
Abstract
Nociceptin/orphanin FQ (N/OFQ) is an opioid-like neuropeptide that binds and signals through a G-protein-coupled receptor called the N/OFQ peptide (NOP) receptor. N/OFQ and the NOP receptor are expressed in the midbrain and have been implicated in the pathogenesis of Parkinson’s disease (PD). Genetic removal of the N/OFQ precursor partially protects midbrain dopaminergic neurons from 1-methyl-4-phenylpyridine-induced toxicity, suggesting that endogenous N/OFQ may be detrimental to dopaminergic neurons. However, whether N/OFQ directly affects the survival and growth of dopaminergic neurons is unknown. Here, we show that N/OFQ has a detrimental effect on the survival of dopaminergic neurons and the growth of their axons in primary cultures of the E14 rat ventral mesencephalon. N/OFQ potentiates the effects of the neurotoxins 6-hydroxydopamine and 1-methyl-4-phenylpyridinium through p38-MAPK signalling. We also show that like α-synuclein, there is a significant reduction in N/OFQ messenger RNA (mRNA) expression in the midbrain of patients with Parkinson’s disease. These results demonstrate for the first time that N/OFQ is detrimental to the survival and growth of dopaminergic neurons and that its expression is altered in the midbrain of patients with Parkinson’s disease.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disorder, which affects 0.5 to 1 % of those 65–69 years of age and 1 to 3 % of people over 80 years of age [1, 2]. PD is characterized by motor deficits, including akinesia, bradykinesia and resting tremor, and non-motor deficits, including cognitive and autonomic disturbances [1, 3]. The progressive degeneration of midbrain dopaminergic (DA) neurons in the substantia nigra pars compacta (SNpc) and the loss of their axonal terminals in the striatum is the classical neuropathological feature of this disease [4–6]. A number of post-mortem studies have shown that the motor symptoms of PD appear when there is a ~30 % loss of total SN neurons [7–9]. In a PET study of striatal dopaminergic terminal loss, no motor symptoms were apparent with a 51 and 56 % reduction in vesicular monoamine transporter ligand and a dopamine transporter ligand, respectively, however, the motor symptoms of PD became apparent when these markers were reduced by 62 and 71 %, respectively [10]. This pre-symptomatic lag phase has been proposed to be due to sprouting of remaining DA neurons [11, 12] or to adaptive changes in non-DA structures [13]. Although 5–10 % of PD cases are of a genetic aetiology, the remaining 90–95 % of cases are of idiopathic origin [14, 15]. Thus, understanding the molecular and cellular mechanisms that regulate DA neuron survival, and the growth and maintenance of their axons is crucial to develop new understanding of the causative basis of their degeneration in PD and new therapies for the disorder. Various pre-clinical and clinical studies suggest an involvement of Nociceptin/orphanin FQ (N/OFQ) in the pathogenesis of PD; however, whether N/OFQ directly affects DA neuronal survival or axonal growth is unknown.
N/OFQ [16, 17] is a neuropeptide belonging to the opioid family [18, 19] and is the endogenous ligand of the N/OFQ peptide (NOP) receptor [20]. The N/OFQ-NOP receptor system is expressed throughout the central nervous system [21, 22] and is involved in the modulation of a variety of central functions such as pain perception, mood, cognition, food intake, reward and locomotion [18, 19, 23]. We have previously demonstrated that N/OFQ also contributes to motor impairment in PD [24]. Indeed, an increase in N/OFQ levels is observed in the brain of Parkinsonian rodents [24, 25] and the CSF of PD patients [25]. Furthermore, NOP receptor antagonists reverse motor impairment in rodent and non-human primate [26, 27] models of the disease. A recent NOP-eGFP reporter mouse has shown that NOP is expressed throughout the neuronal cell body and in the neuronal processes [28], and NOP messenger RNA (mRNA) was found in DA neurons in the SN [29, 30]. The source of N/OFQ in the midbrain is less clear, but N/OFQ is expressed by both neurons and astrocytes [31, 32]. Specifically in SNc/VTA, about half of N/OFQ-positive neurons also expressed GAD, suggesting that they are GABAergic [30]. The role of N/OFQ in PD, however, may go beyond its involvement in sustaining motor symptoms. Indeed, N/OFQ knockout mice were partially protected from the neurotoxic action of 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine (MPTP), with a higher number of DA neurons and striatal tyrosine-hydroxylase (TH)-positive terminals spared after acute neurotoxin injection with respect to controls [24, 33], suggesting that N/OFQ also contributes to the neurodegeneration associated with PD. Interestingly, no neuroprotection was observed in N/OFQ knockout mice after injection of methamphetamine [33], which is known to destroy striatal DA terminals and DA neurons in a retrograde manner, possibly indicating that N/OFQ neurotoxic action is accomplished in SNpc, where DA neurons are located. In fact, high levels of N/OFQ are found in both SNpc and SN pars reticulata (SNpr) [34]. It could be speculated that N/OFQ is neurotoxic through indirect mechanisms, e.g. via releasing glutamate [35] or pro-inflammatory mediators (for a review see [36]). However, SNpc DA neurons express the NOP receptor [21, 22], and although N/OFQ exerts acute inhibitory effects on the activity of SNpc DA neurons [37], the possibility that prolonged exposure to N/OFQ can exert direct, slower neurotoxic actions cannot be ruled out. In fact, N/OFQ [38], and more generally opioids [39], not only act through the canonical Gi pathway but also through the MAP-kinase pathway, most likely recruited by the βγ subunits [38]. The involvement of MAP-kinase signalling in the degeneration of DA neurons is well established, and administration of the DA neurotoxin 6-hydroxydopamine (6-OHDA) leads to the activation of p38-MAPK signalling in DA neurons [40].
Therefore, the aim of this study was to determine if N/OFQ exerts a direct detrimental effect on DA neuronal survival and axonal growth, and whether there were alterations in N/OFQ and/or its receptor in the human Parkinsonian midbrain. To do this, the SH-SY5Y neuroblastoma cell line, a widely used and well-validated model of human midbrain DA neurons [41], and primary cultures of embryonic rat ventral mesencephalon (VM) [42, 43] were used. Both the SH-SY5Y cell line [44, 45] and midbrain DA neurons [21, 22] express the NOP receptor. Analysis of N/OFQ and NOP expression in the Parkinsonian midbrain was performed using datasets derived from the gene expression omnibus of previously published microarray data of the SN of PD patients.
Methods
Preparation and Treatment of SH-SY5Y Cell Cultures
SH-SY5Y cells were maintained in Dulbecco’s Modified Eagle Medium:F12 (DMEM:F12; Sigma), supplemented with 10 % foetal calf serum (FCS; Sigma), 100 nM l-Glutamine (Sigma), 100 U/ml Penicillin (Sigma), and 10 μg/ml Streptomycin (Sigma). The cells were incubated under a humidified atmosphere containing 5 % CO2 at 37 °C and were treated following 1 day in vitro (DIV) with 0, 5, 10, 15, 25, 30, 35, 50 or 100 μM 6-OHDA (pre-stabilized in 0.01 % ascorbic acid; Sigma) with/without 0.1, 1, 3 or 10 μM N/OFQ, 3 μM UFP112 (NOP receptor agonist [46]), 10 μM UFP101 (NOP receptor antagonist [47]) or 3 μM SB-612111 (NOP receptor antagonist; [48, 49]) for 24 h before analysis of cell viability using an MTT and lactate dehydrogenase (LDH) assay.
MTT Assay
The MTT assay was used to determine the number of viable cells in SH-SY5Y cultures. The assay is based on the cleavage of the yellow tetrazolium salt [3-(4,5-dimethylthiazol-2yl)-2,5-diphenyl tetrazolium bromide] (MTT) to purple formazan crystals by metabolically active cells. Following the treatments, 0.5 mg/ml of MTT labelling reagent was added to each well, and plates were incubated at 37 °C for 4 h. Cultures were then solubilized in dimethyl sulfoxide (DMSO), and the spectrophotometric absorbance of the samples was detected by a microtiter plate reader. The wavelength to measure absorbance of formazan product is 570 nm, with a reference wavelength of 750 nm.
LDH Assay
The LDH cytotoxicity assay was used to quantitatively measure LDH released into the media from damaged cells as a biomarker for cellular cytotoxicity and cytolysis. All the reactions were performed at room temperature, and samples in each experimental group were prepared in triplicate. Briefly, after seeding cells in a 96-well flat bottom microtiter plate, lysis solution was added to the latter, and after a 45-min incubation in a humidified 37 °C, 5 % CO2 atmosphere, 50 μl of culture supernatant from each well was collected and transferred to a new microtiter plate. Fifty microliters of LDH assay buffer was added to the supernatants, and the plate was gently agitated for 30 s to allow reagents’ mix. After a 10–30-min incubation at room temperature, 50 μl of the stop solution was added to all the wells and, after a gentle agitation, absorbance was measured between 490 and 520 nm.
Preparation of E14 Rat Ventral Mesencephalon (VM) Cultures
VM cultures were prepared from embryonic (E) day 14 embryos of time-mated Sprague-Dawley rats (Biological Services Unit, University College Cork) as previously described [42]. Briefly, dissociated VM neurons were seeded at a density of 1 × 105 cells per poly-d-lysine-coated 24-well tissue culture dish in DMEM:F12 (Sigma) supplemented with 1 % penicillin/streptomycin (Sigma), 1 % l-glutamine (Sigma), 1 % FCS (Sigma) and 2 % B27 (Invitrogen). Cultures were treated for 24 h with either 6-OHDA (0-30 μM) or MPP+ (0-30 μM) or N/OFQ (0-500nM) 2 h after plating, and cultures were incubated under a humidified atmosphere containing 5 % CO2 at 37 °C before being processed for immunocytochemistry. All experiments were repeated at least three times with three individual replicates per group, per experiment.
Immunocytochemistry
At the end of the experiment, cells were washed in pre-warmed Hank’s balanced salt solution (HBSS) and fixed with ice-cold methanol for 10 min at −20 °C followed by 4 % paraformaldehyde for 15 min at room temperature and washed in 10 mM PBS with 0.02 % Triton (Sigma) (PBST) three times for 5 min. To block non-specific antibody binding, cells were incubated in 5 % bovine serum albumin (BSA) in 10 mM PBS for 1 h at room temperature. Cells were then incubated in an antibody against tyrosine hydroxylase (TH; 1:200; mouse monoclonal; Millipore) in 1 % BSA in 10 mM PBS for 16 h at 4 °C. The cells were washed three times in PBST and incubated with Alexa Fluor 594-conjugated donkey anti-mouse IgG (1:2000; Invitrogen) in 1 % BSA in PBS for 1 h at 20 °C. Following three washes in PBST, cultures were imaged under an Olympus IX70 Provis Inverted microscope fitted with an Olympus DP70 camera and AnalysisDTM software. For preparation of figures, representative photomicrographs were converted to grayscale and inverted.
Analysis of Neuronal Complexity
Images of the neurite arbours of individual DA neurons were used to analyse the Sholl profiles of TH-positive cells using Sholl analysis as previously described [50]. Morphological analysis of total neurite length and number of branch points were carried out as previously described [51]. Briefly, neurite length was calculated using the formula n × T × π/2, where n = the number of times the neurites intersect the grid lines and T = the distance between the gridlines taking the magnification into account.
Statistical Analyses
Statistical analyses were performed using unpaired Student’s t test or one-way analysis of variance (ANOVA) with a post hoc Tukey’s, Newman-Keuls’ or Dunnett’s test as appropriate. Data were expressed as mean plus and minus standard error of the mean (mean ± SEM) and deemed significant when p < 0.05.
Microarray Gene Expression
Microarray gene expression data comparing SN from normal and age-matched PD patients derived from three independent studies was downloaded from Gene Expression Omnibus (GEO, series GSE20295 [52], GSE8397 [53] and GSE7621 [54]). RMA or TMM-normalized probe expression values were summarized to Ensembl Gene IDs by averaging individual probe values annotated to the same Ensembl ID (probes that matched more than one gene were removed from the analysis). The resulting pool of 12,210 genes with expression data in all three datasets was subsequently quantile-normalized before extracting expression values for PNOC, OPRL, SNCA and MAPT. The expression values of these genes were normalized against total RNA to allow comparison across samples. Statistical comparisons between control and disease-derived samples were carried using three separate approaches: RANK products [55], t test (parametric) and Wilcoxon test (non-parametric). Charts show the mean ± SEM for the indicated genes comparing all resulting control (n = 38) and PD samples (n = 51).
Results
N/OFQ Potentiates the Neurotoxic Effect of 6-OHDA in SH-SY5Y Cells
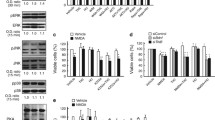
To examine if N/OFQ affected cell viability, we first used the SH-SY5Y human neuroblastoma cell line, which has been used as a model of human midbrain dopaminergic neurons [56]. A dose-response experiment was initially performed to determine a concentration of 6-OHDA which was submaximal in its neurotoxic effect. SH-SY5Y cells were cultured in increasing concentrations of 6-OHDA (0-100 μM) for 24 h after which time viability was assessed by MTT assay. Increasing concentrations of 6-OHDA resulted in a dose-dependent reduction in cell viability (Fig. 1a). A 30-μM concentration of 6-OHDA was selected for subsequent experiments as this resulted in a 50 % reduction in cell viability, thus facilitating examination of any additive effects of N/OFQ.

N/OFQ potentiates the effects of 6-OHDA on SH-SY5Y cells. a SH-SY5Y cells were incubated for 24 h with increasing concentrations (5–100 μM) of 6-OHDA after which cell viability was quantified using an MTT assay. b MTT assay showing the effect of N/OFQ (1–10 μM), with or without 30 μM 6-OHDA on SH-SY5Y cells viability for 24 h. c MTT assay and d LDH assay examining the effect of the NOP agonist UFP112 (3 μM) on the neurotoxic effect of 30 μM 6-OHDA after 24 h and whether this was prevented by the NOP antagonists UFP101 (10 μM) and SB612111 (3 μM). Data are means ± SEM of eight determinations per group and are expressed as percent of control. *p < 0.05; **p < 0.01 different from control; # p < 0.05, ## p < 0.01 different from 6-OHDA; § p < 0.05, §§ p < 0.01 different from 6-OHDA + UFP112 (ANOVA followed by Newman-Keuls)
SH-SY5Y cells were cultured in the presence of 30 μM 6-OHDA together with either 1 μM or 10 μM of N/OFQ for 24 h at which time an MTT assay was performed. While N/OFQ alone did not significantly reduce cell viability, there was a trend towards a reduction at the higher dose (Fig. 1b). Interestingly, we found that 1 μM or 10 μM N/OFQ significantly exacerbated the effects of 6-OHDA on cell viability (Fig. 1b). To confirm that this was mediated through the NOP receptor, a similar experiment was performed using the NOP agonist, UFP112 (3 μM) and cell viability was assessed using an MTT and LDH assay (Fig. 1c, d). We found using an MTT assay that UFP112 also exacerbated the effects of 6-OHDA on cell viability (Fig. 1c). This effect was prevented by the NOP antagonists UFP101 (10 μM) and SB-612111 (3 μM) (Fig. 1c). These findings were confirmed using a LDH assay (Fig. 1d). In agreement with the MTT assay (Fig. 1c), the NOP agonist, UFP112 combined with 6-OHDA resulted in significantly higher levels of LDH compared to 6-OHDA alone, an effect that was prevented by the NOP receptor antagonists (Fig. 1d). These data suggested that N/OFQ-NOP may affect DA neuronal survival. To investigate this further and to assess whether N/OFQ affected DA axons, we used primary cultures of the E14 rat VM.
N/OFQ Inhibits the Survival and Growth of Midbrain DA Neurons
To confirm our hypothesis that N/OFQ is detrimental to midbrain DA neurons, the effect of N/OFQ only on the survival and axonal growth of primary cultures of DA neurons was examined and compared to the effect of the neurotoxins, 6-OHDA and MPP+. To do this, a dose response was initially performed. E14 rat VM cultures were prepared and cultured for 24 h in the presence of increasing concentrations of 6-OHDA (0–30 μM), MPP+ (0–30 μM) and N/OFQ (0–500nM) and immunocytochemically stained for TH to visualize DA neurons and their axons. Treatment with 6-OHDA (Fig. 2a, b, g) and MPP+ (Fig. 2c, d, g) significantly reduced the numbers of DA neurons and those with processes when compared to vehicle-treated controls. Intriguingly, N/OFQ was found to be as effective as both DA neurotoxins in reducing the numbers of DA neurons and those with axons (Fig. 2e–g).

N/OFQ reduces the numbers of DA neurons in a dose-dependent manner. Primary cultures of E14 rat VM were incubated for 24 h with increasing concentrations of 6-OHDA (2–30 μM), MPP+ (2–30 μM) or N/OFQ (10–500nM) after which the numbers of DA neurons, and those with a visible neurite were assessed. a–f Graphical representations of the average number of TH-immunopositive DA neurons and those with intact neurites in a, b 6-OHDA, c, d MPP+ and e, f N/OFQ-treated E14 VM DA neurons at 24 h post-treatment. g Representative photomicrographs of E14 VM DA neurons immunocytochemically stained for TH. Scale bar = 50 μm. Data are expressed as mean ± SEM from three separate experiments. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. control; one-way ANOVA with post hoc Dunnett’s test
The effect of N/OFQ on the axonal growth of DA neurons was determined by analysing total neurite length and branching and by performing Sholl analysis, which provides a graphic illustration of neuronal complexity at defined distances from the cell body [50]. These effects of N/OFQ were next compared to those of 6-OHDA and MPP+. Treatment with increasing concentrations of 6-OHDA (Fig. 3a–c) and MPP+ (Fig. 3a, d, e) significantly reduced neurite length and neurite branching of DA neurons compared to vehicle-treated controls at the same time point. Similarly, it was found that increasing concentrations of N/OFQ also reduced neurite length and branching in a dose-dependent manner (Fig. 3a, f, g). These data were confirmed by the Sholl profiles which demonstrated that increasing concentrations of 6-OHDA (Fig. 4a, b), MPP+ (Fig. 4c, d) and N/OFQ (Fig. 4e, f) resulted in a significant decrease in the Sholl profiles of DA neurons, indicating a decrease in the complexity of the neurite field. These data support the hypothesis that N/OFQ exerts a direct neurotoxic and neuritotoxic effect on midbrain DA neurons.

6-OHDA, MPP+ and N/OFQ reduce the length and branching of VM dopaminergic neurons in a dose-dependent manner. a Representative photomicrographs of 6-OHDA, MPP+- and N/OFQ-treated E14 VM cultures immunocytochemically stained for TH. Scale bar = 50 μm. Graphical representations of the average neurite length and branching of b, c 6-OHDA, d, e MPP+ and f, g N/OFQ-treated DA neurons in E14 VM cultures. Data are expressed as mean ± SEM from three separate experiments. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. control; one-way ANOVA with post hoc Dunnett’s test

6-OHDA, MPP+ and N/OFQ reduce the complexity of VM DA neurons. Modified Sholl plots indicating the complexity of the neurite field of a, b 6-OHDA, c, d MPP+ and e, f N/OFQ-treated E14 VM DA neurons, as indicated. Data are expressed as mean ± SEM from three separate experiments. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. control; one-way ANOVA with post hoc Dunnett’s test
N/OFQ Potentiates the Effects of 6-OHDA and MPP+ on DA Neurons
It was next examined if N/OFQ could potentiate the effects of 6-OHDA and MPP+ on DA neurons. To do this, E14 rat VM cultures were cultured in sub-maximal concentrations of 6-OHDA (5 μM), MPP+ (5 μM), N/OFQ (100 nM) or a combination of N/OFQ and 6-OHDA or MPP+ for 24 h. Treatment with 6-OHDA, MPP+ and N/OFQ significantly reduced the numbers of DA neurons with processes when compared to vehicle-treated controls at the same time point (Fig. 5a, b). However, the effects of combined N/OFQ and MPP+ were significantly greater than either neurotoxin alone (Fig. 5a, b). Similarly, N/OFQ also exacerbated the effects of 6-OHDA and MPP+ on neurite length (Fig. 5c) and in the Sholl profiles which indicated that the effects of combined N/OFQ and either 6-OHDA or MPP+ on DA neuronal complexity were significantly greater than either 6-OHDA or MPP+ alone (Fig. 5d, e).

N/OFQ potentiates the neurotoxic effects of 6-OHDA and MPP+ in DA neurons. a Representative photomicrographs and b the numbers of DA neurons with processes and c DA neurite length as a percentage of control following treatment with 6-OHDA, MPP+, N/OFQ or a combination of N/OFQ and either toxin for 24 h. d, e Modified Sholl plots indicating the complexity of the neurite field of DA neurons comparing 6-OHDA to N/OFQ + 6-OHDA and MPP+ to MPP++N/OFQ. Data are expressed as mean ± SEM from three separate experiments. *p < 0.05, **p < 0.01, ****p < 0.0001 vs. control or as indicated. $$ p < 0.01 and $$$ p < 0.001 vs. 6-OHDA alone. ## p < 0.01 and ### p < 0.001 vs. MPP+ alone. ^ p < 0.05, and ^^ p <0.01 vs. N/OFQ alone. ANOVA followed by Newman-Keuls
N/OFQ Affects Midbrain DA Neurons Through a NOP-p38-MAPK Mechanism
The molecular mechanisms by which N/OFQ may exert its effects on midbrain DA neurons were subsequently examined. To determine if the effects of N/OFQ are mediated by the NOP receptor, a pharmacological inhibitor of the NOP receptor, SB612111 was utilized. A dose-response experiment was performed initially which established that treatment of primary cultures of the E14 rat VM with concentrations up to 10 μM SB612111 did not have any adverse effects on DA neurons (data not shown). To determine if the detrimental effects of N/OFQ on DA neurons were mediated by the NOP receptor, primary cultures of the E14 rat VM were pre-treated with 10 μM SB612111 for 1 h prior to treatment with 100 nM N/OFQ for 24 h. Pre-treatment with the NOP receptor antagonist protected DA neurons against the detrimental effects of N/OFQ on DA neurite length (Fig. 6a, b). This demonstrates that N/OFQ exerts its effects on DA neurons through the NOP receptor.

N/OFQ exerts its effects on DA neurons through a NOP-p38-MAPK pathway. a Graphical representation of average DA neuron length and b representative photomicrographs of E14 VM DA neurons following treatment with 100 nM N/OFQ in the presence or absence of 10 μM SB612111 (NOP receptor antagonist) for 24 h. c Graphical representations of c average neurite length and d neurite branching, e representative photomicrographs and f Sholl plots of E14 VM DA neurons treated with 100 nM N/OFQ in the presence or absence of 30 μM SB600125 (JNK inhibitor) or 30 μM SB203580 (p38 inhibitor) for 24 h. Data are expressed as mean ± SEM from three separate experiments. (***p < 0.001 vs. control; one-way ANOVA with Tukey’s test)
The intracellular mechanisms mediating the effects of N/OFQ on DA neurons may be through the modulation of a number of intracellular pathways. We hypothesized that p38 and/or JNK signalling may be involved as it has previously been demonstrated that these pathways affect DA neuron survival and furthermore that they are activated by N/OFQ, depending on the cellular context. Primary cultures of the E14 rat VM were pre-treated with the p-38 MAPK inhibitor, SB203580 or the JNK inhibitor, SP600125 (both at 30 μM; which does not affect DA neurons at this concentration) for 1 h prior to treatment with 100 nM N/OFQ for 24 h. Pre-treatment with SB203580, but not SP600125, protected DA neurons against the N/OFQ-mediated reductions in total neurite length (Fig. 6c), neurite branching (Fig. 6d, e) and in the Sholl profiles (Fig. 6f). Collectively, these data indicate that N/OFQ exerts its effects on DA neurons through a p38-MAPK-dependent mechanism.
Reduced N/OFQ and α-Synuclein mRNA Expression in the Human Midbrain in PD
We next examined N/OFQ and NOP receptor mRNA expression in the SN of PD patients and age-matched controls using microarray gene expression data. These data were obtained from three independent studies downloaded from Gene Expression Omnibus (GEO, series GSE20295 [52], GSE8397 [53] and GSE7621 [54]) (Table 1). Interestingly, we found a statistically significant down regulation of N/OFQ (Fig. 7a) but no change in the NOP receptor (Fig. 7b) in the PD SN compared to controls using three separate statistical tests RANK products [55], t test (parametric) and Wilcoxon test (non-parametric) (Fig. 7e). Interestingly, α-synuclein (SCNA) (Fig. 1c, e) also showed a strong down regulation in the PD SN (Fig. 7c, e), whereas in contrast, Tau (MAPT) mRNA levels were not different between controls and PD samples (Fig. 7d, e). These data provide the first evidence of alterations in N/OFQ expression in the human SN in PD that parallel those of α-synuclein.

Reduced expression of N/OFQ and a-synuclein in the human PD midbrain. a–d Gene expression comparisons for N/OFQ (PNOC), NOP (OPRL1), α-synuclein (SNCA) and Tau (MAPT) using microarray expression data derived from normal and Parkinson’s disease SN (see Table 1). e Statistical comparisons between control and disease-derived samples were carried using three separate approaches: RANK products, t test (parametric) and Wilcoxon test (non-parametric). Charts show the mean ± SEM for the indicated genes comparing all resulting control (n = 38) and PD samples (n = 51)
Discussion
In this study, the direct effects of the neuropeptide N/OFQ on the survival and growth of midbrain DA neurons were examined. While we have previously reported that N/OFQ contributes to motor impairment in PD [24], and that elevated levels of N/OFQ are found in the pre-clinical rodent models of PD as well as in patients with PD [24, 25], it was not known whether N/OFQ may contribute to the on-going degeneration of DA neurons, the pathological hallmark of the disorder. Since SNpc DA neurons express the NOP receptor [21, 22], we examined the effects of N/OFQ on survival and neurite growth in the SH-SY5Y neuroblastoma cell line, which has been used extensively as a model of human VM DA neurons [56, 57] and in primary cultures of the E14 rat VM, which is a widely used, experimentally tractable model for studying the physiological and pathological molecular mechanisms regulating these processes [42, 43, 58]. Here, we show that N/OFQ is as potent as the DA neurotoxins 6-OHDA and MPP+ in reducing DA neuron survival and growth. These findings suggest that elevations of N/OFQ observed in the Parkinsonian brain may play a causative role in degeneration of DA neurons and their processes. In agreement with this hypothesis, we found that N/OFQ exacerbated the detrimental effects of MPP+ and 6-OHDA on DA neuron survival and growth. This is supported by studies showing that deletion of the N/OFQ gene in mice conferred partial protection against MPTP-induced loss of SN dopamine neurons and against the loss of TH-positive DA nerve terminals in the caudate-putamen [24, 33]. These findings suggest that a perturbation in endogenous N/OFQ-NOP signalling may be involved in DA neuronal degeneration observed in PD. This suggestion is supported by the fact that NOP receptor antagonists reverse motor impairment in rodent [24, 26, 37, 59, 60] and non-human primate [26, 27] models of PD.
Interestingly, we also found that N/OFQ exerted a dose-dependent, detrimental effect on DA neuron survival and growth. Given that N/OFQ levels were found to be approximately 3.5-fold elevated in the CSF of Parkinsonian patients compared with non-parkinsonian neurologic controls [25], CSF profiling of endogenous N/OFQ levels may be a useful indicator of disease progression. This is supported by the finding that DA depletion was associated with increased mRNA N/OFQ levels in the SNpc, whereas no change was observed in globus pallidus, nucleus accumbens, thalamus and motor cortex [25], possibility indicating a specific relationship between N/OFQ and DA neurons.
In terms of molecular mechanisms, N/OFQ [38, 61] and opioids in general [39], not only act through the canonical Gi pathway but also through the MAP-kinase pathway most likely recruited by the βγ subunits [38]. We hypothesized that p38-MAPK-kinase signalling specifically may be involved in mediating the effects of N/OFQ on DA neurons given that it has been shown to be activated by N/OFQ [38, 61] and multiple lines of evidence implicate p38-MAPK signalling in the DA degeneration observed in PD. Increased staining of phospho-p38 has been shown in the SNpc (but not VTA) DA neurons in human brain sections from PD patients and in MPTP-treated mice [62]. Furthermore, the DA neurotoxins 6-OHDA and MPTP activate p38-MAPK in SNpc DA neurons [40, 62, 63], and the vulnerability of DA neurons to neurotoxic insult is dependent, at least in part, upon p38-MAPK signalling [62, 64–67]. Moreover, neurotrophic factors and kinases which promote DA neuronal survival and neurite growth dampen p38-MAPK signalling, suggesting that activation of p38-MAPK signalling may contribute to DA degeneration. When we examined the molecular mechanisms by which N/OFQ affected DA neurons we found that the neurotoxic and neuritoxic effects of N/OFQ are dependent on p38-MAPK signalling. Indeed, we found that the detrimental effect of N/OFQ on DA neurons is prevented by a p38 but not a JNK inhibitor, implying that the neurotoxic and neuritoxic effects of N/OFQ are dependent on p38-MAPK signalling. Given that SNpc DA neurons express the NOP receptor [21, 22] and that high levels of N/OFQ are found in the SNpc and are increased in the CSF of PD patients [24, 34] along with elevated phospho-p38 [62], these data suggest that elevations in N/OFQ in the parkinsonian brain may act directly on DA neurons through the NOP receptor thereby activating p38-MAPK signalling which leads to DA neuron degeneration.
Under this perspective, the reduction of N/OFQ mRNA observed in the human SNpc might be viewed as a compensatory mechanism to prevent excessive NOP receptor stimulation and protect residual DA neurons. These data do not easily reconcile with previous findings showing an increase in N/OFQ expression in 6-OHDA hemilesioned rats [24, 25, 68] or MPTP/MPP+-treated mice [68, 69], which correlate with the increase of N/OFQ levels monitored in the rat SNpr [24] and in the CSF of PD patients [25]. However, it is possible that the impact of acute (and selective) loss of SNpc DA neurons on N/OFQ transmission, as that produced by toxins in rodents, is different from that caused by chronic and progressive degeneration of DA and non-DA neurons in humans. Moreover, CSF samples processed in the Marti 2010 study [25] were taken from patients much earlier (mean age 60.6 ± 1.8), when compared to samples used in the Zhang et al. study (GSE20295 [52]; mean age 76.7 ± 6.2) and in the Moran et al. study (GSE8397 [53]; mean age: 80.0 ± 5.7). This suggests that PD progression might cause biphasic changes of N/OFQ expression in SN, reflecting plastic adaptations inside and outside the SN.
In summary, these results demonstrate for the first time that N/OFQ is detrimental to the survival and growth of DA neurons and support the theory that NOP receptor antagonists may represent a novel class of neuroprotective therapy for the protection of DA neurons and their neurite processes in PD.
References
Lees AJ, Hardy J, Revesz T (2009) Parkinson’s disease. Lancet 373(9680):2055–2066. doi:10.1016/S0140-6736(09)60492-X
de Lau LM, Breteler MM (2006) Epidemiology of Parkinson’s disease. Lancet Neurol 5(6):525–535. doi:10.1016/S1474-4422(06)70471-9
Jankovic J (2008) Parkinson’s disease: clinical features and diagnosis. J Neurol Neurosurg Psychiatry 79(4):368–376. doi:10.1136/jnnp.2007.131045
Bethlem J, Den Hartog Jager WA (1960) The incidence and characteristics of Lewy bodies in idiopathic paralysis agitans (Parkinson’s disease). J Neurol Neurosurg Psychiatry 23:74–80
Jellinger KA (1991) Pathology of Parkinson’s disease. Changes other than the nigrostriatal pathway. Mol Chem Neuropathol 14(3):153–197
Jellinger KA (2012) Neuropathology of sporadic Parkinson’s disease: evaluation and changes of concepts. Mov Disord 27(1):8–30. doi:10.1002/mds.23795
Fearnley JM, Lees AJ (1991) Ageing and Parkinson’s disease: substantia nigra regional selectivity. Brain 114(Pt 5):2283–2301
Ma SY, Roytta M, Rinne JO, Collan Y, Rinne UK (1997) Correlation between neuromorphometry in the substantia nigra and clinical features in Parkinson’s disease using disector counts. J Neurol Sci 151(1):83–87
Greffard S, Verny M, Bonnet AM, Beinis JY, Gallinari C, Meaume S, Piette F, Hauw JJ, Duyckaerts C (2006) Motor score of the Unified Parkinson Disease Rating Scale as a good predictor of Lewy body-associated neuronal loss in the substantia nigra. Arch Neurol 63(4):584–588. doi:10.1001/archneur.63.4.584
Lee CS, Samii A, Sossi V, Ruth TJ, Schulzer M, Holden JE, Wudel J, Pal PK, de la Fuente-Fernandez R, Calne DB, Stoessl AJ (2000) In vivo positron emission tomographic evidence for compensatory changes in presynaptic dopaminergic nerve terminals in Parkinson’s disease. Ann Neurol 47(4):493–503
Finkelstein DI, Stanic D, Parish CL, Tomas D, Dickson K, Horne MK (2000) Axonal sprouting following lesions of the rat substantia nigra. Neuroscience 97(1):99–112
Stanic D, Finkelstein DI, Bourke DW, Drago J, Horne MK (2003) Timecourse of striatal re-innervation following lesions of dopaminergic SNpc neurons of the rat. Eur J Neurosci 18(5):1175–1188
Bezard E, Gross CE, Brotchie JM (2003) Presymptomatic compensation in Parkinson’s disease is not dopamine-mediated. Trends Neurosci 26(4):215–221. doi:10.1016/S0166-2236(03)00038-9
Samii A, Nutt JG, Ransom BR (2004) Parkinson’s disease. Lancet 9423:1783–1793. doi:10.1016/S0140-6736(04)16305-8
Toulouse A, Sullivan AM (2008) Progress in Parkinson’s disease-where do we stand? Prog Neurobiol 85(4):376–392. doi:10.1016/j.pneurobio.2008.05.003
Meunier JC, Mollereau C, Toll L, Suaudeau C, Moisand C, Alvinerie P, Butour JL, Guillemot JC, Ferrara P, Monsarrat B et al (1995) Isolation and structure of the endogenous agonist of opioid receptor-like ORL1 receptor. Nature 377(6549):532–535. doi:10.1038/377532a0
Reinscheid RK, Nothacker HP, Bourson A, Ardati A, Henningsen RA, Bunzow JR, Grandy DK, Langen H, Monsma FJ Jr, Civelli O (1995) Orphanin FQ: a neuropeptide that activates an opioidlike G protein-coupled receptor. Science 270(5237):792–794
Calo G, Bigoni R, Rizzi A, Guerrini R, Salvadori S, Regoli D (2000) Nociceptin/orphanin FQ receptor ligands. Peptides 21(7):935–947
Mogil JS, Pasternak GW (2001) The molecular and behavioral pharmacology of the orphanin FQ/nociceptin peptide and receptor family. Pharmacol Rev 53(3):381–415
Mollereau C, Moisand C, Butour JL, Parmentier M, Meunier JC (1996) Replacement of Gln280 by His in TM6 of the human ORL1 receptor increases affinity but reduces intrinsic activity of opioids. FEBS Lett 395(1):17–21
Neal CR Jr, Mansour A, Reinscheid R, Nothacker HP, Civelli O, Akil H, Watson SJ Jr (1999) Opioid receptor-like (ORL1) receptor distribution in the rat central nervous system: comparison of ORL1 receptor mRNA expression with (125)I-[(14)Tyr]-orphanin FQ binding. J Comp Neurol 412(4):563–605
Neal CR Jr, Mansour A, Reinscheid R, Nothacker HP, Civelli O, Watson SJ Jr (1999) Localization of orphanin FQ (nociceptin) peptide and messenger RNA in the central nervous system of the rat. J Comp Neurol 406(4):503–547
Lambert DG (2008) The nociceptin/orphanin FQ receptor: a target with broad therapeutic potential. Nat Rev Drug Discov 7(8):694–710. doi:10.1038/nrd2572
Marti M, Mela F, Fantin M, Zucchini S, Brown JM, Witta J, Di Benedetto M, Buzas B, Reinscheid RK, Salvadori S, Guerrini R, Romualdi P, Candeletti S, Simonato M, Cox BM, Morari M (2005) Blockade of nociceptin/orphanin FQ transmission attenuates symptoms and neurodegeneration associated with Parkinson’s disease. J Neurosci 25(42):9591–9601. doi:10.1523/JNEUROSCI.2546-05.2005
Marti M, Sarubbo S, Latini F, Cavallo M, Eleopra R, Biguzzi S, Lettieri C, Conti C, Simonato M, Zucchini S, Quatrale R, Sensi M, Candeletti S, Romualdi P, Morari M (2010) Brain interstitial nociceptin/orphanin FQ levels are elevated in Parkinson’s disease. Mov Disord 25(11):1723–1732. doi:10.1002/mds.23271
Viaro R, Sanchez-Pernaute R, Marti M, Trapella C, Isacson O, Morari M (2008) Nociceptin/orphanin FQ receptor blockade attenuates MPTP-induced parkinsonism. Neurobiol Dis 30(3):430–438. doi:10.1016/j.nbd.2008.02.011
Visanji NP, de Bie RM, Johnston TH, McCreary AC, Brotchie JM, Fox SH (2008) The nociceptin/orphanin FQ (NOP) receptor antagonist J-113397 enhances the effects of levodopa in the MPTP-lesioned nonhuman primate model of Parkinson’s disease. Mov Disord 23(13):1922–1925. doi:10.1002/mds.22086
Ozawa A, Brunori G, Mercatelli D, Wu J, Cippitelli A, Zou B, Xie XS, Williams M, Zaveri NT, Low S, Scherrer G, Kieffer BL, Toll L (2015) Knock-in mice with NOP-eGFP receptors identify receptor cellular and regional localization. J Neurosci 35(33):11682–11693. doi:10.1523/JNEUROSCI.5122-14.2015
Maidment NT, Chen Y, Tan AM, Murphy NP, Leslie FM (2002) Rat ventral midbrain dopamine neurons express the orphanin FQ/nociceptin receptor ORL-1. Neuroreport 13(9):1137–1140
Norton CS, Neal CR, Kumar S, Akil H, Watson SJ (2002) Nociceptin/orphanin FQ and opioid receptor-like receptor mRNA expression in dopamine systems. J Comp Neurol 444(4):358–368
Acosta C, Davies A (2008) Bacterial lipopolysaccharide regulates nociceptin expression in sensory neurons. J Neurosci Res 86(5):1077–1086. doi:10.1002/jnr.21565
Buzas B, Symes AJ, Cox BM (1999) Regulation of nociceptin/orphanin FQ gene expression by neuropoietic cytokines and neurotrophic factors in neurons and astrocytes. J Neurochem 72(5):1882–1889
Brown JM, Gouty S, Iyer V, Rosenberger J, Cox BM (2006) Differential protection against MPTP or methamphetamine toxicity in dopamine neurons by deletion of ppN/OFQ expression. J Neurochem 98(2):495–505. doi:10.1111/j.1471-4159.2006.03902.x
Neal CR Jr, Akil H, Watson SJ Jr (2001) Expression of orphanin FQ and the opioid receptor-like (ORL1) receptor in the developing human and rat brain. J Chem Neuroanat 22(4):219–249
Marti M, Guerrini R, Beani L, Bianchi C, Morari M (2002) Nociceptin/orphanin FQ receptors modulate glutamate extracellular levels in the substantia nigra pars reticulata. A microdialysis study in the awake freely moving rat. Neuroscience 112(1):153–160
Mallimo EM, Kusnecov AW (2013) The role of orphanin FQ/nociceptin in neuroplasticity: relationship to stress, anxiety and neuroinflammation. Front Cell Neurosci 7:173. doi:10.3389/fncel.2013.00173
Marti M, Mela F, Veronesi C, Guerrini R, Salvadori S, Federici M, Mercuri NB, Rizzi A, Franchi G, Beani L, Bianchi C, Morari M (2004) Blockade of nociceptin/orphanin FQ receptor signaling in rat substantia nigra pars reticulata stimulates nigrostriatal dopaminergic transmission and motor behavior. J Neurosci 24(30):6659–6666. doi:10.1523/JNEUROSCI.0987-04.2004
New DC, Wong YH (2002) The ORL1 receptor: molecular pharmacology and signalling mechanisms. Neuro-Signals 11(4):197–212
Law PY, Wong YH, Loh HH (2000) Molecular mechanisms and regulation of opioid receptor signaling. Annu Rev Pharmacol Toxicol 40:389–430. doi:10.1146/annurev.pharmtox.40.1.389
Crotty S, Fitzgerald P, Tuohy E, Harris DM, Fisher A, Mandel A, Bolton AE, Sullivan AM, Nolan Y (2008) Neuroprotective effects of novel phosphatidylglycerol-based phospholipids in the 6-hydroxydopamine model of Parkinson’s disease. Eur J Neurosci 27(2):294–300. doi:10.1111/j.1460-9568.2007.06018.x
Presgraves SP, Ahmed T, Borwege S, Joyce JN (2004) Terminally differentiated SH-SY5Y cells provide a model system for studying neuroprotective effects of dopamine agonists. Neurotox Res 5(8):579–598
Collins LM, O’Keeffe GW, Long-Smith CM, Wyatt SL, Sullivan AM, Toulouse A, Nolan YM (2013) Mitogen-activated protein kinase phosphatase (MKP)-1 as a neuroprotective agent: promotion of the morphological development of midbrain dopaminergic neurons. Neruomol Med 15(2):435–446. doi:10.1007/s12017-013-8230-5
Hegarty SV, Collins LM, Gavin AM, Roche SL, Wyatt SL, Sullivan AM, O’Keeffe GW (2014) Canonical BMP-Smad signalling promotes neurite growth in rat midbrain dopaminergic neurons. Neruomol Med. doi:10.1007/s12017-014-8299-5
Cheng J, Standifer KM, Tublin PR, Su W, Pasternak GW (1995) Demonstration of kappa 3-opioid receptors in the SH-SY5Y human neuroblastoma cell line. J Neurochem 65(1):170–175
Connor M, Yeo A, Henderson G (1996) The effect of nociceptin on Ca2+ channel current and intracellular Ca2+ in the SH-SY5Y human neuroblastoma cell line. Br J Pharmacol 118(2):205–207
Rizzi A, Spagnolo B, Wainford RD, Fischetti C, Guerrini R, Marzola G, Baldisserotto A, Salvadori S, Regoli D, Kapusta DR, Calo G (2007) In vitro and in vivo studies on UFP-112, a novel potent and long lasting agonist selective for the nociceptin/orphanin FQ receptor. Peptides 28(6):1240–1251. doi:10.1016/j.peptides.2007.04.020
Calo G, Rizzi A, Rizzi D, Bigoni R, Guerrini R, Marzola G, Marti M, McDonald J, Morari M, Lambert DG, Salvadori S, Regoli D (2002) [Nphe1, Arg14, Lys15]nociceptin-NH2, a novel potent and selective antagonist of the nociceptin/orphanin FQ receptor. Br J Pharmacol 136(2):303–311. doi:10.1038/sj.bjp.0704706
Zaratin PF, Petrone G, Sbacchi M, Garnier M, Fossati C, Petrillo P, Ronzoni S, Giardina GA, Scheideler MA (2004) Modification of nociception and morphine tolerance by the selective opiate receptor-like orphan receptor antagonist (−)-cis-1-methyl-7-[[4-(2,6-dichlorophenyl)piperidin-1-yl]methyl]-6,7,8,9-tetrahy dro-5H-benzocyclohepten-5-ol (SB-612111). J Pharmacol Exp Ther 308(2):454–461. doi:10.1124/jpet.103.055848
Spagnolo B, Carra G, Fantin M, Fischetti C, Hebbes C, McDonald J, Barnes TA, Rizzi A, Trapella C, Fanton G, Morari M, Lambert DG, Regoli D, Calo G (2007) Pharmacological characterization of the nociceptin/orphanin FQ receptor antagonist SB-612111 [(−)-cis-1-methyl-7-[[4-(2,6-dichlorophenyl)piperidin-1-yl]methyl]-6,7,8,9-tetrah ydro-5H-benzocyclohepten-5-ol]: in vitro studies. J Pharmacol Exp Ther 321(3):961–967. doi:10.1124/jpet.106.116764
Gutierrez H, Davies AM (2007) A fast and accurate procedure for deriving the Sholl profile in quantitative studies of neuronal morphology. J Neurosci Methods 163(1):24–30. doi:10.1016/j.jneumeth.2007.02.002
O’Keeffe GW, Dockery P, Sullivan AM (2004) Effects of growth/differentiation factor 5 on the survival and morphology of embryonic rat midbrain dopaminergic neurones in vitro. J Neurocytol 33(5):479–488
Zhang Y, James M, Middleton FA, Davis RL (2005) Transcriptional analysis of multiple brain regions in Parkinson’s disease supports the involvement of specific protein processing, energy metabolism, and signaling pathways, and suggests novel disease mechanisms. Am J Med GenPart B Neuropsychiatr Genet 137B(1):5–16. doi:10.1002/ajmg.b.30195
Moran LB, Duke DC, Deprez M, Dexter DT, Pearce RK, Graeber MB (2006) Whole genome expression profiling of the medial and lateral substantia nigra in Parkinson’s disease. Neurogenetics 7(1):1–11. doi:10.1007/s10048-005-0020-2
Lesnick TG, Papapetropoulos S, Mash DC, Ffrench-Mullen J, Shehadeh L, de Andrade M, Henley JR, Rocca WA, Ahlskog JE, Maraganore DM (2007) A genomic pathway approach to a complex disease: axon guidance and Parkinson disease. PLoS Genet 3(6), e98. doi:10.1371/journal.pgen.0030098
Hong F, Breitling R, McEntee CW, Wittner BS, Nemhauser JL, Chory J (2006) RankProd: a bioconductor package for detecting differentially expressed genes in meta-analysis. Bioinformatics 22(22):2825–2827. doi:10.1093/bioinformatics/btl476
Hegarty SV, Sullivan AM, O’Keeffe GW (2013) BMP2 and GDF5 induce neuronal differentiation through a Smad dependant pathway in a model of human midbrain dopaminergic neurons. Mol Cell Neurosci 56:263–271. doi:10.1016/j.mcn.2013.06.006
Xie HR, Hu LS, Li GY (2010) SH-SY5Y human neuroblastoma cell line: in vitro cell model of dopaminergic neurons in Parkinson’s disease. Chin Med J (Engl) 123(8):1086–1092
Collins LM, Adriaanse LJ, Theratile SD, Hegarty SV, Sullivan AM, O’Keeffe GW (2014) Class-IIa histone deacetylase inhibition promotes the growth of neural processes and protects them against neurotoxic insult. Mol Neurobiol. doi:10.1007/s12035-014-8820-8
Marti M, Trapella C, Morari M (2008) The novel nociceptin/orphanin FQ receptor antagonist Trap-101 alleviates experimental parkinsonism through inhibition of the nigro-thalamic pathway: positive interaction with L-DOPA. J Neurochem 107(6):1683–1696. doi:10.1111/j.1471-4159.2008.05735.x
Volta M, Mabrouk OS, Bido S, Marti M, Morari M (2010) Further evidence for an involvement of nociceptin/orphanin FQ in the pathophysiology of Parkinson’s disease: a behavioral and neurochemical study in reserpinized mice. J Neurochem 115(6):1543–1555. doi:10.1111/j.1471-4159.2010.07061.x
Armstead WM (2006) Differential activation of ERK, p38, and JNK MAPK by nociceptin/orphanin FQ in the potentiation of prostaglandin cerebrovasoconstriction after brain injury. Eur J Pharmacol 529(1–3):129–135. doi:10.1016/j.ejphar.2005.08.059
Karunakaran S, Saeed U, Mishra M, Valli RK, Joshi SD, Meka DP, Seth P, Ravindranath V (2008) Selective activation of p38 mitogen-activated protein kinase in dopaminergic neurons of substantia nigra leads to nuclear translocation of p53 in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated mice. J Neurosci 28(47):12500–12509. doi:10.1523/JNEUROSCI.4511-08.2008
Hu X, Weng Z, Chu CT, Zhang L, Cao G, Gao Y, Signore A, Zhu J, Hastings T, Greenamyre JT, Chen J (2011) Peroxiredoxin-2 protects against 6-hydroxydopamine-induced dopaminergic neurodegeneration via attenuation of the apoptosis signal-regulating kinase (ASK1) signaling cascade. J Neurosci 31(1):247–261. doi:10.1523/JNEUROSCI.4589-10.2011
Izumi Y, Yamamoto N, Matsuo T, Wakita S, Takeuchi H, Kume T, Katsuki H, Sawada H, Akaike A (2009) Vulnerability to glutamate toxicity of dopaminergic neurons is dependent on endogenous dopamine and MAPK activation. J Neurochem 110(2):745–755. doi:10.1111/j.1471-4159.2009.06178.x
Choi WS, Eom DS, Han BS, Kim WK, Han BH, Choi EJ, Oh TH, Markelonis GJ, Cho JW, Oh YJ (2004) Phosphorylation of p38 MAPK induced by oxidative stress is linked to activation of both caspase-8-and-9-mediated apoptotic pathways in dopaminergic neurons. J Biol Chem 279(19):20451–20460. doi:10.1074/jbc.M311164200
Jeohn GH, Cooper CL, Wilson B, Chang RC, Jang KJ, Kim HC, Liu B, Hong JS (2002) p38 MAP kinase is involved in lipopolysaccharide-induced dopaminergic neuronal cell death in rat mesencephalic neuron-glia cultures. Ann N Y Acad Sci 962:332–346
Gomez-Lazaro M, Galindo MF, Concannon CG, Segura MF, Fernandez-Gomez FJ, Llecha N, Comella JX, Prehn JH, Jordan J (2008) 6-Hydroxydopamine activates the mitochondrial apoptosis pathway through p38 MAPK-mediated, p53-independent activation of Bax and PUMA. J Neurochem 104(6):1599–1612. doi:10.1111/j.1471-4159.2007.05115.x
Di Benedetto M, Cavina C, D’Addario C, Leoni G, Candeletti S, Cox BM, Romualdi P (2009) Alterations of N/OFQ and NOP receptor gene expression in the substantia nigra and caudate putamen of MPP+ and 6-OHDA lesioned rats. Neuropharmacology 56(4):761–767. doi:10.1016/j.neuropharm.2008.12.009
Gouty S, Brown JM, Rosenberger J, Cox BM (2010) MPTP treatment increases expression of pre-pro-nociceptin/orphanin FQ mRNA in a subset of substantia nigra reticulata neurons. Neuroscience 169(1):269–278. doi:10.1016/j.neuroscience.2010.04.033
Acknowledgements
The authors acknowledge Dr. Sean Crampton for the technical assistance. This work was supported by grant support from Science Foundation Ireland (Grant No. :10/RFP/NES2786) (GO’K).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Collins, L.M., Dal Bo, G., Calcagno, M. et al. Nociceptin/Orphanin FQ Inhibits the Survival and Axon Growth of Midbrain Dopaminergic Neurons Through a p38-MAPK Dependent Mechanism. Mol Neurobiol 53, 7284–7297 (2016). https://doi.org/10.1007/s12035-015-9611-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-015-9611-6