
Abstract
Advanced backcross QTL (AB-QTL) analysis was carried out in two Oryza nivara-derived BC2F2 populations. For nine traits, we identified 28 QTL in population 1, and 26 QTL in population 2. The two most significant yield-enhancing QTL, yldp9.1 and yldp2.1, showed an additive effect of 16 and 7 g per plant in population 1, while yld2.1 and yld11.1 showed an additive effect of 11 and 10 g per plant in population 2. At least one O. nivara-derived QTL with a phenotypic variance of more than 15% was detected for seven traits in population 1 and three traits in population 2. The O. nivara-derived QTL ph1.1, nt12.1, nsp1.1, nfg1.1, bm11.1, yld2.1, and yld11.1 were conserved at the same chromosomal locations in both populations. Two major QTL clusters were detected at the marker intervals RM488–RM431 and RM6–RM535 on chromosomes 1 and 2, respectively. The co-location of O. nivara-derived yield QTL with yield meta-QTL on chromosomes 1, 2, and 9 indicates their accuracy and consistency. The major-effect QTL reported in this study are useful for marker-assisted breeding and are also suitable for further fine mapping and candidate gene identification.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice is one of the most important and widely adapted cereal crops in the world. Its wild relatives are naturally grown in diverse environmental conditions and they harbour a wide range of allelic variations (Tanksley and Nelson 1996; Yano 2001; Quan et al. 2012). Further, the de novo genetic variations that arise in the progenies of crosses between wild species and elite varieties add to the available genetic diversity (Dong et al. 2006). In the past, wild species were largely used to improve disease and pest resistance in rice, but in the last 15 years, they have been increasingly used as a source of yield-enhancing QTL (Tanksley and McCouch 1997; Gur and Zamir 2004).
Yield is a complex trait and it needs to be continuously improved. The identification of novel yield-enhancing QTL from wild species using the advanced backcross QTL method is a popular strategy for mapping and simultaneously making available introgression lines that can be evaluated for the presence of QTL and for selecting high-yielding lines (Tanksley and Nelson 1996). Using this strategy, numerous yield-enhancing QTL have been mapped from wild species (Swamy and Sarla 2008).
Wild progenitor species Oryza rufipogon and O. nivara are easily crossable with cultivated rice and can be readily exploited in breeding programmes without a need for embryo rescue procedures (Lu et al. 1998; Niroula et al. 2009). O. rufipogon, a perennial and diverse wild progenitor of Asian cultivated rice, was most frequently used in yield QTL mapping studies and was found to contribute more favourable alleles than other AA genome wild species. In addition, the O. rufipogon-derived QTL are highly congruent and detected on the same chromosomal locations, and this clearly shows the consistency and accuracy of QTL detection (Swamy and Sarla 2011). The other wild progenitor species, O. nivara, is also a potential source of yield and yield-related QTL, but still largely needs to be exploited by both conventional and molecular breeding approaches.
O. nivara (Sharma et Shastry) is an annual and a close wild progenitor of Asian cultivated rice. Its accessions have abundant genetic diversity (Joshi et al. 2000; Sarla et al. 2003; Juneja et al. 2006) and contributed resistance genes for grassy stunt virus, bacterial leaf blight, blast, brown plant hopper and drought avoidance (Khush 2000; Brar and Khush 2004; Thanh et al. 2006; Ali et al. 2010), O. nivara is also a major source of cytoplasmic male sterility (Hoan et al. 1998). Because of their out crossing nature, O. nivara accessions are used extensively to study the flow of transgenes (Chen et al. 2004). Even though O. nivara has contributed significantly to pest and disease resistance in rice, it has rarely been used in yield improvement programmes.
A study was undertaken to exploit the naturally occurring alleles from O. nivara by AB-QTL mapping. Two accessions of O. nivara, IRGC81848 (Uttar Pradesh) and IRGC81832 (Bihar) from India were used, for developing mapping populations in the genetic background of the popular cultivated rice mega variety, Swarna (MTU 7029). These two accessions showed moderate molecular genetic diversity from Swarna and were genetically distinct from O. nivara accessions collected from 16 other states (Sarla et al. 2003).
The main objectives of this study were to map QTL for yield and yield-related traits, to know the proportion of trait-enhancing alleles that can be obtained from two O. nivara accessions and their congruence in the two populations, to compare the colocation of yield QTL with meta-QTL reported for yield, and to identify high-yielding BC2F5 Swarna introgression lines.
Materials and methods
Development of mapping populations
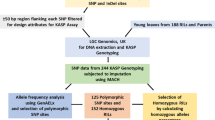
The parental material for the experiment included two accessions of O. nivara from Uttar Pradesh (IRGC81848) and Bihar (IRGC81832), a popular rainfed lowland cultivated rice variety, Swarna. An advanced backcross strategy as described by Thomson et al. (2003) was followed to develop the mapping populations. Crosses were made using Swarna as a female parent and O. nivara (IRGC81848 and IRGC81832) as a male parent. The BC2F2 population derived from Swarna × IRGC81848 is designated as population 1 and that derived from Swarna × IRGC81832 is designated as population 2.
Phenotypic evaluation of mapping populations
Two mapping populations consisting of 227 and 245 BC2F2 families were grown in two replications in an augmented block design at the Directorate of Rice Research (DRR), Hyderabad, in southern-central India during the wet season (WS) of 2005 and 2006, respectively. Each backcross family and the control (Swarna) consisted of 30 plants planted in three rows of 10 plants each, adopting a uniform spacing of 20 cm between rows and 15 cm between plants. Thirty high-yielding BC2F5 lines were evaluated in a 10-m 2 area in two replicates along with the control Swarna during WS2008 and WS2009 at DRR. Standard agronomic practices and need-based plant protection measures were adopted uniformly to raise the crop. Five plants from the middle row were used for taking the following data: days to maturity (DTM): duration in days from seeding to the time when more than 80% of the grains on the panicles were fully ripened; plant height (PH): height from the soil surface to the tip of the primary panicle was measured at the time of harvest; number of tillers per plant (NT): average number of tillers from five plants at the time of harvest; number of productive tillers per plant (NP): average number of panicle-bearing tillers at the time of harvest; number of spikelets per plant (SNP): average number of spikelets from five plants; number of filled grains per panicle (GP): average number of grains from five plants; 1000 grain weight (GW): weight of 1000 randomly selected dried grains; vegetative biomass (BM): average weight of five well-dried plants and grain yield per plant (YDLP): average weight of the dried (14% moisture) and cleaned grains from five plants.
Genotyping of the mapping populations
A set of 250 microsatellite markers uniformly distributed on all 12 chromosomes were screened for polymorphism between Swarna and O. nivara accessions, IRGC81848 and IRGC81832. A total of 100 polymorphic microsatellite markers were used to analyse the segregation in population 1, and 75 markers were used in population 2. DNA was extracted from the leaves of two-month-old seedlings using the protocol of Zheng et al. (1995). PCR for simple sequence repeat (SSR) primers was performed with 15 μL of final volume containing, 45 ng of genomic DNA, 10×buffer, 0.125 mM final concentration of each dNTP, 0.2 μM each of forward and reverse primers, 2% formamide and 1 U of biogene Taq DNA polymerase. PCR amplification was performed under the following conditions: initial denaturation at 94∘C for 5 min, followed by 35 cycles of denaturation at 94∘C for 1 min, annealing at 55∘C for 1 min, extension at 72∘C for 2 min, followed by a final extension of 72∘C for 5 min. The amplified products were checked for polymorphism or marker segregation on an agarose gel (3%) and recorded for segregating bands. BC2F5 lines were genotyped using SSR markers linked to O. nivara-derived QTL for yield and related traits.
Linkage map construction and QTL analysis
A linkage map was constructed from the genotypic data of 100 markers in population 1, and 75 markers in population 2 using Join Map 3.0 (Van Ooijen and Voorrips 2001). Linkage groups were assigned to the respective chromosomes based on the rice linkage maps developed by Temnykh et al. (2001). Both the maps were integrated to develop a consensus map using Biomercator ver. 2.0 (Arcade et al. 2004). Significant thresholds were fixed based on permutation tests at an experiment-wise significance level of 0.05 (Kosambi 1944). Based on 10,000 permutations for each trait, an average LOD value of 3.0 was used for declaring significant QTL. The QTL were identified separately for the two populations by interval mapping (IM) or composite interval mapping (CIM) using QTL Cartographer ver. 2.5 (Wang et al. 2011).
Results
Trait analysis
The frequency distribution for yield and yield-related traits is presented in figure 1. The range and number of families showing significant improvement over Swarna are presented in table 1. Transgressive segregants for Swarna were observed for seven traits in both the populations. More than 80 families (35%) showed at least 15% improvement over Swarna for plant height, number of tillers per plant, number of productive tillers per plant, and 1000-grain weight in population 1 and for number of tillers per plant, 1000-grain weight, and vegetative biomass in population 2. All the traits followed a near normal distribution except plant height and number of productive tillers per plant in both the populations (figure 1). Correlations between trait pairs were estimated and are presented in table 2. Eighteen correlations in population 1 and 22 correlations in population 2 were found to be significant, and these were observed between days to maturity and number of tillers per plant, number of productive tillers per plant, number of spikelets per plant, and yield per plant; plant height and 1000-grain weight and vegetative biomass; between number of tillers per plant and number of productive tillers per plant; and between number of spikelets per plant and number of filled grains per plant.



Frequency distribution of nine yield and related traits in two O. nivara derived populations. Bars in white indicates population 1; bars in black indicates population 2; arrow indicates Swarna values.
Marker analysis
A parental polymorphism survey using 250 microsatellite or SSR markers revealed that 120 (48%) were polymorphic between IRGC81848 and Swarna, and 108 (44%) were polymorphic between IRGC81832 and Swarna. One hundred polymorphic SSR markers were used for genotyping 227 BC2F2 families in population 1 and 75 SSR markers were used for genotyping 245 families in population 2. The overall mean introgression of O. nivara alleles ranged from 2.6 to 38%, with an overall mean introgression of 16.6% in both the populations. Introgression percentage varied from chromosome to chromosome and between the families in both populations.
QTL analysis
In all, 28 QTL were identified in population 1 and 26 QTL in population 2 (table 3; figure 2). Of these, 22 (78%) and 18 (69%) QTL were derived from O. nivara and had a trait-increasing effect. Details on only the significant major QTL identified for each trait are given below.

Chromosomal location of QTL for yield and yield-related traits. QTL underlined are from population 2. QTL in bold are derived from O. nivara.
Days to maturity (DTM)
Three QTL were detected in population 1 and two QTL in population 2. All had an increasing effect from Swarna.
Plant height (PH)
Three QTL were detected in each population. All were derived from O. nivara. The phenotypic variance varied from 5 to 63% in population 1 and from 7 to 46% in population 2. Each of these QTL contributed more than 5% to the total phenotypic variance (PV) and the additive effect varied from 5 to 39 cm.
Number of tillers per plant (NT)
Two QTL (nt5.1 and nt12.1) were detected in population 1 and four QTL (nt2.1, nt3.1, nt11.1 and nt12.1) in population 2. All the QTL except nt2.1 was derived from O. nivara. These QTL contributions varied from 6 to 37% to the total PV and also showed a high additive effect.
Number of productive tillers per plant (NPT)
Three QTL were detected in each population; all were derived from O. nivara except npt12.2. Two QTL (npt6.1 and npt9.1) from O. nivara contributed 36 and 30%, respectively, to the total PV. These also had a high additive effect on the trait (18 and 30).
Number of spikelets per plant (NSP)
Three QTL were detected in each population; all were derived from O. nivara except nsp12.1. QTL nsp2.1 was detected at high LOD (6.0). This explained the high PV (15%) with an additive effect of 367 spikelets per plant.
Number of filled grains per plant (NFG)
Three QTL were detected in population 1 and two QTL in population 2. All the QTL in population 1 were derived from O. nivara. The nfg2.1 from O. nivara was detected at high LOD (6.0) and had PV of 15% with an additive effect of 333 grains per plant. In population 2, nfg1.1 was derived from O. nivara and had a PV of 10% with an additive effect of 274 grains per plant.
Thousand-grain weight (GW)
Three QTL were detected for 1000-grain weight in each population. In population 1, all the QTL were derived from Swarna. In population 2, two QTL were derived from O. nivara, gw2.1 contributed 9% to the total PV and added 1 g to grain weight.
Vegetative biomass (BM)
Four QTL were detected in population 1, and all were derived from O. nivara. Two QTL (bm2.1 and bm9.1) contributed more than 10% to the total PV and also had a high additive effect on the trait (169 and 121 g). Two QTL (bm5.1 and bm11.1) were detected in population 2; both were derived from O. nivara and had a PV of 5 and 10%, and an additive effect of 26 and 32 g, respectively.
Yield per plant (YLDP)
In population 1, four QTL were detected and all were derived from O. nivara. Three QTL (yldp2.1, yldp9.1, and yldp11.1)each influenced the trait with more than 10% PV and the additive effects of these QTL were 7.2, 16.6, and 4.1 g, respectively. In population 2, four QTL were detected; all were derived from O. nivara. Three QTL (yldp2.1, yldp8.1, and yldp11.1) each contributed more than 10% to the total PV and added 6, 12, and 8 g to the yield per plant.
Discussion
Even though wild progenitor species are phenotypically inferior, they harbour many superior alleles which have been left behind during domestication and are often masked by the overall poor genetic background. These superior alleles upon transferring to elite genetic background can further enhance the yield potential of elite rice varieties (Wang et al. 2005). In several earlier studies using wild progenitor species as donor parents, superior and favourable alleles have been identified (Xiao et al. 1998; Thomson et al. 2003; Tian et al. 2005; Rahman et al. 2008). In our study, in both the O. nivara derived populations, transgressive segregants with more than 15% improvement over Swarna. These transgressive segregants are important evidence of the favourable effects of introgressions from wild species.
In general, about half of the QTL mapped from wild species are reported to be favourable (Xiao et al. 1998; Septiningsih et al. 2003; Thomson et al. 2003; Marri et al. 2005; Yoon et al. 2006; McCouch et al. 2007; Swamy and Sarla 2008). The O. nivara accessions also contributed trait-enhancing alleles at 78% of the loci in population 1 and at 69% of the loci in population 2 in the Swarna genetic background. The results from a parental polymorphism survey also supports this hypothesis. O. nivara accessions showed 15% less polymorphism with O. sativa in comparison with the polymorphism of O. rufipogon with O. sativa (McCouch et al. 2007). The correlations among the traits in both populations were similar, along with the direction of the effect of QTL. Most of the correlated traits were colocated and their QTL were found on the same chromosomal locations, indicating their conservation in the two O. nivara accessions.
Several O. nivara-derived QTL with a major effect on yield and related traits were identified in both the populations. The major effect yield QTL yldp9.1 was also found to have pleiotropic effect on several traits. In another O. rufipogon-derived population, this chromosomal region has been fine mapped to a 37.4 kb length in which seven traits (plant height, heading date, panicle length, spikelets per panicle, spikelet density, grains per panicle and grain weight) were colocated. Hence, this is a priority locus for map-based cloning, candidate gene identification and allele mining to discover novel naturally occurring variations for rice improvement (Marri et al. 2005; Xie et al. 2008).
Two yield QTL (yldp2.1 and yld11.1) were detected in both populations and these were colocated with QTL for number of spikelets per plant, number of filled grains per plant and vegetative biomass, and the trait enhancing alleles were from O. nivara. Thus, at this locus, many positive alleles were identified from O. nivara. QTL for yield have been reported previously at the same chromosomal region from O. rufipogon (Xiao et al. 1998; Septiningsih et al. 2003; Liu et al. 2011). One interesting observation is that QTL for number of spikelets per plant were found to colocate with both yldp9.1 and yldp2.1. If we consider colocation of QTL equivalent to coexpression and compare yldp9.1 and yldp2.1, it is clear that at both loci the O. nivara allele increases the number of spikelets per plant. The other significant grain yield QTL (yldp3.1, yldp8.1 and yldp12.1) that increased yield per plant in specific populations is important in QTL pyramiding programs. All the QTL for number of spikelets per plant were derived fromO. nivaraexcept nsp12.1. The most significant QTL for spikelets per plant (nsp1.1) was detected at the same chromosomal location in both populations and also in several O. rufipogon-derived populations (Xiao et al. 1998; Septiningsih et al. 2003; Thomson et al. 2003; Marri et al. 2005). We noticed that major-effect QTL for number of spikelets per panicle have been frequently reported on chromosomes 1 and 7 (Xiong et al. 1999; Tian et al. 2005; Li et al. 2006; Xing et al. 2008). QTL on chromosome 1 was found in both O. nivara-and O. sativa-derived populations, but the one on chromosome 7 was found only in O. rufipogon-derived populations. If these QTL from different sources are pooled by pyramiding, the number of spikelets can be improved significantly.
The major-effect QTL nfg1.1 has also been detected in several earlier mapping studies that involved O. nivara and O. rufipogon as donor parents (Marri et al. 2005; Xiao et al. 1998; Septiningsih et al. 2003; Wickneswari et al. 2012). In most instances, this QTL was co-located with QTL for number of spikelets per plant and it is a worthy locus for marker-assisted breeding to improve the yield of rice varieties, but it has to be ascertained whether this locus has a pleiotropic effect or tight linkage. The numbers of spikelets and grains per plant have a significant influence on yield; up to 18% improvement in yield was observed in several O. rufipogon-derived lines by an increased number of spikelets and grains per plant (Xiao et al. 1998; Xiong et al. 1999). The yield advantage in an O. rufipogon-derived popular rice variety, Dhanarsi, is also because of its higher number of grains per panicle (251 grains) and this variety yielded 6.48 t/ha (Ram et al. 2007).
Oryza nivara has high vegetative biomass and all the QTL for vegetative biomass were derived from O. nivara. Interestingly, only one QTL (bm11.1)was identified at the same locus in both the populations. There are no previous reports of biomass QTL mapped from wild species. The increase in the number of spikelets per plant coupled with higher biomass led to an increase in number of filled grains in the O. nivara-derived progenies.
The major plant height QTL (ph1.1) was dentified in both the populations. At this chromosomal location, QTL for plant height have been frequently reported in earlier studies using wild species (Xiao et al. 1998; Brondani et al. 2001; Septiningsih et al. 2003; Thomson et al. 2003; Wickneswari et al. 2012). It is also interesting to note that this QTL region is near the the sd1 gene, which is also popularly called the green revolution gene (Cho et al. 1994). Detecting new major-effect QTL for plant height is useful in diversifying the genetic base of the green revolution rice varieties, which have the sd1 gene. Further dissection of this locus provides a basis for comparative mapping and allele mining to detect a new source of plant height genes from distantly related cereal crops.
The nt5.1 detected in population 1 is congruent with the tiller number QTL reported in O. rufipogon (Marri et al. 2005; Wickneswari et al. 2012), while nt12.1 was detected in both the populations. We report two new QTL (npt6.1 and npt9.1) from O. nivara with a phenotypic variance of > 30%. QTL npt6.1 has been mapped to a region near the monoculm gene (MOC1), which is important for tillering in rice (Li et al. 2003). Such congruency of QTL/genes for a trait from different populations provide confidence in the accuracy of QTL mapping (Price 2006). It can also help in deciphering the role of candidate genes when alleles derived from different wild species are used in overexpression or loss-of-function studies.
A comparison was made between the yield QTL derived from O. nivara with meta-QTL for yield (Swamy and Sarla 2011). The major-effect yield QTL (yldp2.1 and yldp11.1) in both populations coincided with MQTL2.3 and MQTL11.1, respectively. QTL yldp9.1 and yldp12.1 found in population 1 coincided with MQTL9.1 and MQTL12.1, whereas yldp3.1 and yldp8.1 identified in population 2 coincided with MQTL3.3 and MQTL8.1, respectively. Thus, a remarkable extent of colocalization was observed in yield QTL from O. nivara and O. rufipogon.
The results clearly show that yield-enhancing QTL are commonly detected at the same chromosomal locations derived from different wild species but their effect varied across genetic backgrounds (McCouch et al. 2007; Swamy and Sarla 2008). It can also be noted that small phenotypic variance values observed in BC2F2 populations may increase in subsequent generations of backcrossing as reported for grain weight QTL gw9.1 (Xie et al. 2008). They reported an increase from 12% in BC2F2 to 42% in BC3F3 and up to 51% in BC3F4. The message is that QTL explaining low PV in BC2F2 should not be ignored which may turn out to be ultimately useful for MAS if the PV increases in subsequent generations. The reverse is also true as shown for the effect of root QTL on grain yield in near isogenic lines (NILs) of Azucena in Kalinga III. Here, the NILs with non-target introgressions performed better than NILs with targeted introgression in subsequent generations (Steele et al. 2007). These two instances emphasize the fact that most QTL are context-specific and the QTL effect may change as the background genetic milieu changes in each backcross generation.
In BC2F5 lines derived from this mapping population, nearly 20% increase in yield was obtained in some lines in small-scale (125 plants in 5 rows) replicated field trials. The increase was largely associated with the increased spikelets per panicle and number of filled grains per plant. The preponderance of yield QTL-associated markers from O. nivara in the 20 high-yielding lines confirms their role in a yield increase (data not shown). If these few high-priority, trait-increasing, major-effect QTL are pooled, it may lead to increased yield even in the presence of extensive phenotypic buffering that often takes place (Mijalski et al. 2005). Yet, a yield increase of 15-20% can be easily expected. The evidence for this is the introgression line IET21542 (derived from Swarna x O. nivara IRGC 81848), which was successfully evaluated for three years of multi-location testing in the All India Coordinated Rice Improvement Project irrigated medium duration trials and released as DRR Dhan 40 for cultivation in three important rice-growing states of India, namely, West Bengal, Maharashtra, and Tamil Nadu. It had a mean yield of 5.5 t/ha over three years and a maximum paddy yield of 10.6 t/ha in the 2011 wet season (DRR AICRIP report on varietal improvement, 2012).
Conclusion
In this study, O. nivara accessions have contributed trait enhancing alleles for several yield and yield-related traits in the back ground of popular rice variety Swarna, indicating their potential use in yield improvement programs. We identified several major effect QTL derived from O. nivara, which are reliable for MAS, fine mapping and candidate gene identification. The high yielding BC2F5 lines clearly show that the yield and adaptability of rice varieties can be improved considerably using closely-related wild species in crosses with farmer-adapted elite cultivars followed by the advanced backcross method. Several O. nivara derived introgression lines developed in this work will benefit breeding efforts to overcome the adverse effects of climate change and help sustain global rice production.
References
Ali M. L., Sanchez P. L., Yu S., Lorieux M. and Eizenga G. C. 2010 Chromosome segment substitution lines: a powerful tool for the introgression of valuable genes from Oryza wild species into cultivated rice (O. sativa). Rice 3, 218–234.
Arcade A., Labourdette A., Falque M., Mangin B., Chardon F., Charcosset A. et al. 2004 BioMercator: integrating genetic maps and QTL towards discovery of candidate genes. Bioinformatics 14, 2324–2326.
Brar D. S. and Khush G. S. 2004 Alien introgression in rice. Plant Mol. Biol. 35, 35–47.
Brondani C., Rangel P. H. N., Brondani R. P. V. and Ferreira M. E. 2001 QTL mapping and introgression of yield-related traits from Oryza glumaepatula to cultivated rice (Oryza sativa) using microsatellite markers. Theor. Appl. Genet. 104, 1192– 1203.
Chen L. J., Lee D. S., Song Z. P., Suh H. S. and Lu B. R. 2004 Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives. Ann. Bot. 93, 67–73.
Cho Y. G., Eun M. Y., McCouch S. R. and Chae Y. A. 1994 The semi dwarf gene sd-1 of rice (Oryza sativa L.). II. Molecular mapping and marker-assisted selection. Theor. Appl. Genet. 89, 54–59.
. Y., Wang Y. M., Zhang Z. J., Shen Y., Lin X. Y., Ou X. F. et al. 2006 Extent and pattern of DNA methylation alteration in rice lines derived from introgressive hybridization of rice and Zizania latifolia Griseb. Theor. Appl. Genet. 113, 196–205.
Gur A. and Zamir D. 2004 Unused natural variation can lift yield barriers in plant breeding. PLOS Biol. 2, e245.
Hoan N. T., Sarma N. P. and Siddiq E. A. 1998 Wide hybridization for diversification of CMS in rice. Int. Rice Res. Notes 23, 5–6.
Joshi S. P., Gupta V. S., Aggarwal R. K., Ranjekar P. K. and Brar D. S. 2000 Genetic diversity and phylogenetic relationship as revealed by inter simple sequence repeat (ISSR) polymorphism in the genus Oryza. Theor. Appl. Genet. 100, 1311–1320.
Das A., Joshi S. V., Sharma S., Yogesh V. Y., Patra B. C. et al. 2006 Oryza nivara (Sharma et Shastry), the progenitor of O. sativa (L.) subspecies indica harbors rich genetic diversity as measured by SSR markers. Curr. Sci. 91, 1079–1085.
Khush G. S. 2000 Rice germplasm enhancement at IRRI. Phillipp. J. Crop Sci. 25, 45–51.
Kosambi D. D. 1944 The estimation of map distances from recombination values. Ann. Eugen. 12, 172–175.
Li X., Qian Q., Fu Z., Wang Y., Xiong G., Zeng D. et al. 2003 Control of tillering in rice. Nature 422, 618–621.
Li C., Zhou A. and Sang T. 2006 Genetic analysis of rice domestication syndrome with the wild annual species, Oryza nivara. New Phytol. 170, 185–194.
Liu T., Li L., Zhang Y., Xui C., Li X. and Xing Y. 2011 Comparison of quantitative trait loci for rice yield, panicle length and spikelet density across three connected populations. J. Genet. 90, 377–382.
Lu B. R., Naredo M. E. B., Juliano A. B. and Jackson M. T. 1998 Taxonomic status of Oryza glumaepatula Steud. III. Assessment of genomic affinity among AA genome species from the New World, Asia and Australia. Genet. Res. Crop Evol. 45, 215– 223.
Marri P. R., Sarla N., Reddy V. L. N. and Siddiq E. A. 2005 Identification and mapping of yield-related QTL from an Indian accession of Oryza rufipogon. BMC Genet. 6, 33.
McCouch S. R., Sweeney M., Li J., Jiang H., Thomson M., Septiningsih E. et al. 2007 Through the genetic bottleneck: O. rufipogon as a source of trait-enhancing alleles for O. sativa. Euphytica 154, 317–339.
Mijalski T., Mijalski A., Harder T., Halder M., Kersten M., Horsch T. et al. 2005 Identification of coexpressed gene clusters in a comparative analysis of transcriptome and proteome in mouse tissues. Proc. Natl. Acad. Sci. USA 102, 8621–8626.
Niroula R. K., Subedi L. P. and Upadhyay M. P. 2009 Cytogenetic analyses of intragenomic rice hybrids derived from Oryza sativa L. and O. nivara Sharma et Shastry. Bot. Res. Int. 4, 277–283.
Price A. H. 2006 Believe it or not QTL are accurate. Trends Plant Sci. 11, 213–216.
Quan C. Z., You Y. F., Qing L. D., Qiong Y. T., Jian F., Jun Y. H. et al. 2012 Genetic diversity of wild rice species in Yunnan province of China. Rice Sci. 19, 21–28.
Rahman M. L., Chu S. H., Choi M., Qiao Y. L., Jiang W., Piao R. et al. 2008 Identification of QTL for some agronomic traits in rice using an introgression line from Oryza minuta. Mol. Cells 24, 16–26.
Ram T., Majumder N. D., Krishnaveni D. and Ansari M. M. 2007 Rice variety, Dhanrasi, an example of improving yield potential Rice variety, Dhanrasi, an example of improving yield potential (Oryza rufipogon). Curr. Sci. 92, 987–992.
Sarla N., Bobba S. and Siddiq E. A. 2003 ISSR and SSR markers based on AG and GA repeats delineate geographically diverse Oryza nivara accessions and reveal rare alleles. Curr. Sci. 84, 683–690.
Septiningsih E. M., Prasetiyono J., Lubis E., Tai T. H., Tjubaryat T., Moeljopawiro S. et al. 2003 Identification of quantitative trait loci for yield and yield components in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor. Appl. Genet. 107, 1419– 1432.
Steele K. A., Virk D. S., Kumar R., Prasad S. C. and Witcombe J. R. 2007 Field evaluation of upland rice lines selected for QTL controlling root traits. Field Crops Res. 101, 180–186.
Swamy B. P. M. and Sarla N. 2008 Yield enhancing quantitative trait loci (QTL) from wild species. Biotechnol. Adv. 26, 106– 120.
Swamy B. P. M. and Sarla N. 2011 Meta-analysis of yield QTL derived from inter-specific crosses of rice reveals consensus regions and candidate genes. Plant. Mol. Biol. Rep. (doi: 10.1007/s11105-010-0274-1).
Tanksley S. D. and Nelson J. C. 1996 Advanced backcross QTL analysis; a method for simultaneous discovery and transfer of valuable QTL from unadapted germplasm into elite breeding lines. Theor. Appl. Genet. 92, 191–203.
Tanksley S. D. and McCouch S. R. 1997 Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277, 1063–1066.
Temnykh S., DeClerck G., Lukashova A., Lipovich L., Cartinhour S. and McCouch S. R. 2001 Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 11, 1441–1452.
Thanh P. T., Sripichitt P., Chanprame S. and Peyachoknagu S. 2006 Transfer of drought resistant character from wild rice (Oryza meridionalis and Oryza nivara) to cultivated rice (Oryza sativa L.) by backcrossing and immature embryo culture. Kasetsart J. 40, 582–594.
Thomson M. J., Tai T. H., McClung A. M., Lai X. H., Hinga M. E., Lobos K. B. et al. 2003 Mapping quantitative trait loci for yield, yield components and morphological traits in an advanced backcross population between Oryza rufipogon and the Oryza sativa cultivar Jefferson. Theor. Appl. Genet. 107, 479–493.
Tian F., Li D. J., Fu Q., Zhu Z. F., Fu Y. C., Wang X. K. et al. 2005 Construction of introgression lines carrying wild rice (Oryza rufipogon Griff.) segments in cultivated rice (Oryza sativa L.) background and characterization of introgressed segments associated with yield-related traits. Theor. Appl. Genet. 112, 570– 580.
Van Ooijen J. W. and Voorrips R. E. 2001 JoinMap 3.0 software for the calculation of genetic linkage maps. Plant Research International, Wageningen, the Netherlands.
Wang Y. M., Dong Z. Y., Zhang Z. J., Lin X. Y., Shen Y., Zhou D. et al. 2005 Extensive de novo variation in rice induced by introgression from wild rice (Zizania latifolia). Genetics 170, 1945–1956.
Wang S., Basten C. J. and Zen Z. B. 2011 Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC.
Wickneswari R., Bhuiyan M. A. R., Kalluvettankuzhy S., Lim L. S., Thomson M. J., Narimah M. K. et al. 2012 Identification and validation of quantitative trait loci for agronomic traits in advanced backcross breeding lines derived from Oryza rufipogon × Oryza sativa cultivar MR219. Plant Mol. Biol. Rep. (doi: 10.1007/s11105-011-0404-4).
Xiao J, Li J., Grandillo S., Ahn S. N., Yuan L., Steven D. et al. 1998 Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics 150, 899–909.
Xie X., Song H. M., Jin F., Ahn S., Suh J., Hwang H. G. et al. 2008 Fine mapping of a grain weight quantitative trait locus on rice chromosome 8 using near-isogenic lines derived from a cross between Oryza sativa and Oryza rufipogon. Theor. Appl. Genet. 113, 885–894.
Xing Y. Z., Tang W. J., Xue W. Y., Xu C. G. and Zhang Q. 2008 Fine mapping of a major quantitative trait loci, qSSP7, controlling the number of spikelets per panicle as a single Mendelian factor in rice. Theor. Appl. Genet. 116, 789–796.
Xiong L. Z., Liu K. D., Dai X. K., Xu C. G. and Zhang Q. 1999 Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon. Theor. Appl. Genet. 98, 243– 251.
Yano M. 2001 Genetic and molecular dissection of naturally occurring variation. Curr. Opin. Plant Biol. 4, 130–135.
Yoon D. B., Kang K. H., Kim H. J., Ju H. G., Kwon S. J. and Suh J. P. 2006 Mapping quantitative trait loci for yield components and morphological traits in an advanced backcross population between Oryza grandiglumis and the O. sativa japonica cultivar Hwaseongbyeo. Theor. Appl. Genet. 12, 1052–1062.
Zheng K., Subudhi P. K., Domingo J., Magpantay G and Huang N. 1995 Rapid DNA isolation for marker assisted selection in rice breeding. Rice Genet. News Lett. 12, 255–258.
Acknowledgements
BPMS thank the University Grants Commission–Council of Scientific and Industrial Research (UGC-CSIR) for providing fellowship; KK thank the Indian Council of Agricultural Research (ICAR), Government of India, for fellowship. This project was also supported by the Department of Biotechnology, Government of India.
Author information
Authors and Affiliations
Corresponding author
Additional information
Swamy B. P. M., Kaladhar K., Reddy G. A., Viraktamath B. C. and Sarla N. 2014 Mapping and introgression of QTL for yield and related traits in two backcross populations derived from Oryza sativa cv. Swarna and two accessions of O. nivara. J. Genet. 93, xx–xx
Rights and permissions
About this article

Cite this article
MALLIKARJUNA SWAMY, B.P., KALADHAR, K., REDDY, G.A. et al. Mapping and introgression of QTL for yield and related traits in two backcross populations derived from Oryza sativa cv. Swarna and two accessions of O. nivara . J Genet 93, 643–654 (2014). https://doi.org/10.1007/s12041-014-0420-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12041-014-0420-x