
Abstract
The accumulation of misfolded α-synuclein in dopamine (DA) neurons is believed to be of major importance in the pathogenesis of Parkinson’s disease (PD). Animal models of PD, based on viral-vector-mediated over-expression of α-synuclein, have been developed and show evidence of dopaminergic toxicity, providing us a good tool to investigate potential therapies to interfere with α-synuclein-mediated pathology. An efficient disease-modifying therapeutic molecule should be able to interfere with the neurotoxicity of α-synuclein aggregation. Our study highlighted the ability of an autophagy enhancer, trehalose (at concentrations of 5 and 2 % in drinking water), to protect against A53T α-synuclein-mediated DA degeneration in an adeno-associated virus serotype 1/2 (AAV1/2)-based rat model of PD. Behavioral tests and neurochemical analysis demonstrated a significant attenuation in α-synuclein-mediated deficits in motor asymmetry and DA neurodegeneration including impaired DA neuronal survival and DA turnover, as well as α-synuclein accumulation and aggregation in the nigrostriatal system by commencing 5 and 2 % trehalose at the same time as delivery of AAV. Trehalose (0.5 %) was ineffective on the above behavioral and neurochemical deficits. Further investigation showed that trehalose enhanced autophagy in the striatum by increasing formation of LC3-II. This study supports the concept of using trehalose as a novel therapeutic strategy that might prevent/reverse α-synuclein aggregation for the treatment of PD.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Parkinson’s disease (PD) is a progressive neurodegenerative disorder characterized by loss of dopamine (DA) neurons and accumulation of cytoplasmic Lewy bodies (LBs) in the substantia nigra (SN) [1, 2]. LBs are predominantly composed of abnormal misfolded α-synuclein [3, 4]. Increased levels of α-synuclein mRNA are found in the midbrain of patients with sporadic PD [5], and duplication or triplication of the α-synuclein gene is found in some forms of familial PD [6]. Missense mutations in the α-synuclein gene including A53T, A30P, and E46K are defined cause of familial PD. These findings suggest that abnormally high expression of α-synuclein contributes to LB formation in PD brains [7, 8]. Therapeutic interventions targeting abnormal α-synuclein aggregation might be promising in preventing or slowing down the degeneration process of PD.
Autophagy is the only known mechanism that eukaryotic cells possess to degrade protein. Recent studies have described trehalose as having an ability to stabilize protein folding and induce autophagy to clear abnormal protein aggregates [9–12]. Trehalose, a natural disaccharide found in invertebrates, fungi, and many plants, functions to protect the integrity of the cells against various environmental stresses [13]. It can be administered safely and orally and has been accepted as a safe food ingredient by the European regulatory authorities following designation, as generally regarded as safe (GRAS) by the Food and Drug Administration in the USA [14]. Orally administered trehalose is absorbed in a similar fashion to glucose, although it is at least in part hydrolyzed to glucose monomers by the enzyme trehalase located on the brush border of the intestinal mucosa [15]. Trehalose does not readily cross cell membranes but can be efficiently loaded into mammalian cells through endocytosis and pinocytosis, exerting a variety of effects such as anti-aggregation and autophagy enhancement [16, 17].
Viral vector approaches have been used to model alpha-synucleinopathy in PD by driving α-synuclein over-expression in the basal ganglia directly through injection into the SN [18]. This approach produces progressive nigrostriatal degeneration along with motor impairments. In a previous study, we developed a new viral-vector-based model of PD that could consistently deliver nigrostriatal pathology and relevant behavioral deficits in a relatively short period of time by using a formulation of chimeric adeno-associated virus 1/2 (AAV1/2) human A53T α-synuclein at a titer of 1.76*1012 gp/ml. This model shows a progressive degeneration along with a behavioral impairment that occurs over the course of 6 weeks. We found that over-expression of A53T α-synuclein produced insoluble aggregates and DA degeneration [19, 20]. In the present study, we carried out a set of in vivo experiments to assess whether trehalose had a beneficial effect by clearing abnormal A53T α-synuclein in a rat model of PD induced by direct injection of AAV1/2 human A53T α-synuclein into the pars compacta of the SN (SNpc).
Results
Trehalose Attenuated AAV1/2 A53T α-Synuclein-Induced Behavioral Impairment in Rats
As shown in Fig. 1, animals that received unilateral AAV1/2 A53T α-synuclein showed a deficit in the use of their paw contralateral to the side of vector injection 3 weeks post-surgery compared with empty vector controls. This was represented by a 36.42 % increase forelimb use asymmetry in AAV1/2 A53T α-synuclein-injected rats, compared with empty vector controls (P < 0.01). Oral administration of 5 and 2 % trehalose provided a 29.1 and 39.85 % improvement in forelimb use asymmetry (P < 0.01) while 0.5 % trehalose and 5 % glucose had no significant effect in AAV1/2 A53T α-synuclein-injected rats (P > 0.05). Six weeks following vector injection, animals injected with AAV1/2 A53T α-synuclein continued to show forelimb use asymmetry (44.07 %) compared to empty vector/vehicle controls (P < 0.01). Oral administration of 5 and 2 % trehalose maintained an improvement with a 32 and 40 % of the forelimb use asymmetry compared to empty vector/vehicle controls (P < 0.01) while 0.5 % trehalose and 5 % glucose were ineffective (both, P > 0.05).

Forelimb asymmetry in the cylinder test. Three weeks and 6 weeks following injection of AAV1/2 A53T α-synuclein and oral administration of trehalose and glucose, the asymmetry in forelimb use was observed in rats. Bars indicate SD for seven to ten rats at each point. *P < 0.05, **P < 0.01, ***P < 0.001 compared with the empty vector controls; ## P < 0.01, ### P < 0.001 compared with the AAV1/2 A53T α-synuclein-injected rats that received either no treatment or 5 % glucose
Trehalose and Glucose did not Affect the Blood Sugar in Rats
Since trehalose and glucose are both saccharides, and trehalose can be metabolized to produce glucose, we measured blood sugar in the rats. Six weeks after drinking 5, 2, 0.5 % trehalose and 5 % glucose, we measured the free blood sugar and random blood sugar in each group. We did not find any significance between empty vector controls and AAV1/2 A53T α-synuclein groups with or without oral trehalose and glucose administration (Fig. 2, P > 0.05).

Effects of trehalose on the blood glucose in AAV1/2 A53T α-synuclein-injected rats. Six weeks following injection of AAV1/2 A53T α-synuclein and trehalose and glucose treatment, the free and random blood glucose was monitored. Bars indicate SD for seven to ten rats at each point
Trehalose Improved DA Neuronal Survival and Inhibited α-Synuclein Accumulation Produced by Nigral Delivery of AAV1/2 A53T α-Synuclein in Rats
The characteristic pathological event of PD is the loss of DA neurons in SN. In this study, we also observed the DA neuronal loss with nigral delivery of AAV1/2 A53T α-synuclein. The data showed the rats developed a significant loss of tyrosine-hydroxylase-immunoreactive (TH-ir) neurons in the right SN of rats 6 weeks following delivery of vectors coding for human A53T α-synuclein compared with that of empty vector controls as well as contralateral controls (Fig. 3a). TH-ir neurons were counted stereologically from anterior to posterior of the SN and showed a 38 % reduction in the right SN of AAV1/2 A53T α-synuclein-injected rats compared with empty vector controls (Fig. 3c, P < 0.001). Administration of 5 and 2 % trehalose protected DA neurons in SN, while neither 0.5 % trehalose nor 5 % glucose had any protective effects on DA neurons in SN compared with solely AAV1/2 A53T α-synuclein-injected rats (Fig. 3a). However, 5 and 2 % trehalose administration made a 23 and 32 % recovery of DA neuronal survival in SN, respectively, while 0.5 % trehalose and 5 % glucose did not have any effects on DA neuronal survival in SN compared with AAV1/2 A53T α-synuclein/vehicle-treated rats.

Effects of trehalose on DA neuronal survival and α-synuclein deposition in SN in AAV1/2 A53T α-synuclein-injected rats. a Immunofluorescence staining showed DA neurons (green) and α-synuclein accumulation (red) in SN of rats challenged with right intranigral injection of AAV1/2 A53T α-synuclein and trehalose administration for 6 weeks. Scale bar is 500 μm. b The red point represents the injection site of AAV1/2 A53T α-synuclein in the SN of rats. c Stereological counting of TH-ir neurons in SN. *P < 0.05, **P < 0.01, ***P < 0.001 compared with the empty vector controls; # P < 0.05, ## P < 0.01 compared with the AAV1/2 A53T α-synuclein-injected rats that received either no treatment or 5 % glucose (Color figure online)
Deposition and aggregation of α-synuclein in LBs are also a pathological hallmark of PD [23]. Figure 3a shows, using double-labeled immunofluorescence, co-localization of TH-ir neurons (green) and α-synuclein expression (red). As shown in Fig. 3a, the expression of α-synuclein in the AAV1/2 A53T α-synuclein-injected SN was obviously enhanced compared with empty vector controls. However, 5 and 2 % trehalose treatment cleared or prevented α-synuclein accumulation compared with AAV1/2 A53T α-synuclein/vehicle-treated rats. Neither 0.5 % trehalose nor 5 % glucose treatment had any obvious effects on α-synuclein expression. This qualitative observation warranted further investigation into the α-synuclein clearing potential of trehalose.
Trehalose Attenuated AAV1/2 A53T α-Synuclein-Reduced TH Expression and α-Synuclein Deposition in Striatum in Rats
Next, we examined the intensity of TH-ir and α-synuclein-ir in the striatum to determine whether dopamine terminals were also affected by treatment with trehalose (Fig. 4a). Similar to results shown in the SN, we found that the intensity of striatal TH-ir staining was reduced by over-expression of A53T α-synuclein in vehicle-treated animals, while animals that received 5 or 2 % trehalose appeared to have TH levels closer to that of empty vector/vehicle controls. To quantify this finding, we conducted optical density analysis (Fig. 4b) and show that TH expression in A53T α-synuclein/vehicle-treated animals is reduced by 32.7 % compared to empty vector/vehicle controls (P < 0.05). However, in rats in groups that received 5 and 2 % trehalose, there was significant protection of TH protein expression, by 22.8 and 24.3 % compared to A53T α-synuclein/vehicle-treated animals, respectively (Fig. 4b, P < 0.05). Furthermore, treatment with 5 and 2 % trehalose significantly reduced the amount of striatal α-synuclein, by 78.9 and 69.3 % compared to A53T α-synuclein/vehicle-treated animals, respectively (Fig. 4c, P < 0.05). Treatment with 0.5 % trehalose or 5 % glucose had no detectable effect on TH-ir staining and α-synuclein expression.

Effects of trehalose on striatal TH and α-synuclein expression in AAV1/2 A53T α-synuclein-injected rats. a Immunofluorescence staining showed TH expression (green) and α-synuclein accumulation (red) in striatum of rats challenged with right intranigral injection of AAV1/2 A53T α-synuclein and trehalose treatment for 6 weeks. Scale bar is 2 mm and 500 μm. b Representative photomicrographs illustrating TH immunoreactivity in striatum. *P < 0.05 compared with the empty vector controls; # P < 0.05 compared with the AAV1/2 A53T α-synuclein-injected rats that received either no treatment or 5 % glucose. c Western blot analyses of TH and α-synuclein in protein extracts from the right sides of the striatum in rats challenged with right intranigral injection of AAV1/2 A53T α-synuclein and trehalose treatment for 6 weeks. Semi-quantification of relative TH and α-synuclein protein was obtained from among at least six different animals. *P < 0.05, **P < 0.01, ***P < 0.001 compared with the empty vector controls; # P < 0.05, ## P < 0.01 compared with the AAV1/2 A53T α-synuclein-injected rats that received either no treatment or 5 % glucose (Color figure online)
Trehalose Treatment Prevents Neurochemical Deficits in Striatum of AAV1/2 A53T α-Synuclein-Injected Rats
To explore whether nigral delivery of AAV1/2 A53T α-synuclein has effects on the neurotransmitters, the levels of DA and its major metabolites dihydroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA) were determined by HPLC 6 weeks after AAV1/2 A53T α-synuclein injection and treatments. The concentrations of DA and DOPAC detected in striatum of AAV1/2 A53T α-synuclein-injected rats were significantly lower than empty vector controls by 53 and 32.7 %, respectively (Fig. 5a, b, P < 0.05), but the levels of HVA were not altered in AAV1/2 A53T α-synuclein-challenged rats. However, DA turnover (HVA/DA) in striatum was significantly increased (2.25-fold higher) in AAV1/2 A53T α-synuclein-injected rats compared with empty vector controls (Fig. 5c, P < 0.05, respectively), suggesting that the rate of DA metabolism was rapid in AAV1/2 A53T α-synuclein-injected rats.

Effects of trehalose on striatal changes in neurochemistry following intranigral injection of AAV1/2 A53T α-synuclein. The levels of DA and its major metabolites DOPAC and HVA in striatum were determined by specific HPLC assay 6 weeks post-AAV1/2 A53T α-synuclein injection and trehalose treatment. a DA; b DOPAC; c the ration of HVA/DA; d the ration of DOPAC/DA. Quantitative results were obtained from among at least six rats. *P < 0.05, **P < 0.01 compared with the empty controls; # P < 0.05, ## P < 0.01 compared with the AAV1/2 A53T α-synuclein-injected rats that received either no treatment or 5 % glucose
The decreased concentration of DA by AAV1/2 A53T α-synuclein injection was alleviated by 26, 42.5, and 55.7 % after oral 5, 2, and 0.5 % trehalose administration separately (Fig. 5a, P > 0.05 for 5 % trehalose, P < 0.05 for 2 and 0.5 % trehalose, respectively). The decreased concentration of DOPAC was alleviated by 27.5 % after 5 % trehalose treatment (Fig. 5b, P < 0.05 for 5 % trehalose). 2 % trehalose treatment seemed to have no effect on DOPAC levels. However, DA turnover (HVA/DA) in striatum of AAV1/2 A53T α-synuclein-challenged rats was greatly reversed by 64, 57.4, and 60.8 % post-oral 5, 2, and 0.5 % trehalose administration (Fig. 5c, P < 0.05 for 5, 2, and 0.5 % trehalose, respectively). Glucose of 5 % had no influence on DA and DOPAC release as well as DA turnover.
Trehalose Cleared the Insoluble α-Synuclein Deposited in the SN in AAV1/2 A53T α-Synuclein-Injected Rats
Considering that insoluble aggregation of α-synuclein is potentially pathogenic, we examined proteinase K (PK)-resistant α-synuclein immunostaining in rats challenged with AAV1/2 A53T α-synuclein. PK-resistant, Lewy-body-like α-synuclein aggregates were observed in the perinuclear compartment of DA neurons in AAV1/2 A53T α-synuclein-injected right SN compared with empty vector controls, indicating that insoluble α-synuclein may participate in the pathogenesis of DA degeneration in AAV1/2 A53T α-synuclein-injected rats. Treatment of 5 and 2 % trehalose effectively cleared α-synuclein aggregates compared with vehicle-treated AAV1/2 A53T α-synuclein-injected rats, while 0.5 % trehalose treatment was ineffective on this measure. Similarly, 5 % glucose had no effect on α-synuclein clearance (Fig. 6).

Effects of trehalose on clearance of α-synuclein aggregates in SN in AAV1/2 A53T α-synuclein-injected rats. Midbrain sections from rats injected with AAV1/2 A53T α-synuclein were treated with proteinase K and subsequently immunostained for human α-synuclein to determine its solubility. Following PK digestion, many neurons maintain expression of α-synuclein aggregates (insoluble α-synuclein). Scale bar is 500 μm
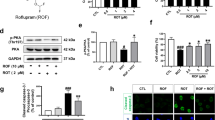
Trehalose Enhanced Autophagy in Striatum of AAV1/2 A53T α-Synuclein-Injected Rats
LC3 localizes to autophagic membranes but not on other membrane structures and thus serves as a specific marker for autophagosomes [21]. Endogenous LC3 is processed post-translationally into LC3-I, which is cytosolic. LC3-I is, in turn, converted to LC3-II, which associates with autophagosome membranes. LC3-II can accumulate due to increased autophagosome formation [22–25]. LC3-II levels relative to GAPDH correlate with autophagosome number per cell. As we are interested in defining a measure of autophagosome number per cell, we have not quantified LC3-II versus LC3-I, as some LC3-II can be converted back to LC3-I [16]. We observed a 3.6- and 3.8-fold increase of LC3-II in striatum after 5 and 2 % oral administration of trehalose for 6 weeks compared with solely AAV1/2 A53T α-synuclein-challenged rats (Fig. 7, P < 0.05 for 5 and 2 % trehalose, respectively). A 1.4-fold increase was also observed in 0.5 % trehalose-administrated rats, but it did not reach significance (P > 0.05). No detectable LC3-II changes were seen in 5 % glucose-administrated rats (P > 0.05).

Effects of trehalose on the LC3-II expression in striatum of AAV1/2 A53T α-synuclein-injected rats. The expression of LC3-II was assessed by Western blotting. Quantitative results were obtained from among at least six rats. *P < 0.05 compared with the AAV1/2 A53T α-synuclein-injected rats that received either no treatment or 5 % glucose
Discussion
Several neurodegenerative disorders are associated with intracellular, misfolded, aggregate-prone proteins, including β-amyloid in Alzheimer’s disease (AD), α-synuclein in PD, prion protein in prion disease, and huntingtin in Huntington’s disease (HD) [22, 25–27]. Novel strategies for preventing or slowing down the progression of these diseases are urgently needed. Trehalose is an autophagy enhancer acting through direct protein-trehalose interactions [13, 22]. Tanaka et al. found that trehalose was effective in alleviating polyglutamine-induced pathology by clearing mutant huntingtin and inhibited protein misfolding in a mouse model of HD [28]. Schaeffer et al. showed that trehalose could activate autophagy to reduce the number of neurons containing tau inclusions and improve neuronal survival in a mutant P301S tau mouse model of AD [29]. In the PD field, several in vitro studies have reported that trehalose could be neuroprotectant, by inhibiting the misfolding of α-synuclein. Sarkar et al. showed that trehalose accelerated the clearance of both A53T and A30P α-synuclein mutants in mammalian cell culture lines via stimulation of autophagy [22]. Our previous work reported that trehalose was effective in disaggregating mutant α-synuclein protofibrils and inhibiting the formation of mutant α-synuclein fibrils in the cell-free system of A53T α-synuclein incubation [30]. Further research confirmed that trehalose could enhance clearance of α-synuclein aggregates through activation of the autophagy pathway [31]. Recently, it has been argued that, given advances in animal models of PD, in vivo assessment of trehalose efficacy in PD is needed [20]. Accordingly, a recent in vivo study showed that trehalose attenuated neurodegeneration in a chronic MPTP mouse model of PD [32]. However, α-synuclein aggregates are scarce in MPTP mice, making it difficult to study the effects of trehalose on α-synuclein pathology and contribution in these mice.
Over-expression of α-synuclein in vivo using viral vectors has opened interesting possibilities to model PD-like synucleinopathy in rodents. In a previous study, we showed that AAV1/2-driven expression of A53T α-synuclein in the SN of rats caused a specific and progressive degeneration of the nigrostriatal system and a sustained behavioral deficit. The model has provided a useful tool to investigate new therapeutics directly targeting α-synuclein and its associated pathology [19, 20]. We have also shown that neuroinflammation (activated microglia, increases in IL-1β and TNF-α) is associated with this model as shown in a previous report [33] and in unpublished studies using the AAV1/2 vector (Koprich et al). The present study aimed to further investigate the potential protective effects of trehalose on motor function and α-synuclein-mediated pathology in this model. Our data showed that trehalose could significantly improve the motor deficits, DA neuronal survival, and α-synuclein deposition in SN and attenuated α-synuclein-induced lesion of dopaminergic terminals in the striatum. Furthermore, proteinase K digestion experiments revealed that trehalose reduced the amount of indigestible nigral α-synuclein. The inhibition of α-synuclein aggregation by trehalose can lead to amelioration of α-synuclein-mediated dysfunction.
The mechanism of action of trehalose in clearing α-synuclein and protecting the nigrostriatal pathway is not completely understood, but it is believed to activate autophagy in an mTOR-independent manner [22]. Our data implicated that trehalose activated autophagy in the striatum, as shown by the increased formation of LC3-II and reduced levels of α-synuclein. Altogether, our results indicate that the in vivo stimulation of autophagy markedly reduced the amount of α-synuclein aggregates, improved DA neuronal survival, and prevented behavioral asymmetry.
A possible adverse effect of trehalose treatment, elevation of blood sugar levels, was examined. We found that trehalose did not affect the fasting and random-sampled blood glucose in rats. In our data, 5 and 2 % trehalose, but not 5 % glucose, could effectively prevent α-synuclein-mediated degeneration of the nigrostriatal system. Trehalose of 0.5 % had no an effect on protecting striatal DA levels, nor was this concentration able to clear α-synuclein aggregates (control levels of PK-treated aggregates remained) or protect DA neurons.
Trehalose is currently being considered for a broad spectrum of applications. It has also been suggested to have anti-inflammatory, anti-oxidative stress, and anti-apoptotic effects [34–36]. These roles, coupled with anti-aggregation properties and lack of toxicity, enhance the potential translational value of trehalose. Extracting trehalose was once a difficult and costly process, but since 2000, an inexpensive mass production technology for extracting trehalose from starch has been available from the Hayashibara Company (Okayama, Japan).
Conclusions
In this study, we show beneficial effects of trehalose on the clearance of α-synuclein aggregation, the protection of nigro-striatal degeneration, the enhancement of autophagy, and prevention of behavioral deficits in an AAV1/2 A53T α-synuclein rat model of PD. Although further investigation is needed to define clinical design strategies and clarify the relationship between autophagy and neuroprotective effect of trehalose, the demonstrated benefit of trehalose in an AAV-driven PD model makes it a promising strategy for the prevention and treatment of α-synuclein-mediated PD disorders.
Methods
Animals and Vector Delivery
Female Sprague-Dawley (SD) rats, 2 months old and ~280 g, were purchased from Shanghai SLAC Laboratory Animal Company (Shanghai, China). All experiments were conducted in accordance with the guidelines of the International Council for Laboratory Animal Science. The study was approved by the Ethics Committee of Fudan University, Shanghai, China. The approval number from IRB is “20150649C001.” Using standard stereotaxic procedures, 60 SD rats were injected intranigrally with either empty AAV1/2 vector, or AAV1/2 expressing human A53T α-synuclein, under chloral hydrate anesthesia. In each case, a single 2-μl injection of viral vector was delivered to the SNpc at a rate of 0.2 μl/min according to the following coordinates from bregma: AP, −5.2 mm; ML, −2.0 mm; and DV, −7.5 mm (from skull at bregma) using a microinjector (Stoelting, Kiel, WI) and according to the atlas of Paxinos and Watson [20]. The AAV1/2 concentration of each vector was 1.76*1012 gp/ml (1:3 dilution). Animals were housed in pairs in a temperature-controlled environment (20 °C), kept on regular 12-h light/dark cycle, and allowed food and water ad libitum.
Vectors
Adeno-associated virus (AAV) vectors of a 1/2 serotype were designed such that expression was driven by a chicken beta actin (CBA) promoter hybridized with the cytomegalovirus (CMV) immediate early enhancer sequence. In addition, a woodchuck post-transcriptional regulatory element (WPRE) and a bovine growth hormone polyadenylation sequence (bGH-polyA) were incorporated to further enhance transcription following transduction. AAV1/2 is a chimeric vector where the capsid expresses AAV1 and AAV2 serotype proteins in a 1:1 ratio and uses the inverted terminal repeats (ITRs) from AAV2 according to the following scheme: CMV/CBA promoter––human A53T α-synuclein–WPRE-bGH-polyA––ITR [36]. The vectors were produced by GeneDetect Ltd., Auckland, New Zealand. Viral titers were determined by quantitative PCR (Applied Biosystems 7900 QPCR) with primers directed to the WPRE region, thus representing the number of functional physical particles of AAV in the solution containing the genome to be delivered. Full details of vector design can be found in the previous report [20].
Trehalose Treatment
After the surgery, the rats were treated with drinking water containing trehalose (Sinopharm Chemical Reagent Co., China) diluted in water with the final concentrations being 5, 2, and 0.5 %, respectively, for 6 weeks. Since glucose is the metabolite of trehalose, it was used as a control with a concentration being 5 % and also provided for 6 weeks.
Cylinder Test
Three and six weeks following AAV1/2 injection, spontaneous forepaw use was evaluated using the cylinder test. Briefly, rats with their right paws marked black were placed into a glass cylinder in front of two mirrors and they were video-recorded. Videos were scored post hoc by an observer blinded to the treatment conditions. For each rearing by the animal, it was recorded whether it used its left or right forepaw to touch the inner surface of the glass cylinder. Scoring was conducted over a period of 6 min. A minimum of 20 total touches was considered acceptable for inclusion in the analysis. The final data are presented as percent asymmetry (disparity between left [contralateral to the injection] and right paw use [ipsilateral to the lesion]), which was derived from the following equation: [(% right paw use) − (% left paw use) / (% right paw use) + (% left paw use)] * 100 [19].
Post-Mortem Measures
Immunohistochemistry
Six weeks after AAV1/2 injection, animals were administered an overdose of pentobarbital (1.0 ml of 240 mg/ml, i.p.) and killed by exsanguination by transcardial perfusion with saline followed by 4 % paraformaldehyde. Brains were then removed and processed for immuno-labeling. Brains were sectioned frozen in the coronal plane at a thickness of 10 μm on a sliding microtome and they were stored in cryoprotectant (30 % glycerol, 30 % ethoxyethanol, 40 % phosphate-buffered saline (PBS)). A single series of sections was processed for visualization of tyrosine hydroxylase (TH) via the biotin-labeled antibody procedure. Briefly, following several washes in a PBS solution containing 0.2 % Triton X-100 (PBS-T), endogenous peroxidase was quenched in a 3 % hydrogen peroxide solution and background staining was then inhibited in a 10 % normal goat serum/2 % bovine serum albumin solution. Tissue was then incubated with primary antibodies overnight: rabbit anti-TH antibody (1:500, Chemicon, USA). After three washes in PBS-T, sections were sequentially incubated in biotinylated goat anti-rabbit IgG (1:300; Vector, Burlingame, CA) for 1 h and the Elite avidin-biotin complex (ABC Kits; Vector, Burlingame, CA) for 1 h separated by three washes in PBS.
Immunostaining was visualized following a reaction with 3,3-diaminobenzidine (Vector, Burlingame, CA). Sections were then mounted on glass slides, allowed to dry, dipped into dH20, dehydrated through graded alcohols (70, 95, 100 %), cleared in xylenes, and coverslipped with DPX mounting medium (Electron Microscopy Sciences, Hatfield, PA).
Immunofluorescence
For immunostaining analysis, non-specific binding was blocked with 3 % bovine serum (Serotec, UK) and permeabilized with 0.1 % Triton X-100 in 1 % BSA-PBS for 30 min. The sections were incubated at 4 °C overnight with monoclonal rabbit anti-TH (1:500; Chemicon, USA) and anti-α-synuclein (1:2000, gift from Dr. Yu) and then incubated with corresponding secondary antibodies at room temperature for 2 h. The nucleus was stained by Hoechst 33342 (1 μg/ml, Sigma-Aldrich, USA). Control sections were run following identical protocols, but omitting the primary antibodies. For each animal, ten sections of brain were examined in a blinded fashion.
Stereology
TH-stained sections of the SN were used for stereological estimation of DArgic neuron numbers using optical fractionator from the Stereo Investigator software package (v. 7, MBF Biosciences, Williston, VT). The user was blinded to group assignment by coded slides. Ten sections spanning the entire anterior/posterior extent of the SN, each separated by 200 μm, were used for counting. All TH-ir neurons of the SN (pars compacta and pars reticulata) were included within each contour, of each section. Parameters used for TH stereological counting were grid size, 300 μm * 300 μm; counting frame, 80 μm * 80 μm; and 2-μm guard zones. Tissue thickness was determined by the user at each counting site. All final values represent estimated total by number weighted section thickness and were only included if their Gunderson coefficient of error (m = 1) was less than 0.09.
The Measurement of Striatal Biogenic Amines
Brains were used to dissect the two striata. The striatum samples were weighed and homogenized in 10 volumes of 0.2 M perchloric acid containing 100 μM EDTA-2Na and 100 ng/ml isoproterenol (the internal standard for the measurement of catecholamine and 5-HT contents) in a homogenizer at a maximum setting for 20 s on ice. After 30 min in the mixture of ice and water, the homogenate was centrifuged at 15,000×g for 20 min at 4 °C. The solution of 1 M sodium acetate was added to adjust the pH value to approximately 3.0. After filtration (0.45 μm), the samples were injected into a HPLC system.
DA, dihydroxyphenylacetic acid (DOPAC), and homovanillic acid (HVA) were detected using a HPLC-electrochemical detection system. Briefly, 20 μl of the homogenate sample was injected into the HPLC-electrochemical detection system by using an L-2200 auto sampler (HITACHI, Tokyo, Japan) at 4 °C, separated at 25 °C on a reverse-phase analytical column (EICOMPAK SC-5ODS, 3.0 mm × 150 mm; Eicom, Kyoto, Japan), eluted at a flow rate of 0.5 ml/min at 30 °C with 0.1 M sodium acetate/citric acid (pH 5.4) containing 17 % methanol, 190 mg/l sodium l-octanesulfonate, and 5 mg/l EDTA-2Na, and determined by an 2465 electrochemical detector (Waters, Milford, MA, USA). The chromatograms were recorded and analyzed using a computer with Millennium 32 Chromatography Manager Software (Waters) [37, 38].
Proteinase K Treatment
Tissue sections from AAV1/2 A53T α-synuclein-injected rats containing the SN were treated with proteinase K to determine whether the α-synuclein seen here was soluble (non-aggregated) or insoluble (aggregated) according to the method of Chu and Koprich et al. [20, 39, 40]. Briefly, sections were mounted and dried on Permafrost+ glass slides for at least 8 h at 55 °C. Sections were then briefly hydrated with TBS-T (10 mM Tris-HCl, pH 7.8; 100 mM NaCl; 0.05 % Tween 20) and digested with 50 μg/ml PK (Invitrogen, Carlsbad, CA) in TBS-T (10 mM Tris-HCl, pH 7.8; 100 mM NaCl; 0.1 % Tween 20) for a period of 2.5 h at 55 °C. Sections were then fixed for 10 min using 4 % paraformaldehyde and then processed for α-synuclein immunofluorescence as described above.
Western Blot Analysis
After removing the brain rapidly, the tissues were separated from the brain and then homogenized on ice with a microcontent motor-operated tissue homogenizer (Kimblekonts, USA) in ice-cold lysis buffer (1× PBS, 1 % Nonidet P-40, 0.5 % sodium deoxycholate, and 0.1 % SDS, RIPA) supplemented with protease inhibitors. Lysates were centrifuged at 10,000×g for 20 min at 4 °C; the supernatants were collected. Protein concentrations were determined by a Bradford protein assay. Equal amounts of protein (30 μg) were separated by SDS-PAGE and electroblotted onto nitrocellulose (NC) filter membrane (Millipore). After non-specific antibody binding was blocked with 5 % non-fat dry milk, membranes were incubated at 4 °C overnight with polyclonal rabbit anti-TH (1:2000, Millipore, USA), human α-synuclein (1:500, Zymed, San Francisco, CA), polyclonal rabbit anti-LC3 (1:1000, Novus, USA), and monoclonal rabbit anti-GAPDH (1:8000, Epitomics, USA). After washing in TBS-T, the immunoblots were incubated with horseradish peroxidase conjugated secondary antibodies (Cell Signaling Technology) for 1 h. The immunoblots were developed with enhanced chemiluminescence (ECL) reagents (Millipore, USA) and measured with Quantity Software (Bio-Rad, CA). To compare protein loading, antibody directed against GAPDH was used. Quantitative results were obtained from among at least six rats.
Statistical Analysis
All measurements stated above were repeated at least three times for each experiment, and the data were expressed as mean ± SEM. For all statistical comparisons, we first used a one-way ANOVA, with significance set at P < 0.05. If ANOVA was significant, all post hoc tests were conducted using Tukey’s multiple comparison test. The software used to conduct statistical analyses and graph all data was GraphPad Prism 4 (GraphPad Software, Inc.).
References
Dawson TM, Dawson VL (2003) Molecular pathways of neurodegeneration in Parkinson’s disease. Science 302:819–822
Siderowf A, Stern M (2003) Update on Parkinson disease. Ann Intern Med 138:651–658
Iwatsubo T (2003) Aggregation of alpha-synuclein in the pathogenesis of Parkinson’s disease. J Neurol 250(Suppl 3):III11–III14
Cole NB, Murphy DD, Lebowitz J, Di Noto L, Levine RL, Nussbaum RL (2005) Metal-catalyzed oxidation of alpha-synuclein: helping to define the relationship between oligomers, protofibrils, and filaments. J Biol Chem 280:9678–9690
Chiba-Falek O, Lopez GJ, Nussbaum RL (2006) Levels of alpha-synuclein mRNA in sporadic Parkinson disease patients. Mov Disord 21:1703–1708
Polymeropoulos MH, Lavedan C, Leroy E, Ide SE, Dehejia A, Dutra A, Pike B, Root H et al (1997) Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 276:2045–2047
Singleton AB, Farrer M, Johnson J, Singleton A, Hague S, Kachergus J, Hulihan M, Peuralinna T et al (2003) Hardy JGwinn-Hardy K: alpha-Synuclein locus triplication causes Parkinson’s disease. Science 302:841
Song W, Patel A, Qureshi HY, Han D, Schipper HM, Paudel HK (2009) The Parkinson disease-associated A30P mutation stabilizes alpha-synuclein against proteasomal degradation triggered by heme oxygenase-1 over-expression in human neuroblastoma cells. J Neurochem 110:719–733
Banerjee R, Beal MF, Thomas B (2010) Autophagy in neurodegenerative disorders: pathogenic roles and therapeutic implications. Trends Neurosci 33:541–549
Rubinsztein DC, Gestwicki JE, Murphy LO, Klionsky DJ (2007) Potential therapeutic applications of autophagy. Nat Rev Drug Discov 6:304–312
Ravikumar B, Duden R, Rubinsztein DC (2002) Aggregate-prone proteins with polyglutamine and polyalanine expansions are degraded by autophagy. Hum Mol Genet 11:1107–1117
Ravikumar B, Vacher C, Berger Z, Davies JE, Luo S, Oroz LG, Scaravilli F, Easton DF et al (2004) Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat Genet 36:585–595
Jain NK, Roy I (2009) Effect of trehalose on protein structure. Protein Sci 18:24–36
Richards AB, Krakowka S, Dexter LB, Schmid H, Wolterbeek AP, Waalkens-Berendsen DH, Shigoyuki A, Kurimoto M (2002) Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food Chem Toxicol 40:871–898
Emanuele E (2014) Can trehalose prevent neurodegeneration? Insights from experimental studies. Curr Drug Targets 15:551–557
Sarkar S, Davies JE, Huang Z, Tunnacliffe A, Rubinsztein DC (2007) Trehalose, a novel mTOR-independent autophagy enhancer, accelerates the clearance of mutant huntingtin and alpha-synuclein. J Biol Chem 282:5641–5652
Wolkers WF, Walker NJ, Tablin F, Crowe JH (2001) Human platelets loaded with trehalose survive freeze-drying. Cryobiology 42:79–87
Löw K, Aebischer P (2012) Use of viral vectors to create animal models for Parkinson’s disease. Neurobiol Dis 48:189–201
Koprich JB, Johnston TH, Huot P, Reyes MG, Espinosa M, Brotchie JM (2011) Progressive neurodegeneration or endogenous compensation in an animal model of Parkinson’s disease produced by decreasing doses of alpha-synuclein. PLoS One 6:e17698
Koprich JB, Johnston TH, Reyes MG, Sun X, Brotchie JM (2010) Expression of human A53T alpha-synuclein in the rat substantia nigra using a novel AAV1/2 vector produces a rapidly evolving pathology with protein aggregation, dystrophic neurite architecture and nigrostriatal degeneration with potential to model the pathology of Parkinson’s disease. Mol Neurodegener 5:43
Kabeya Y, Mizushima N, Ueno T, Yamamoto A, Kirisako T, Noda T, Kominami E, Ohsumi Y et al (2000) LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J 19:5720–5728
Sarkar S, Perlstein EO, Imarisio S, Pineau S, Cordenier A, Maglathlin RL, Webster JA, Lewis TA et al (2007) Small molecules enhance autophagy and reduce toxicity in Huntington’s disease models. Nat Chem Biol 3:331–338
Mizushima N, Yamamoto A, Matsui M, Yoshimori T, Ohsumi Y (2004) In vivo analysis of autophagy in response to nutrient starvation using transgenic mice expressing a fluorescent autophagosome marker. Mol Biol Cell 15:1101–1111
Tanida I, Ueno T, Kominami E (2004) LC3 conjugation system in mammalian autophagy. Int J Biochem Cell Biol 36:2503–2518
Pickford F, Masliah E, Britschgi M, Lucin K, Narasimhan R, Jaeger PA, Small S, Spencer B et al (2008) The autophagy-related protein beclin 1 shows reduced expression in early Alzheimer disease and regulates amyloid beta accumulation in mice. J Clin Invest 118:2190–2199
Webb JL, Ravikumar B, Atkins J, Skepper JN, Rubinsztein DC (2003) Alpha-Synuclein is degraded by both autophagy and the proteasome. J Biol Chem 278:25009–25013
Casarejos MJ, Solano RM, Gomez A, Perucho J, de Yebenes JG, Mena MA (2011) The accumulation of neurotoxic proteins, induced by proteasome inhibition, is reverted by trehalose, an enhancer of autophagy, in human neuroblastoma cells. Neurochem Int 58:512–520
Tanaka M, Machida Y, Niu S, Ikeda T, Jana NR, Doi H, Kurosawa M, Nekooki MNukina N (2004) Trehalose alleviates polyglutamine-mediated pathology in a mouse model of Huntington disease. Nat Med 10:148–154
Schaeffer V, Lavenir I, Ozcelik S, Tolnay M, Winkler DT, Goedert M (2012) Stimulation of autophagy reduces neurodegeneration in a mouse model of human tauopathy. Brain 135:2169–2177
Yu WB, Jiang T, Lan DM, Lu JH, Yue ZY, Wang J, Zhou P (2012) Trehalose inhibits fibrillation of A53T mutant alpha-synuclein and disaggregates existing fibrils. Arch Biochem Biophys 523:144–150
Lan DM, Liu FT, Zhao J, Chen Y, Wu JJ, Ding ZT, Yue ZY, Ren HM et al (2012) Effect of trehalose on PC12 cells overexpressing wild-type or A53T mutant α-synuclein. Neurochem Res 37:2025–2032
Sarkar S, Chigurupati S, Raymick J, Mann D, Bowyer JF, Schmitt T, Beger RD, Hanig JP et al (2014) Neuroprotective effect of the chemical chaperone, trehalose in a chronic MPTP-induced Parkinson’s disease mouse model. Neurotoxicology 44:250–262
Chung CY, Koprich JB, Siddiqi H, Isacson O (2009) Dynamic changes in presynaptic and axonal transport proteins combined with striatal neuroinflammation precede dopaminergic neuronal loss in a rat model of AAV alpha-synucleinopathy. J Neurosci 29:3365–3373
Minutoli L, Altavilla D, Bitto A, Polito F, Bellocco E, Lagana G, Fiumara T, Magazu S et al (2008) Trehalose: a biophysics approach to modulate the inflammatory response during endotoxic shock. Eur J Pharmacol 589:272–280
Echigo R, Shimohata N, Karatsu K, Yano F, Kayasuga-Kariya Y, Fujisawa A, Ohto T, Kita Y et al (2012) Trehalose treatment suppresses inflammation, oxidative stress, and vasospasm induced by experimental subarachnoid hemorrhage. J Transl Med 10:80
Li J, Roubeix C, Wang Y, Shi S, Liu G, Baudouin C, Chen W (2012) Therapeutic efficacy of trehalose eye drops for treatment of murine dry eye induced by an intelligently controlled environmental system. Mol Vis 18:317–329
Wang YQ, Tu ZC, Xu XY, Li R, Qu WM, Urade Y, Huang ZL (2012) Acute administration of fluoxetine normalizes rapid eye movement sleep abnormality, but not depressive behaviors in olfactory bulbectomized rats. J Neurochem 120:314–324
Hong ZY, Huang ZL, Qu WM, Eguchi N (2005) Orexin A promotes histamine, but not norepinephrine or serotonin, release in frontal cortex of mice. Acta Pharmacol Sin 26:155–159
Chu Y, Kordower JH (2007) Age-associated increases of alpha-synuclein in monkeys and humans are associated with nigrostriatal DA depletion: Is this the target for Parkinson’s disease? Neurobiol Dis 25:134–149
Chu Y, Dodiya H, Aebischer P, Olanow CW, Kordow-er JH (2009) Alterations in lysosomal and proteasomal markers in Parkinson’s disease: relationship to alpha-synuclein inclusions. Neurobiol Dis 35:385–398
Acknowledgments
This work was supported by grants from the National Foundation of Natural Science of China (No.81071018 and No.81371413), key project from Science and Technology Commission of Shanghai Municipality (13JC1401103), and project of Shanghai Municipal Commission of Health (XBR2013088). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. We thank Dr. Shun Yu from Xuanwu Hospital of the Capital University of Medical Sciences, Beijing, China, for kindly providing the human α-synuclein antibody for immunofluorescence.
Conflict of interest
The authors declare that they have no competing interests.
Authors’ contributions
WJ, Koprich JB, and Brotchie JM designed this study. HQ, WY, and YWB performed this study, analyzed data, and wrote the manuscript. XBG assisted in behavioral analysis. All authors discussed the results and provided comments on the manuscript. All authors read and approved the final manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Qing He and James B. Koprich contributed equally to this work.
Rights and permissions
About this article

Cite this article
He, Q., Koprich, J.B., Wang, Y. et al. Treatment with Trehalose Prevents Behavioral and Neurochemical Deficits Produced in an AAV α-Synuclein Rat Model of Parkinson’s Disease. Mol Neurobiol 53, 2258–2268 (2016). https://doi.org/10.1007/s12035-015-9173-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-015-9173-7