additionally, there was an inverse correlation between PC and tumor-associated macrophages (TAM), with an increase in TAM count while PC was reduced (r = 0.768, p < 0.001). Likewise, Li et al. [32] reported a decrease in pericyte number by observing a reduction of the rate of α-SMA-positive vessels in angiopoietin-2 (Ang-2)-positive cases of OSCC (vascular maturation index—VMI: 0.26 ± 0.13) compared to Ang-2-negative cases (VMI: 0.33 ± 0.13) (p = 0.014). In contrast, the expression of vascular endothelial growth factor (VEGF) did not affect VMI (p = 0.073).
In an immunohistochemical characterization of tumor vessels, Margaritescu et al. [31] stated that, regardless of the histological degree of OSCC, there was a low proportion of mature vessels (low PC) in tumor stroma (p < 0.001), with more numerous mature vessels as the invasive component of OSCC (p < 0.05).
In a study of angiogenesis, Zhou et al. [33] examined semaphorin 4D (SEMA4D), a pro-angiogenic molecule whose function is analogous to that of VEGF when acting on the Plexin-B1 receptor. Tumors xenografts with silenced SEMA4D showed reduction of tumor vessels and pericytes (p < 0.05). Conversely, when the Plexin-B1 receptor was silenced, little effect on endothelial cell number and a slight inhibition of pericytes were observed in tumors, indicating that SEMA4D can possibly act through different pathways (p < 0.01). The cited study attempted to show that SEMA4D production by tumor cells induces the expression of PDGF-β and angiopoietin-like protein 4 (ANGPTL4) from ECs in a plexin-B1/Rho-dependent manner, thereby influencing proliferation and differentiation of pericytes and vascular permeability. In contrast, VEGF lacks these effects. However, a direct influence of SEMA4D on tumor features such as growth and invasiveness was not evaluated.
Ludwig et al. [35], assessing the effect of tumor-derived exosomes (TEX) on angiogenesis, demonstrated that TEX were angiogenesis inducers, promoting the formation of defined vascular structures in vivo. TEX treatment also resulted in an increased PC in blood vessels within the tumors. α-SMA expression was increased and was significantly higher after the injection of PCI-13 and SCCVII-derived exosomes (p < 0.005). In the untreated group, pericytes were observed only in blood vessels in the border zone between tumor tissue and healthy tongue tissue, but not in the tumor center. This study did not attempt to integrate TEX-driven angiogenesis with tumoral behavior.
Evaluating the effects of bevacizumab on cetuximab uptake during OSCC therapy by means of tumor treatment response analyses, Chung et al. [34] observed higher drug perfusion after neoadjuvant treatment (bevacizumab followed by cetuximab). This neoadjuvant therapy acted by promoting vessel normalization and higher PC compared to the administration of cetuximab only (p = 0.009). Similarly, Prince et al. [36], investigating anti-VEGFR approaches, observed an increase in PC (NG2:CD31 ratio) after anti-VEGFR2 therapy (p < 0.01) and after all tested anti-VEGFR3 doses compared to control (p < 0.02).
Regarding anti-VEGFR2 and anti-VEGFR3 optimal timing, data on the NG2:CD31 ratio demonstrated a vascular maturation effect over time. Tumors studied seven days after anti-VEGFR2 and anti-VEGFR3 administration showed a significant increase in the NG2:CD31 ratio than tumors of animals sacrificed at other experimental time points and controls (p < 0.02 and p < 0.01 respectively) [36].
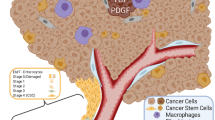
The reviewed studies clarified the fact that the pericyte population changes in OSCC after cancer progression. A summary of these data is illustrated in Fig. 2. A schematic representation demonstrates pericytes as constituents of the healthy oral mucosa and the OSCC microenvironment, together with several different cell types with which they interact, and points to the major changes that occur in microvascular density and PC after OSCC.

Pericytes as pivotal components of tumor stroma. Pericytes interplay with various extracellular matrix constituents and different cell types (i.e. fibroblasts, cancer cells, mural cells, endothelial cells and leukocytes) in the normal oral mucosa and in the OSCC microenvironment. In neoplasms, pericytes participate in endothelial cell normalization and maturation, vascular permeability and influx of inflammatory cells. In OSCC, the pericyte population is significantly reduced during cancer progression compared to healthy oral mucosa. Despite the increased microvascular density, PC reduction has been observed, with the occurrence of tumoral vessels that are often immature and defective
Discussion
Pericytes Definition and Heterogeneity
Pericytes are elemental constituents of the crosstalk among cells in the perivascular space [40]. An essential function of EC-pericyte is vascular maturation by the induction of the production and deposition of matrix components such as fibronectin, collagen, and laminin [41,42,43]. Furthermore, pericyte functions include controlling immune cells migration [6, 44], blood flow regulation [45] and tissue regeneration [5], since these cells may behave as stem cells [46,47,48,49,50,51]. However, the precise contribution of pericytes to these effects is still dubious due to the difficulty in accurately identifying these cells [51, 52].
Reinforcing the heterogeneity of the pericyte population, Birbrair et al. [53] categorized these cells into two types according to their distribution, function, expression markers and differentiation potential. Type-1 pericytes mostly contribute to adipose tissue and fibrosis formation, while Type-2 pericytes have the potential to differentiate into the neural lineage, skeletal muscle and blood vessels, being the only pericyte type recruited in tumor vessel formation [53, 54].
An accurate combinations of criteria should be employed for pericyte identification, including morphology, localization, relationship with ECs and expression of particular genes [2]. These features are not provided by most studies, hampering the proper identification of these cells.
Immunohistochemical markers commonly used to identify pericytes include α-SMA, NG2, PDGFR-β and desmin [2]. Some studies have assumed that an α-SMA-positive cell around a blood vessel is a pericyte, and the term pericyte is broadly used in the literature to designate any peri-endothelial mesenchymal cell [2, 30,31,32, 35].
Pericytes in OSCC Tumoral Angiogenesis
The onset of solid tumor growth is angiogenesis dependent [55]. In malignancies, there is an increase in microvascular density despite the reduced maturation index and the presence of vessels with a defective wall structure. Hence, in tumor microvasculature, pericytes are poorly associated with ECs [2, 30, 31]. Liu et al. [30] demonstrated that the percentage of tumor vessels covered by pericytes ranged from 42 to 78%, with a mean of 57%, while some blood vessels revealed a loss of PC in OSCC.
Differently from intermediate and immature vessels that presented no pericytes (α-SMA-), mature blood vessels in OCSS were predominantly covered with pericytes (CD105 + and α-SMA +) and were significantly more numerous at the invasive front of the tumor [32].
The status of pericytes throughout malignant transformation has been addressed in a study of prostatic intraepithelial neoplasia, suggesting that pericytes seem to retain their regular shape and functionality in potentially malignant disorders. In the normal prostate and in prostatic intraepithelial neoplasia, the major precursor of prostatic carcinoma [56], pericytes still show a close association with blood vessels [57]. In contrast, in poorly differentiated prostatic tumors, the shape of pericytes is unusual and their size is irregular, and these cells are loosely associated with blood vessels and cytoplasmic processes spreading towards nearby blood vessels [57]. However, these features have not yet been determined in OSCC.
These changes in PC during carcinogenesis may be related to a shift in protein expression. For example, in the presence of VEGF, Ang-2 induces neovascularization and reduces vascular integrity by interfering with the perivascular-endothelial cell interaction and providing improved access to angiogenic factors. Ang-2 overexpression in OSCC led to a reduction of mural cell count (including pericytes) and of VMI. The Ang-2 pathway is likely to be involved in such changes [32]. Zhou et al. [33] revealed another pathway involved in pericyte recruitment during angiogenesis. The authors showed that the plexin ligand semaphorin 4D (SEMA4D) acting on the plexin-B1 receptor in ECs is a pro-angiogenic factor for malignancies. Tumor-derived SEMA4D-Plexin-B1 actively induces the production of PDGF-β and ANGPTL-4. This process drives vascular permeability and can stimulate pericyte differentiation, proliferation and recruitment from stem cells. SEMA4D may be produced by tumor cells, promoting blood vessel growth with a stimulatory effect similar to that of VEGF [58].
Interestingly, the tumoral angiogenesis axis can also undergo interference from TEX, small vesicles produced by tumor cells which accumulate into the tumor microenvironment and are responsible for several cellular interactions [59]. Specifically, in OSCC, TEX produced and released by the tumor are capable of reprogramming ECs and pericytes, inducing their proliferation and migration [35].
Pericytes in Cancer Development and Metastasis
Lymph node metastasis is associated with a reduced percentage of microvessels in which ECs are surrounded by pericytes in OSCC, suggesting that the lack of pericytes results in disruption of the vascular structure and contributes to tumor metastasis [30].
Evidence that pericyte reduction can participate in metastatic events has been obtained from an experimental model of pancreatic cancer in pericyte-deficient Pdgfbret/ret mice. Pericyte dysfunction in those animals resulted in tumor β cell metastases to distant organs and local lymph nodes, demonstrating a role for pericytes in limiting tumor cell metastasis [60].
In addition, overexpression of PDGF-β by cancer cells can stimulate pericytes to acquire mesenchymal progenitor characteristics and turn into fibroblasts over a process named pericyte-fibroblast transition (PFT) [21, 61]. Cancer-associated fibroblasts (CAFs) are the main components of the stroma that surrounds cancer cells. They not only provide mechanical support for tumoral cells [62], but also secrete growth factors that directly affect the motility of cancer cells and control proliferation, survival and immunogenicity [63]. Indirectly, CAFS may also interfere with other stromal cells (e.g., immune cells and perivascular cells) and are a source of extracellular matrix-degrading proteases [62, 63]. These phenotypic changes caused by PFT might greatly contribute to tumor invasion and metastasis [21]. Moreover, Hosaka et al. [21] have clinically demonstrated that overexpression of PDGF-β was correlated with low survival in patients with different types of cancers, such as colorectal, glioblastoma and lung cancer, probably due to the discussed PFT events, although these mechanisms have not yet been described in OSCC.
Pericytes, the basement membrane and the endothelium itself constitute physical barriers against the flow of inflammatory cells to tumor tissue. Voisin et al. [64] suggested that inflammatory cells migrate through the vessel wall at permissive sites without PC. This finding was confirmed in a study by Liu et al. [30] in which the TAM count was increased while pericytes were reduced.
Remarkably, pericytes have shown the capacity to be converted into tumor cells. In oral cancer, pericytes with positive staining for the GT198 + oncoprotein were able to detach from blood vessels, proliferate into tumor nodules and originate GT198 + tumor cells, which migrated into lymph nodes. This up-regulation of GT198 was associated with increased lymph node metastasis [65].
Furthermore, in lung cancer, Murgai et al. [50] showed that pericytes activated premetastatic niche formation and metastasis. The primary tumor released factors that induced the phenotypic switching of pericytes and their detachment from blood vessels, contributing to the increase of extracellular matrix content and the formation of a niche prone to colonization by cancer cells.
Pericytes as Therapeutic Targets
Malignant cells need to access the blood circulation system in order to obtain oxygen and nutrients to survive and proliferate [66]. In cancer therapy, techniques seek to develop antiangiogenic agents that disrupt the formation of new blood vessels and destroy the existing tumoral vessels [67]. In this context, since pericytes substantially interact with ECs and other perivascular cells, they can also be considered as therapeutic targets in anticancer interventions [20].
The assessment of the causes of resistance to anti-VEGF therapy emphasizes the key role of pericytes as endothelium protectors [68]. The reduction of VEGF-A in pancreatic cancer models led to selective elimination of tumor blood vessels without PC. In this respect, tumoral vessels with disrupted PC or completely lacking pericytes are more responsive to drugs targeting ECs [34, 36, 68]. Double targeting for both ECs and pericytes has been suggested to have a broader anti-tumor effect by pruning tumor vessels rather than targeting ECs alone [69].
Prince et al. [36] explored the simultaneous approach to angiogenic normalization and stabilization by administering anti-VEGFR2 and anti-VEGFR3 drugs to increase the action of cetuximab. When assessing pericyte density after dual adjuvant therapy, PC increased compared to the treatment with cetuximab alone and the dual adjuvant anti-angiogenic therapy increased tumor chemotherapy uptake by restoring fluid dynamics [36]. Promoting pericyte regain can lead to this vascular normalization and restrict vessel sprouting, which could benefit chemotherapy delivery into the tumor [70].
Similarly, advances in vessel stabilization for drug delivery improvement in OSCC were also reported by Chung et al. [34] These authors evaluated the potential of pretreatment with bevacizumab for increasing tumor uptake of cetuximab. Neoadjuvant therapy provides adequate time for vascular normalization by increasing PC and has demonstrated better drug intake. Further therapy-based studies conducted on head and neck squamous cell cancer models have also shown that effective anticancer approaches produce vessel normalization and increased PC [71, 72].
On the other hand, since PDGF-β induces pericyte recruitment and provides stability for new vessels, PDGF-β depletion in models of melanoma and lung carcinoma did not improve tumor sensitivity when associated with VEGF-A. These observations suggest that additional therapeutic modalities targeting pericytes may not increase the antitumor effect already induced by anti-VEGF drugs in cancer models. Accordingly, the reduction of tumor vascularity may be secondary to pericyte regression [73].
In this respect, so far the prerogative of using pericytes as therapeutic targets has not been well determined [70]. In OSCC, however, significant results have been demonstrated, indicating that increased PC in tumor vessels and vascular normalization are favorable events for neoadjuvant therapies [34, 36].
The present systematic review has some limitations that should be acknowledged. There was a lack of homogeneous data for meta-analysis, since the studies involved different methodologies for pericyte identification. The second flaw noted during data compilation was the limited number of articles that met our eligibility criteria. Further research more focused at pericyte detection in OSCC should be conducted in order to examine the participation of these cells in oral carcinogenesis and therefore their potential therapeutic use.
Conclusions
In summary, the pericyte population is significantly affected during OSCC development and cancer therapy. PC is decreased in OSCC immature intratumoral vessels and data suggest that anticancer therapies restore blood vessels with pericyte regain. Although these findings might suggest a role for pericytes in cancer progression, the limited data available do not allow us to conclude whether changes in PC modify the tumor microenvironment and clinical outcome in OSCC.
References
Rouget C. Memoire sur le developpement, la structures et les proprietes des capillaires sanguins et lymphatiques. Archs Physiol Norm Pathol. 1873;5:603–33.
Armulik A, Genové G, Betsholtz C. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell. 2011;21:193–21515.
Prazeres PH, Sena IFG, Borges IDT, et al. Pericytes are heterogeneous in their origin within the same tissue. Dev Biol. 2017;427:6–11.
Diaz-Flores L, Gutierrez R, Varela H, Rancel N, Valladares F. Microvascular pericytes: A review of their morphological and functional characteristics. Histol Histopathol. 1991;6:269–86.
Crisan M, Yap S, Casteilla L, et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008;3:301–13.
Ayres-Sander CE, Lauridsen H, Maier CL, Sava P, Pober JS, Gonzalez AL. Transendothelial migration enables subsequent transmigration of neutrophils through underlying pericytes. PLoS ONE. 2013;8:1–12.
Kitahara H, Kajikawa S, Ishii Y, et al. The novel pathogenesis of retinopathy mediated by multiple RTK signals is uncovered in newly developed mouse model. EBioMedicine. 2018;31:190–201.
Shaw I, Rider S, Mullins J, Hughes J, Péault B. Pericytes in the renal vasculature: roles in health and disease. Nat Rev Nephrol. 2018;14:521–34.
Von Tell D, Armulik A, Betsholtz C. Pericytes and vascular stability. Exp Cell Res. 2006;312:623–9.
Mathiisen TM, Lehre KP, Danbolt NC, Ottersen OP. The perivascular astroglial sheath provides a complete covering of the brain microvessels: an electron microscopic 3D reconstruction. Glia. 2010;58:1094–103.
Shepro D, Morel NM. Pericyte physiology. FASEB J. 1993;7:1031–8.
Díaz-Flores L, Gutiérrez R, Madrid JF, et al. Pericytes. Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histol Histopathol. 2009;24:909–69.
Park DY, Lee J, Kim J, et al. Plastic roles of pericytes in the blood–retinal barrier. Nat Commun. 2017;8:1–16.
Sagare AP, Bell RD, Zhao Z, Ma Q, Winkler EA, Ramanathan A, Zlokovic BV. Pericyte loss influences Alzheimer-like neurodegeneration in mice. Nat Commun. 2013;4:1–14.
Eberhard A, Kahlert S, Goede V, Hemmerlein B, Plate KH, Augustin HG. Heterogeneity of angiogenesis and blood vessel maturation in human tumors: implications for antiangiogenic tumor therapies. Cancer Res. 2000;60:1388–93.
Gee MS, Procopio WN, Makonnen S, Feldman MD, Yeilding NM, Lee WMF. Tumor vessel development and maturation impose limits on the effectiveness of anti-vascular therapy. Am J Pathol. 2003;162:183–93.
Sinha D, Chong L, George J, et al. Pericytes promote malignant ovarian cancer progression in mice and predict poor prognosis in serous ovarian cancer patients. Clin Cancer Res. 2006;22:1813–24.
Yonenaga Y, Mori A, Onodera H, et al. Absence of smooth muscle actin-positive pericyte coverage of tumor vessels correlates with hematogenous metastasis and prognosis of colorectal cancer patients. Oncology. 2005;69:159–66.
Baluk P, Morikawa S, Haskell A, Mancuso M, McDonald DM. Abnormalities of basement membrane on blood vessels and endothelial sprouts in tumors. Am J Pathol. 2002;160:985–1000.
Raza A, Franklin MJ, Dudek AZ. Pericytes and vessel maturation during tumor angiogenesis and metastasis. Am J Hematol. 2010;85:593–8.
Hosaka K, Yang Y, Seki T, et al. Pericyte-fibroblast transition promotes tumor growth and metastasis. Proc Natl Acad Sci USA. 2016;113:5618–27.
Supakul S, Yao K, Ochi H, et al. Pericytes as a source of osteogenic cells in bone fracture healing. Int J Mol Sci. 2019;20:1–14.
Dellavalle A, Sampaolesi M, Tonlorenzi R, et al. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat Cell Biol. 2007;9:255–67.
Farrington-Rock C, Crofts NJ, Doherty MJ, Ashton BA, Griffin-Jones C, Canfield AE. Chondrogenic and adipogenic potential of microvascular pericytes. Circulation. 2004;110:2226–32.
Bergers G, Song S. The role of pericytes in blood-vessel formation and maintenance. Neuro-Oncology. 2005;7:452–64.
Lu C, Shahzad MM, Moreno-Smith M, et al. Targeting pericytes with a PDGF-B aptamer in human ovarian carcinoma models. Cancer Biol Ther. 2010;3:176–82.
Zhou W, Chen C, Shi Y, et al. Targeting glioma stem cell-derived pericytes disrupts the blood-tumor barrier and improves chemotherapeutic efficacy. Cell Stem Cell. 2017;5:591–603.
Guerra DAP, Paiva AE, Sena IFG, Azevedo PO, Silva WN, Mintz A, Birbrair A. Targeting glioblastoma-derived pericytes improves chemotherapeutic outcome. Angiogenesis. 2018;4:667–75.
Thijssen VL, Paulis YW, Nowak-Sliwinska P, et al. Targeting PDGF-mediated recruitment of pericytes blocks vascular mimicry and tumor growth. J Pathol. 2018;4:447–58.
Liu SY, Chang LC, Pan LF, Hung YJ, Lee CH, Shieh YS. Clinicopathologic significance of tumor cell-lined vessel and microenvironment in oral squamous cell carcinoma. Oral Oncol. 2008;44:277–85.
Margaritescu C, Simionescu C, Pirici D, Mogoanta L, Ciurea R, Stepan A. Immunohistochemical characterization of tumoral vessels in oral squamous cell carcinoma. Rom J Morphol Embryol. 2008;49:447–58.
Li C, Sun CJ, Fan JC, et al. Angiopoietin-2 expression is correlated with angiogenesis and overall survival in oral squamous cell carcinoma. Med Oncol. 2013;30:1–10.
Zhou H, Yang Y-H, Basile J. The Semaphorin 4D- Plexin-B1- RhoA signaling axis recruits pericytes and regulates vascular permeability through endothelial production of PDGF-B and ANGPTL4. Angiogenesis. 2014;17:261–74.
Chung TK, Warram J, Day KE, Hartman Y, Rosenthal EL. Time-dependent pretreatment with bevacuzimab increases tumor specific uptake of cetuximab in preclinical oral cavity cancer studies. Cancer Biol Ther. 2015;16:790–8.
Ludwig N, Yerneni SS, Razzo BM, Whiteside TL. Exosomes from HNSCC promote angiogenesis through reprogramming of endothelial cells. Mol Cancer Res. 2018;16:1798–808.
Prince AC, Patel NG, Moore LS, et al. Adjuvant anti-angiogenic therapy enhances chemotherapeutic uptake in a murine model of head and neck cancer. J Drug Target. 2018;27:193–200.
Moher D, Liberati A, Tetzlaff J, Altman DG. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. Ann Intern Med. 2009;151:264–9.
Hooijmans CR, Rovers MM, de Vries RBM, Leenaars M, Ritskes-Hoitinga M, Langendam MW. SYRCLE’s risk of bias tool for animal studies. BMC Med Res Methodol. 2014;14:1–9.
Moola S, Munn Z, Tufanaru C, et al (2017) Checklist for analytical cross sectional studies. JBI Reviewer's Manual
Vanlandewijck M, He L, Mäe MA, et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature. 2018;554:475–80.
Lindahl P, Johansson BR, Levéen P, Betsholtz C. Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science. 1997;277:242–5.
Stratman AN, Malotte KM, Mahan RD, et al. Pericyte recruitment during vasculogenic tube assembly stimulates endothelial basement membrane matrix formation. Blood. 2009;114:5091–101.
Velez DO, Tsui B, Goshia T, et al. 3D collagen architecture induces a conserved migratory and transcriptional response linked to vasculogenic mimicry. Nat Commun. 2017;8:1651.
Rudziak P, Ellis CG, Kowalewska PM. Role and molecular mechanisms of pericytes in regulation of leukocyte diapedesis in inflamed tissues. Mediators Inflamm. 2019. https://doi.org/10.1155/2019/4123605.
Hall CN, Reynell C, Gesslein B, et al. Capillary pericytes regulate cerebral blood flow in health and disease. Nature. 2014;7494:55–60.
Birbrair A, Borges IDT, Gilson Sena IF, et al. How plastic are pericytes? Stem Cells Dev. 2017;26:1013–9.
Asada N, Kunisaki Y, Pierce H, et al. Differential cytokine contributions of perivascular haematopoietic stem cell niches. Nat Cell Biol. 2017;19:214–23.
Azevedo PO, Sena IFG, Andreotti JP, et al. Pericytes modulate myelination in the central nervous system. J Cell Physiol. 2018;8:5523–9.
Borges I, Sena I, Azevedo P, et al. Lung as a niche for hematopoietic progenitors. Stem Cell Rev. 2017;13:567–74.
Murgai M, Ju W, Eason M, et al. KLF4-dependent perivascular cell plasticity mediates pre-metastatic niche formation and metastasis. Nat Med. 2018;23:1176–90.
Hill RA, Tong L, Yuan P, Murikinati S, Gupta S, Grutzendler J. Regional blood flow in the normal and ischemic brain is controlled by arteriolar smooth muscle cell contractility and not by capillary pericytes. Neuron. 2015;87:95–110.
Guimaraes-Camboa N, Cattaneo P, Sun Y, et al. Pericytes of multiple organs do not behave as mesenchymal stem cells in vivo. Cell Stem Cell. 2017;20:345–59.
Birbrair A, Zhang T, Wang ZM, Messi ML, Enikolopov GN, Mintz A, Delbono O. Role of pericytes in skeletal muscle regeneration and fat accumulation. Stem Cells Dev. 2013;16:2298–314.
Birbrair A, Zhang T, Wang ZM, Messi ML, Mintz A, Delbono O. Pericytes at the intersection between tissue regeneration and pathology. Clin Sci. 2015;128:81–93.
Shivamallappa SM, Venkatraman NT, Shreedhar B, Mohanty L, Shenoy S. Role of angiogenesis in oral squamous cell carcinoma development and metastasis: an immunohistochemical study. Int J Oral Sci. 2011;3:216–24.
Brawer MK. Prostatic intraepithelial neoplasia: an overview. Rev Urol. 2005;7(Suppl 3):S11–S18.
Ozawa MG, Yao VJ, Chanthery YH, et al. Angiogenesis with pericyte abnormalities in a transgenic model of prostate carcinoma. Cancer. 2005;10:2104–15.
Basile JR, Castilho RM, Williams VP, Gutkind JS. Semaphorin 4D provides a link between axon guidance processes and tumor-induced angiogenesis. Proc Natl Acad Sci USA. 2006;103:9017–22.
Saleem SN, Abdel-Mageed AB. Tumor-derived exosomes in oncogenic reprogramming and cancer progression. Cell Mol Life Sci. 2015;72:1–10.
Xian X, Håkansson J, Ståhlberg A, et al. Pericytes limit tumor cell metastasis. J Clin Invest. 2006;116:642–51.
Ning X, Zhang H, Wang C, Song X. Exosomes released by gastric cancer cells induce transition of pericytes into cancer-associated fibroblasts. Med Sci Monit. 2018;24:2350–9.
Ishii G, Ochiai A, Neri S. Phenotypic and functional heterogeneity of cancer-associated fibroblast within the tumor microenvironment. Adv Drug Deliv Rev. 2016;1:186–96.
Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer. 2006;6:392–401.
Voisin MB, Pröbstl D, Nourshargh S. Venular basement membranes ubiquitously express matrix protein low-expression regions. Am J Pathol. 2010;176:482–95.
Zhang L, Wang Y, Rashid MH. Malignant pericytes expressing GT198 give rise to tumor cells through angiogenesis. Oncotarget. 2017;8:51591–607.
Lugano R, Ramachandran M, Dimberg A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell Mol Life Sci. 2019;77:1745–70.
Maj E, Papiernik D, Wietrzyk J. Antiangiogenic cancer treatment: The great discovery and greater complexity. Int J Oncol. 2016;5:1773–844.
Loges S, Schmidt T, Carmeliet P. Mechanisms of resistance to anti-angiogenic therapy and development of third-generation anti-angiogenic drug candidates. Genes Cancer. 2010;1:12–25.
Bergers G, Song S, Meyer-Morse N, Bergsland E, Hanahan D. Benefits of targeting both pericytes and endothelial cells in the tumor vasculature with kinase inhibitors. J Clin Investig. 2003;111:1287–95.
Meng MB, Zaorsky NG, Deng L, et al. Pericytes: a double-edged sword in cancer therapy. Future Oncol. 2015;11:169–79.
Koonce NA, Griffin RJ, Dings RPM. Galectin-1 inhibitor otx008 induces tumor vessel normalization and tumor growth inhibition in human head and neck squamous cell carcinoma models. Int J Mol Sci. 2017;18:1–9.
Mondini M, Nizard M, Tran T, et al. Synergy of radiotherapy and a cancer vaccine for the treatment of HPV-associated head and neck cancer. Mol Cancer Ther. 2015;6:1336–455.
Nisancioglu MH, Betsholtz C, Genové G. The absence of pericytes does not increase the sensitivity of tumor vasculature to vascular endothelial growth factor-A blockade. Cancer Res. 2010;70:5109–15.
Acknowledgements
Research supported by the Brazilian National Council for Scientific and Technological Development (CNPq). T.A.S. and L.G.A. are research fellows of CNPq. I.B.V. and L.F.S. are the recipients of a fellowship granted by the Coordination of Improvement of Higher Education Personnel—Brazil (CAPES, Finance code 001).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Valle, I.B., Schuch, L.F., da Silva, J.M. et al. Pericyte in Oral Squamous Cell Carcinoma: A Systematic Review. Head and Neck Pathol 14, 1080–1091 (2020). https://doi.org/10.1007/s12105-020-01188-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12105-020-01188-2