
Abstract
Astrocyte activation, associated with the release of pro-inflammatory cytokines interleukin 1-β (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor alpha (TNF-α), is a hallmark of multiple brain diseases, including mesial temporal lobe epilepsy. In recent years, several microRNAs have emerged as important controllers of Toll-like receptor (TLR) signaling. In this study, we investigated the effect of miR-132, miR-146a, and miR-155 on myeloid-related protein-8 (MRP8) induced astrocyte-related inflammation. Using quantitative polymerase chain reaction (qPCR) and western blot, we found clear upregulation of TLR4 and downstream inflammatory cytokines, along with dysregulation of miR-132, miR-146a, and miR-155 in in vitro astrocytes after exposing them to different concentrations of MRP8. In addition, we focused on the effect of miR-132 on astrocyte-related inflammation induced by MRP8 via lentiviral infection then evaluated the expression of its possible target genes: acetylcholinesterase (AChE) and interleukin-1 receptor-associated kinase (IRAK4). Our results show that miR-132 is a negative feedback regulator of IL-1β and IL-6, but not TNF-α, by targeting IRAK4. Together, our findings demonstrate the novel role of TLR4-related microRNAs, especially miR-132, in the regulation of MRP8-induced astrocyte activation and highlight the importance of miR-132 in the modulation of innate immune response induced by endogenous ligands in neurological diseases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Astrocytes, constituting the major glial cell population in the central nervous system (CNS), play a crucial role in the regulation of immune response in many human CNS diseases (De Keyser et al. 2008; Seifert et al. 2006). They become activated under a diversity of insults, with changes in morphology, proliferation, and molecular expression (Hamby and Sofroniew 2010; Sofroniew 2009). This reactive astrogliosis, associated with the release of pro-inflammatory cytokines interleukin 1-β (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor alpha (TNF-α), is a hallmark of multiple brain diseases, including mesial temporal lobe epilepsy (MTLE), migraines, brain edema, and neurodegenerative disorders (Carpentier et al. 2005; Dong and Benveniste 2001; Li et al. 2011; Vezzani et al. 2008).
Toll-like receptors (TLRs) are a family of transmembrane proteins of mammalian cells. They are widely expressed in many central nervous system cells and play key roles in the response of innate immunity cells to pathological stimuli (Brown et al. 2011; Bsibsi et al. 2002). Besides recognition of bacterial and viral components, TLRs can also play a part in the recognition of several endogenous ligands, including myeloid-related protein-8 (MRP8) (Gill et al. 2010).
MicroRNAs (miRNAs) are a class of gene regulators that block subsequent protein production either by inhibiting translation or promoting RNA degradation at the post-transcriptional level (Filipowicz et al. 2008). In recent years, several miRNAs have emerged as important controllers of TLR signaling, which is being extensively researched (O’Neill et al. 2011). Indeed, they may contribute to the pathogenesis of endotoxin tolerance and major age-related diseases, such as cardiovascular diseases, diabetes, Alzheimer’s disease (AD), and cancers (Nahid et al. 2011a; Olivieri et al. 2013). Although there is a perception that miR-146a, miR-155, and miR-21 are key regulators of TLR signaling (Quinn and O’Neill 2011), miR-132 has been shown to be upregulated in response to several TLRs in multiple monocytic cells (Nahid et al. 2011a, 2013). miR-132 is a cyclic AMP-responsive element binding (CREB)-regulated miRNA (Cheng et al. 2007). It was classified as a “neurimmiR” that functions in the “cross-talk” between neuronal and immune systems (Soreq and Wolf 2011). Currently, most studies are focused on its neuronal functions, such as the regulation of neurite outgrowth, spine morphology, and cortical plasticity (Bicker et al. 2014; Hancock et al. 2014; Scott et al. 2012). However, the immune functions of this miRNA in the CNS have not been completely illustrated. Soreq and his colleagues have reported that miR-132 develops an anti-inflammatory effects by suppressing acetylcholinesterase (AChE) (Shaked et al. 2009). This enzyme limits the level of acetylcholine (ACh), an important inhibitor of peripheral inflammation and one of the characterized mediators of “cholinergic antiinflammatory pathway” (Tracey 2007). In addition, Soreq showed that excess AChE in transgenic mice induces astrocytic activation (Sternfeld et al. 2000). These implied that AChE may play a role in the regulation of astrocytic inflammation. Another study demonstrated that miR-132 plays a crucial role in regulating TLR2-mediated tolerance and cross-tolerance through targeting interleukin-1 receptor-associated kinase (IRAK4) (Nahid et al. 2013).
In this study, we first detected the changes of TLR4 and downstream signaling in the MRP8/TLR4 pathway, including IL-1β, IL-6, and TNF-α, in astrocytes with different concentrations of MRP8 stimulation. The expression of miR-132, miR-146a, and miR-155 in astrocytes was also evaluated by quantitative polymerase chain reaction (qPCR). We focused on the effect of miR-132 modulation on MRP8-induced astrocyte-related inflammation via lentiviral infection, and then evaluated the expression of its possible target genes: AChE and IRAK4.
Materials and Methods
Cell Cultures
The U251 human astrocytoma cell line was obtained from the Department of Cell Biology in Xiangya Medical College of Central South University. Cells were cultured in high glucose of Dulbecco’s modified Eagle’s medium (DMEM; Hyclone, USA) supplemented with 10 % fatal bovine serum (FBS), 100 units/ml penicillin, and 100 μg/ml streptomycin and were kept at 37 °C in humid 5 % CO2/95 % air.
Cell Treatments
Prepared U251 cells were cultured without serum DMEM for 24 h. Then cells were stimulated with increasing concentrations (0.125 μg/ml, 0.25 μg/ml, 0.5 μg/ml, 0.75 μg/ml, 1.0 μg/ml) of purified MRP8 protein (Abcam, Hong Kong) for 24 h. Or, cells were cultured with no serum medium for 24 h after lentivirus transfection, then stimulated with 0.75 μg/ml MRP8 for 24 h. Stimulated cells were used for RNA and protein extraction.
RNA Extraction
For RNA isolation, prepared human U251 cells (1 × 106) from three different experimental sets were homogenized in 1 ml Trizol Reagent (Invitrogen, USA). After adding 0.2 ml of chloroform, mixed vigorously for 15 s, the aqueous phase was isolated by centrifuging at 4 °C at a speed of 12,000 rpm for 15 min. RNA was precipitated with 0.5 ml isopropyl alcohol, washed twice with 75 % ethanol by centrifugation at 4 °C at a speed of 7500 rpm for 5 min, then dissolved in 20–50 μl nuclease-free water. The concentration and purity of RNA were determined at 260/280 nm using a Nanodrop spectrophotometer (Thermo, USA).
Western Blot Analysis
The U251 human astrocytoma cells were washed with PBS before collecting for total protein extraction on ice. After adding NP-40 lysis solution, the homogenate was centrifuged at 12,000 rpm for 30 min at 4 °C. The supernate was collected, and the protein concentration was determined by a bicinchoninic acid (BCA) protein assay kit. Equal amounts of protein (50 μg) were separated by a 10–15 % SDS-PAGE gel and transferred onto a PVDF membrane. The membrane was blocked with 5 % nonfat milk in TBST for 1 h at room temperature. The proteins were detected with specific primary antibodies (TLR4 (1:200; Abcam), IL-1β (1:500; Novus Biologicals), IL-6 (1:1000; Abcam), TNF-α (1:1000; R&D), and IRAK4 (1:1000; Cell Signaling)) with gentle shaking at 4 °C overnight. The expression of β-actin (1:10,000; Sigma) was used as an internal reference. Anti-mouse/rabbit IgG (1:7500; Jackson Immuno Research) peroxidase reactions were used for immunostaining. The membranes were developed using an enhanced chemiluminescence (ECL) kit (Millipore) and exposed to X-ray film. Image J software (Nation Institutes of Health) was used to quantitatively analyze protein expression levels.
Lentivirus Transfection
Packaging and production of lentivirus, which carry the hsa-pri-miR-132-3p precursor, hsa-anti-miR-132-3p, and their controls were obtained from Shanghai GeneChem Biotech Co. (Shanghai, China). The lentiviral transduction was carried out according to the manufacturer’s instructions (GeneChem). Astrocytoma cells were cultured in six-well uncoated plastic plates at a density of 5 × 105 cells/well and divided into five groups: blank control group (without lentivirus vector (Con)), miR-132 over-expression group (miR-132-up) and its negative control group (miR-132up-con), and the down-expression group (miR-132-down) and its negative control group (miR-132down-con). Cells were infected with a multiplicity of infection (MOI) of 50 for 10 h at 37 °C, followed by addition of fresh medium. Three days later, cells infected with lentiviral vectors were selected using PURO at a concentration of 0.25 μg/ml for 48 h. The transfection efficiency was evaluated by enhanced green fluorescent protein (GFP) and validated by qPCR. Cells were harvested for RNA 5 days after infection or stimulated with MRP8 for further experiments.
TLR4, IL-1β, IL-6, TNF-α, AChE, and IRAK4 Expression Evaluated by qPCR
We used the following primers: TLR4 (forward: atc atc cag gaa ggc ttc ca, reverse: gct gcc tca gca agg act tct), IL-1β (forward: gcc aac aag tgg tat tct cca, reverse: tgc cgt ctt tca tca cac ag), IL-6 (forward: gtc aac tcc atc tgc cct tc, reverse: tgt ggg tgg tat cct ctg tg), TNF-α (forward: aag gag gag aag ttc cca aa, reverse: tct ttg aga tcc atg cca tt), AChE (forwad: ggg gct cag cag tac gtt ag, reverse: gtc gag cgt gtc ggt gg), IRAK4 (forward: cga gcg aca gtg aca acc t, reverse: atg tgc cag gaa agt ggt g), and β-actin (forward: ccc atc tat gag ggt tac gc, reverse: ttt aat gtc acg cac gat ttc). Complementary DNA (cDNA) was synthesized from 500 pg of total RNA using a Prime Script RT Reagent Kit (TAKARA, Dalian, China). RNase-free dH2O was added to make the total reaction volume 10 μl. TLR4, IL-1β, IL-6, TNF-α, AChE, and IRAK4 expressions were analyzed using SYBR® Premix ExTaqTM II (TAKARA, Dalian, China), which was run on the BioRad CFX384 Touch Real-Time PCR detection system. All procedures were performed according to the manufacturer’s instructions. All assays were performed in triplicate. Data analysis was performed using the 2−(△△CT) method to determine the relative quantitative level and was expressed as a fold-difference to the relevant control (recognized as 1 ± 0.00). β-actin expression was used as an internal reference.
miR-132, miR-146a, and miR-155 Expression Evaluated by qPCR
miRNA primer was purchased from GeneCopoeia. The catalog of each primer is as follows. Hsa-miR-132-3p: HmiRQP0161. Hsa-miR-146a-5p: HmiRQP0196. Has-miR-155-5p: HmiRQP0221. Homo sapiens snRNA U6: HmiRQP9001. cDNA synthesis was performed using the One Step PrimeScript® miRNA cDNA Synthesis Kit (TAKARA, RR716) according to the manufacturer’s instructions. Total RNA was 500 pg, and total reaction volume was 20 μl. The qPCR reaction was performed using the SYBR® Premix Ex Taq™ II (TAKARA, RR716) kit. Data analysis was performed using the 2−(△△CT) method to determine the relative quantitative level and was expressed as a fold-difference to the relevant control (recognized as 1 ± 0.00). The expression of the U6 small nuclear RNA gene was used as an internal reference.
Statistical Analysis
Data are presented as the mean ± standard deviation for at least three independent experiments. We performed a Student’s t test to determine significant differences between two groups. One-way analysis of variance was utilized to determine significant differences among multiple groups. A p value of less than 0.05 was considered statistically significant.
Results
The Effects of MRP8 Treatment on the Expression of TLR4, IL-1β, IL-6, and TNF-α
qPCR and western bolt was performed to examine the expression of TLR4, IL-1β, IL-6, and TNF-α in U251 human astrocytoma cells after exposure to different concentrations of MRP8. Our qPCR results showed that the gene expression of TLR4, IL-1β, and IL-6 had a similar tendency. All of them increased significantly after MRP8 stimulation at a concentration of 0.125 μg/ml and reached the highest point at a concentration of 0.75 μg/ml (Fig. 1a–c). Expression changes of TNF-α were not as obvious as with IL-1β or IL-6. TNF-α expression began to rise significantly at a concentration of 0.125 μg/ml, and increased the most at 0.5 μg/ml (Fig. 1d). Western blot results showed the same tendency with gene expressions (Fig. 1e, f).

Expression of TLR4, IL-1β, IL-6, and TNF-α after stimulation with MRP8. U251 human astrocytoma cells were left untreated or were treated for 24 h with the indicated additions. a–c TLR4, IL-1β, and IL-6 mRNA increased significantly at 0.125 μg/ml MRP8 and reached the highest point after exposure to 0.75 μg/ml MRP8. d Expression of TNF-α mRNA increased significantly after exposure to 0.125 μg/ml MRP8, reaching the highest point at 0.5 μg/ml MRP8. e, f Western blot results showed the same tendency with gene expressions (*p < 0.05)
The Effects of MRP8 Treatment on the Expression of miR-132, miR-146a, and miR-155
To verify that miR-132, miR-146a, and miR-155 participate in MRP8-induced inflammation, we exposed U251 human astrocytoma cells to different concentrations of MRP8 for 24 h. As shown in Fig. 2a, b, the expression of miR-132 and miR-146a began to increase significantly compared with controls after exposure to 0.5 μg/ml MRP8. As the concentration of MRP8 increased, the expression of miR-132 also increased. The expression of miR-146a reached its top point at a concentration of 0.75 μg/ml MRP8, then declined from that point on. The general trend of miR-155 expression was similar to miR-146a, except that it augmented significantly with a concentration level of 0.125 μg/ml and gradually reached its peak at 0.75 μg/ml (Fig. 2c).

Gene expression of miR-132, miR-146a, and miR-155 after exposure to MRP8. a Expression of miR-132 began to increase significantly after exposure to 0.5 μg/ml MRP8. As MRP8 concentration increased, the expression of miR-132 upregulated further. b miR-146a upregulated significantly compared with controls after exposure to 0.5 μg/ml MRP8 and reached its top point after exposure to 0.75 μg/ml MRP8. c The expression of miR-155 augmented significantly at a concentration of 0.125 μg/ml MRP8 and reached its peak after exposure to 0.75 μg/ml MRP8 (* p < 0.05)
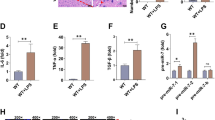
The Effects of Modulated miR-132 on the Expression of IL-1β, IL-6, and TNF-α
Over 80 % of U251 human astrocytoma cells infected with lentivirus expressed enhanced green fluorescent protein (GFP) (Fig. 3a). We also validated that using qPCR method (Fig. 3b). qPCR results showed that the expression of IL-1β, IL-6, and TNF-α increased significantly after MRP8 stimulation compared with controls (Fig. 4). Downregulation of miR-132 before MRP8 stimulation promoted the release of both IL-1β and IL-6, whereas upregulation of miR-132 had the opposite effect (Fig. 4a, b). Pre-downregulation or pre-upregulation of miR-132 before MRP8 stimulation had no effect on the release of TNF-α compared with MRP8 stimulation alone (Fig. 4c).

The lentivirus transfection efficiency of astrocytes. a The enhanced green fluorescent protein (GFP)-positive rate of U251 human astrocytoma cells after transfection was more than 80 %. Scale bar in (a), 25 μm. b, c qPCR of miR-132 after lentivirus transfection. miR-132 expression in the miR-132-down group was significantly lower than in its negative control group (miR-132down-con). And in the miR-132-up group, miR-132 expression was significantly higher than in its control group. (*p < 0.05; **p < 0.01)

Effect of miR-132 on the MRP8-induced release of IL-1β, IL-6, and TNF-α. a–c IL-1β, IL-6, and TNF-α increased significantly after MRP8 stimulation compared with controls. Downregulation of miR-132 before MRP8 stimulation promoted the release of both IL-1β and IL-6, whereas upregulation of miR-132 had the opposite effect. Downregulation or upregulation of miR-132 before MRP8 stimulation had no effect on the release of TNF-α compared with MRP8 stimulation alone. (*p < 0.05; # p > 0.05)
The Effects of Modulated miR-132 on the Expression of AChE and IRAK4
Our qPCR result showed that the expression of AChE increased conspicuously in U251 human astrocytoma cells after MRP8 stimulation compared with control. But it did not show an inhibited regulatory role with aberrant expression of miR-132 (Fig. 5a). IRAK4 is an important kinase in TLR4 signaling and has been verified as a target of miR-132 (Nahid et al. 2013). Here, both our qPCR and western blot results showed that IRAK4 increased significantly after MRP8 pretreatment compared with control. Greatly upregulated miR-132 through lentivirus transfection could inhibit the expression of IRAK4. And pre-downregulation of miR-132 significantly promoted the expression of IRAK4, whereas upregulation of miR-132 before MRP8 stimulation had the opposite effect (Fig. 5b–d).

AChE and IRAK4 expression after regulating miR-132 in astrocytes exposed to MRP8. a AChE mRNA increased significantly after MRP8 stimulation in U251 human astrocytoma cells. Modulation miR-132 via lentivirus transfection has no effect on AChE mRNA expression. mRNA expression (b) and protein expression of IRAK4 (c, d) after exposure to MRP8 for 24 h in U251 human astrocytoma cells transfected with anti-miR-132 or pre-miR-132 lentivirus vector (MOI = 50). Greatly upregulated miR-132 could inhibit the expression of IRAK4, and downregulated miR-132 promotes the expression of IRAK4. IRAK4 increased significantly after MRP8 pretreatment in U251 cells compared with the control group. Pre-downregulation of miR-132 enhanced the effect of MRP8 inducing the expression of IRAK4, whereas upregulation of miR-132 before MRP8 stimulation had the opposite effect (*p < 0.05; **p < 0.01; # p > 0.05). MRP8 myeloid-related protein 8, MOI multiplicity of infection
Discussion
Accumulating experimental evidence suggests that pro-inflammatory molecules can alter glio-neuronal communication, contributing to the generation of seizures and seizure-related neuronal damage (Devinsky et al. 2013; Vezzani et al. 2013). Although microglia are an important type of immune cell of the CNS, astrocytes are thought to be both the major source and targets of epileptogenic inflammatory signaling (Aronica et al. 2012). Our previous findings showed that astrocyte-related neuroinflammation induced by MRP8 may play an important role in the development of immature MTLE (Gan et al. 2014). In this study, we now report the dose-dependent relationship of MRP8, an endogenous ligand of TLR4, and inflammation-related miR-132, miR-146a, and miR-155 in U251 human astrocytoma cells. Furthermore, we detected the expression of proinflammatory cytokines IL-1β, IL-6, and TNF-α and found that miR-132 participated in the regulation of IL-1β and IL-6 secretion but not TNF-α, by targeting IRAK4. This finding highlights the importance of miR-132 in the modulation of the innate immune response induced by endogenous ligands in the CNS.
TLR4 is one of the membrane-bound pattern-recognition receptors profoundly expressed in astrocytes (Bsibsi et al. 2002). It can sense a variety of microbial-specific motifs or endogenous ligands to trigger an innate immune response. TLR4 activation results in the induction of transcriptional factors such as nuclear factor-κB (NF-κB), which has the ability to trigger various pro-inflammatory genes such as those encoding cytokines, chemokines, proteins of the complement system, cyclooxygenase-2, and inducible nitric oxide. As an endogenous ligand of TLR4, MRP8 participates in the neuroinflammation of multiple CNS diseases, including MTLE, traumatic brain injury, and ischemic and neurodegenerative diseases (Engel et al. 2000; Gan et al. 2014; Sheng et al. 1994; Ziegler et al. 2009). Our study is the first to demonstrate the dose-dependent relationship of MRP8 and TLR4 activation in astrocytes in vitro. Although MRP8 always presents as a heterocomplex together with MRP14 (Floris et al. 2004; Viemann et al. 2007; Vogl et al. 2004), it has been argued that MRP8 is the active component that binds to the receptor site (Vogl et al. 2007; Yonekawa et al. 2011). The significant activation of TLR4 after MRP8 exposure in our experiments supports that point. Furthermore, we found that the expression of IL-1β, IL-6, and TNF-α, downstream inflammatory cytokines in the MRP8/TLR4 pathway, increased to various degrees. In our previous study, we reported elevated expression of IL-1β along with activation of the MRP8/TLR4/NF-κB pathway in astrocytes (Gan et al. 2014). In addition, other research has shown that activation of the TLR4/NF-κB signal pathway results in the transcription of TNF-α, IL-1α and IL-1β in other immune cells (Vainas et al. 2006; Vogl et al. 2007; Ziegler et al. 2009). So we speculate that MRP8 may influence the expression of IL-1β, IL-6, and TNF-α in a similar fashion. These findings provide a possible mechanism for how MRP8 contributes to neurological diseases.
The discovery of miRNAs represents a tremendous revolution in the regulation of transcription and translation. Activation of mammalian immune response needs exquisite manipulation. The capability of miRNAs to fine-tune gene expression naturally lends itself to the regulation of the immune system (Virtue et al. 2012). Therefore, this study focused on the relationship between TLR4-related miR-132, miR-146a, and miR-155 and MRP8-induced inflammation in astrocytes.
The dysregulation of miR-132 is associated with several neurological disorders, such as Toxoplasma gondii infection, MTLE, primary glioblastoma multiforme, schizophrenia, AD, and Parkinson’s disease (Parker et al. 2013; Peng et al. 2013; Tognini and Pizzorusso 2012; Xiao et al. 2014). However, the underlying molecular mechanisms are still incompletely illustrated. In this study, we report a dose-dependent reaction between miR-132 upregulation and MRP8 stimulation in astrocytes. To further evaluate the function of miR-132 in response to MRP8 stimulation, we investigated the effect of miR-132 on IL-1β, IL-6, and TNF-α, three major inflammatory cytokines related to neuroinflammation and epilepsy (Li et al. 2011; Shimada et al. 2014; Vezzani et al. 2013). Our results show that upregulation of miR-132 reduced IL-1β and IL-6 gene levels in cultures stimulated with MRP8, while downregulation of miR-132 had the opposite effect. This finding testifies to the fact that miR-132 is a negative feedback regulator of IL-β and IL-6 expression in activated astroctyes. Combined with our previous in vivo studies (Gan et al. 2014; Peng et al. 2013), increased miR-132 probably protects the immature brain by suppressing the neuroinflammation that would facilitate seizures. miR-132 can also modulate antiviral innate immunity induced by Kaposi’s sarcoma-associated herpesvirus (KSHV) infection through suppression of the p-300 transcriptional co-activator (Lagos et al. 2010). Due to the homologous sequence, each miRNA has the potential to regulate the translation of hundreds of genes (Selbach et al. 2008). Coinciding with previous study (Nahid et al. 2013), we found that miR-132 might influence the MRP8-induced activation of astrocytes by targeting IRAK4, a pivotal adaptor kinase. As illustrated in Fig. 6, IRAK4 is recruited to MyD88 after TLR4 activation, forming a helical assembly of the Myd88-IRAK4-IRAK2/1 complex that further activates TNFR-associated factor 6 (TRAF6) and eventually leads to gene transcription and inflammation (Nahid et al. 2011b). In cultured glia, AChE levels increase in response to dibutyrylcyclic-AMP (Karpel et al. 1996). Moreover, AChE activities were found in astrocytoma and glioblastoma tumors (Grisaru et al. 1999). Consistently, we found significant upregulation of AChE messenger RNA (mRNA) after MRP8 stimulation in this study. Although miR-132 can attenuate inflammation by suppressing AChE, enhancing the brain’s ability to govern inflammation via cholinergic signaling in macrophages (O'Neill 2009; Shaked et al. 2009). What is more, it may participate in amelioration of the inflammation in human inflammatory bowel disease (Maharshak et al. 2013) and mediate stress-inducible cognitive deficits through targeting AChE (Shaltiel et al. 2013). Nevertheless, the expression of AChE mRNA was not influenced by modulating miR-132 in our study. This may be due to that different circumstances call for different manipulations of the AChE transcript (Shaked et al. 2009; Berson et al. 2008; Sklan et al. 2004). In addition, the impact of altering a miRNA on a biological process is influenced by several factors, including the expressions level of the miRNA and the mRNA target (Ebert and Sharp 2012). AChE levels are known to be very low in all types of normal glia (Karpel et al. 1996).

miR-132 negative regulation on MRP8-induced inflammation in astrocytes. MRP8 binds to TLR4 and activates the MyD88-dependent pathway. This eventually leads to gene transcription with the production of IL-1β, IL-6, and TNF-α. Upregulated miR-132 in turn negatively regulates the release of inflammatory factors by targeting IRAK4
miR-146a is a NF-κB-dependent miRNA and plays an important role in innate immune regulation (Taganov et al. 2006). It is known to target key elements of the MyD88 signaling pathway (such as IRAK1 and TRAF6), modulating the inflammatory response mediated by several TLR ligands in endotoxin tolerance (Nahid et al. 2011a). As observed in the MTLE hippocampal region, miR-146a is mainly localized in activated astrocytes and increases significantly in the acute and latent stages of disease (Aronica et al. 2010; Omran et al. 2012). Our study consistently found that miR-146a increased significantly after exposure to 0.5 μg/ml MRP8, accompanied by an obvious release of IL-1β and IL-6. This specific location and expression pattern could help miR-146a to modulate seizure-related neuroinflammation. Other studies have also indicated that the aberrant expression of miR-146a contributes to the age-associated dysfunction of macrophages (Jiang et al. 2012). Specifically, Anand found that miR-146a was a negative feedback regulator of IL-1β-induced inflammation in both primary human astroctyes and the astrocytoma cell line by targeting IRAK1, IRAK2, and TRAF6 (Iyer et al. 2012). All of the above findings indicate that miR-146a may be involved in the regulation of astroctye activation induced by endogenous ligand MRP8 through modification of different target genes together with miR-132.
Although the expression of pro-inflammatory factor TNF-α also increased with MRP8 stimulation, the changes were much weaker compared with IL-1β and IL-6. We speculate that it is due to TNF-α not playing a main role in this inflammatory process. It is worth noting that modulating miR-132 before MRP8 stimulation did not influence the expression of TNF-α in our experiment. Although, they observed dose-dependent production of TNF-α in PBMCs and THP-1 monocytes after stimulation with several TLR ligands accompanying the upregulation of several miRNAs (Nahid et al. 2013). It is widely believed that the regulation of TNF-α is associated with miR-155. In our previous report, we found that suppressing TNF-α with its inhibitor resulted in a significant decline of miR-155 (Omran et al. 2013). Other reports have also shown a direct relationship between TNF-α and miR-155 in other diseases, such as epilepsy (Ashhab et al. 2013), B cell lymphomas (Pedersen et al. 2009), alcoholic liver disease (Bala et al. 2011), and septic shock (Tili et al. 2007). Therefore, it is very likely that the upregulation of miR-155 after MRP8 stimulation is mainly responsible for the changes in TNF-α in our study.
In summary, we report the activation of MRP8/TLR4 signaling in vitro in astrocytes and the effect on downstream inflammatory cytokines (IL-1β, IL-6 and TNF-α) accompanying the upregulation of miR-132, miR-146a, and miR-155. Furthermore, we demonstrate that miR-132 modulates the release of IL-1β and IL-6 induced by MRP8 in astrocytes by targeting IRAK4. These findings open up a new perspective on modulating astrocyte-related neuroinflammation in noninfectious CNS diseases.
References
Aronica E, Fluiter K, Iyer A et al (2010) Expression pattern of miR-146a, an inflammation-associated microRNA, in experimental and human temporal lobe epilepsy. Eur J Neurosci 31:1100–1107
Aronica E, Ravizza T, Zurolo E, Vezzani A (2012) Astrocyte immune responses in epilepsy. Glia 60:1258–1268
Ashhab MU, Omran A, Kong H et al (2013) Expressions of tumor necrosis factor alpha and microRNA-155 in immature rat model of status epilepticus and children with mesial temporal lobe epilepsy. J Mol Neurosci 51:950–958
Bala S, Marcos M, Kodys K et al (2011) Up-regulation of microRNA-155 in macrophages contributes to increased tumor necrosis factor {alpha} (TNF{alpha}) production via increased mRNA half-life in alcoholic liver disease. J Biol Chem 286:1436–1444
Berson A, Knobloch M, Hanan M et al (2008) Changes in readthrough acetylcholinesterase expression modulate amyloid-beta pathology. Brain 131:109–119
Bicker S, Lackinger M, Weiß K, Schratt G (2014) MicroRNA-132, −134, and −138: a microRNA troika rules in neuronal dendrites. Cell Mol Life Sci 71:3987–4005
Brown J, Wang H, Hajishengallis GN, Martin M (2011) TLR-signaling networks: an integration of adaptor molecules, kinases, and cross-talk. J Dent Res 90:417–427
Bsibsi M, Ravid R, Gveric D, van Noort JM (2002) Broad expression of Toll-like receptors in the human central nervous system. J Neuropathol Exp Neurol 61:1013–1021
Carpentier PA, Begolka WS, Olson JK, Elhofy A, Karpus WJ, Miller SD (2005) Differential activation of astrocytes by innate and adaptive immune stimuli. Glia 49:360–374
Cheng HY, Papp JW, Varlamova O et al (2007) microRNA modulation of circadian-clock period and entrainment. Neuron 54:813–829
De Keyser J, Mostert JP, Koch MW (2008) Dysfunctional astrocytes as key players in the pathogenesis of central nervous system disorders. J Neurol Sci 267:3–16
Devinsky O, Vezzani A, Najjar S, De Lanerolle NC, Rogawski MA (2013) Glia and epilepsy: excitability and inflammation. Trends Neurosci 36:174–184
Dong Y, Benveniste EN (2001) Immune function of astrocytes. Glia 36:180–190
Ebert MS, Sharp PA (2012) Roles for microRNAs in conferring robustness to biological processes. Cell 149:515–524
Engel S, Schluesener H, Mittelbronn M et al (2000) Dynamics of microglial activation after human traumatic brain injury are revealed by delayed expression of macrophage-related proteins MRP8 and MRP14. Acta Neuropathol 100:313–322
Filipowicz W, Bhattacharyya SN, Sonenberg N (2008) Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet 9:102–114
Floris S, van der Goes A, Killestein J et al (2004) Monocyte activation and disease activity in multiple sclerosis. A longitudinal analysis of serum MRP8/14 levels. J Neuroimmunol 148:172–177
Gan N, Yang L, Omran A et al (2014) Myoloid-related protein 8, an endogenous ligand of Toll-like receptor 4, is involved in epileptogenesis of mesial temporal lobe epilepsy via activation of the nuclear factor-kappaB pathway in astrocytes. Mol Neurobiol 49:337–351
Gill R, Tsung A, Billiar T (2010) Linking oxidative stress to inflammation: Toll-like receptors. Free Radic Biol Med 48:1121–1132
Grisaru D, Sternfeld M, Eldor A, Glick D, Soreq H (1999) Structural roles of acetylcholinesterase variants in biology and pathology. Eur J Biochem 264:672–686
Hamby ME, Sofroniew MV (2010) Reactive astrocytes as therapeutic targets for CNS disorders. Neurotherapeutics 7:494–506
Hancock ML, Preitner N, Quan J, Flanagan JG (2014) MicroRNA-132 is enriched in developing axons, locally regulates Rasa1 mRNA, and promotes axon extension. J Neurosci 34:66–78
Iyer A, Zurolo E, Prabowo A et al (2012) MicroRNA-146a: a key regulator of astrocyte-mediated inflammatory response. PLoS One 7:e44789
Jiang M, Xiang Y, Wang D et al (2012) Dysregulated expression of miR-146a contributes to age-related dysfunction of macrophages. Aging Cell 11:29–40
Karpel R, Sternfeld M, Ginzberg D, Guhl E, Graessmann A, Soreq H (1996) Overexpression of alternative human acetylcholinesterase forms modulates process extensions in cultured glioma cells. J Neurochem 66:114–123
Lagos D, Pollara G, Henderson S et al (2010) miR-132 regulates antiviral innate immunity through suppression of the p300 transcriptional co-activator. Nat Cell Biol 12:513–519
Li G, Bauer S, Nowak M et al (2011) Cytokines and epilepsy. Seizure 20:249–256
Maharshak N, Shenhar-Tsarfaty S, Aroyo N et al (2013) MicroRNA-132 modulates cholinergic signaling and inflammation in human inflammatory bowel disease. Inflamm Bowel Dis 19:1346–1353
Nahid MA, Satoh M, Chan EK (2011a) MicroRNA in TLR signaling and endotoxin tolerance. Cell Mol Immunol 8:388–403
Nahid MA, Satoh M, Chan EK (2011b) Mechanistic role of microRNA-146a in endotoxin-induced differential cross-regulation of TLR signaling. J Immunol 186:1723–1734
Nahid MA, Yao B, Dominguez-Gutierrez PR, Kesavalu L, Satoh M, Chan EK (2013) Regulation of TLR2-mediated tolerance and cross-tolerance through IRAK4 modulation by miR-132 and miR-212. J Immunol 190:1250–1263
O’Neill LA, Sheedy FJ, McCoy CE (2011) MicroRNAs: the fine-tuners of Toll-like receptor signalling. Nat Rev Immunol 11:163–175
Olivieri F, Rippo MR, Prattichizzo F et al (2013) Toll like receptor signaling in “inflammaging”: microRNA as new players. Immun Ageing 10:11
Omran A, Peng J, Zhang C et al (2012) Interleukin-1beta and microRNA-146a in an immature rat model and children with mesial temporal lobe epilepsy. Epilepsia 53:1215–1224
Omran A, Ashhab MU, Gan N, Kong H, Peng J, Yin F (2013) Effects of MRP8, LPS, and lenalidomide on the expressions of TNF-alpha, brain-enriched, and inflammation-related microRNAs in the primary astrocyte culture. ScientificWorldJournal 2013:208309
O'Neill LA (2009) Boosting the brain's ability to block inflammation via microRNA-132. Immunity 31:854–855
Parker NR, Correia N, Crossley B, Buckland ME, Howell VM, Wheeler HR (2013) Correlation of microRNA 132 up-regulation with an unfavorable clinical outcome in patients with primary glioblastoma multiforme treated with radiotherapy plus concomitant and adjuvant temozolomide chemotherapy. Transl Oncol 6:742–748
Pedersen IM, Otero D, Kao E et al (2009) Onco-miR-155 targets SHIP1 to promote TNFalpha- dependent growth of B cell lymphomas. EMBO Mol Med 1:288–295
Peng J, Omran A, Ashhab MU et al (2013) Expression patterns of miR-124, miR-134, miR-132, and miR-21 in an immature rat model and children with mesial temporal lobe epilepsy. J Mol Neurosci 50:291–297
Quinn SR, O’Neill LA (2011) A trio of microRNAs that control Toll-like receptor signalling. Int Immunol 23:421–425
Scott HL, Tamagnini F, Narduzzo KE et al (2012) MicroRNA-132 regulates recognition memory and synaptic plasticity in the perirhinal cortex. Eur J Neurosci 36:2941–2948
Seifert G, Schilling K, Steinhäuser C (2006) Astrocyte dysfunction in neurological disorders: a molecular perspective. Nat Rev Neurosci 7:194–206
Selbach M, Schwanhausser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N (2008) Widespread changes in protein synthesis induced by microRNAs. Nature 455:58–63
Shaked I, Meerson A, Wolf Y et al (2009) MicroRNA-132 potentiates cholinergic anti-inflammatory signaling by targeting acetylcholinesterase. Immunity 31:965–973
Shaltiel G, Hanan M, Wolf Y et al (2013) Hippocampal microRNA-132 mediates stress-inducible cognitive deficits through its acetylcholinesterase target. Brain Struct Funct 218:59–72
Sheng JG, Mrak RE, Griffin WS (1994) S100 beta protein expression in Alzheimer disease: potential role in the pathogenesis of neuritic plaques. J Neurosci Res 39:398–404
Shimada T, Takemiya T, Sugiura H, Yamagata K (2014) Role of inflammatory mediators in the pathogenesis of epilepsy. Mediat Inflamm 2014:901902
Sklan EH, Lowenthal A, Korner M et al (2004) Acetylcholinesterase/paraoxonase genotype and expression predict anxiety scores in Health, Risk Factors, Exercise Training, and Genetics study. Proc Natl Acad Sci U S A 101:5512–5517
Sofroniew MV (2009) Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci 32:638–647
Soreq H, Wolf Y (2011) NeurimmiRs: microRNAs in the neuroimmune interface. Trends Mol Med 17:548–555
Sternfeld M, Shoham S, Klein O et al (2000) Excess “read-through” acetylcholinesterase attenuates but the “synaptic” variant intensifies neurodeterioration correlates. Proc Natl Acad Sci U S A 97:8647–8652
Taganov KD, Boldin MP, Chang KJ, Baltimore D (2006) NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A 103:12481–12486
Tili E, Michaille JJ, Cimino A et al (2007) Modulation of miR-155 and miR-125b levels following lipopolysaccharide/TNF-alpha stimulation and their possible roles in regulating the response to endotoxin shock. J Immunol 179:5082–5089
Tognini P, Pizzorusso T (2012) MicroRNA212/132 family: molecular transducer of neuronal function and plasticity. Int J Biochem Cell Biol 44:6–10
Tracey KJ (2007) Physiology and immunology of the cholinergic antiinflammatory pathway. J Clin Invest 117:289–296
Vainas T, Stassen FR, Bruggeman CA et al (2006) Synergistic effect of Toll-like receptor 4 and CD14 polymorphisms on the total atherosclerosis burden in patients with peripheral arterial disease. J Vasc Surg 44:326–332
Vezzani A, Balosso S, Ravizza T (2008) The role of cytokines in the pathophysiology of epilepsy. Brain Behav Immun 22:797–803
Vezzani A, Aronica E, Mazarati A, Pittman QJ (2013) Epilepsy and brain inflammation. Exp Neurol 244:11–21
Viemann D, Barczyk K, Vogl T et al (2007) MRP8/MRP14 impairs endothelial integrity and induces a caspase-dependent and -independent cell death program. Blood 109:2453–2460
Virtue A, Wang H, Yang XF (2012) MicroRNAs and toll-like receptor/interleukin-1 receptor signaling. J Hematol Oncol 5:66
Vogl T, Ludwig S, Goebeler M et al (2004) MRP8 and MRP14 control microtubule reorganization during transendothelial migration of phagocytes. Blood 104:4260–4268
Vogl T, Tenbrock K, Ludwig S et al (2007) Mrp8 and Mrp14 are endogenous activators of Toll-like receptor 4, promoting lethal, endotoxin-induced shock. Nat Med 13:1042–1049
Xiao J, Li Y, Prandovszky E et al (2014) MicroRNA-132 dysregulation in Toxoplasma gondii infection has implications for dopamine signaling pathway. Neuroscience 268:128–138
Yonekawa K, Neidhart M, Altwegg LA et al (2011) Myeloid related proteins activate Toll-like receptor 4 in human acute coronary syndromes. Atherosclerosis 218:486–492
Ziegler G, Prinz V, Albrecht MW et al (2009) Mrp-8 and −14 mediate CNS injury in focal cerebral ischemia. Biochim Biophys Acta 1792:1198–1204
Acknowledgments
This work was kindly supported by the National Natural Science Foundation of China (Nos. 81371434, 81370771, 81301031) and Hunan Provincial Innovation Foundation for Postgraduate (No. 2501–71380100017). The authors thank Dr. Chao Chen for revising the manuscript. We also thank all members of the laboratory for insightful discussions.
Conflict of Interest
The authors declare that there are no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kong, H., Yin, F., He, F. et al. The Effect of miR-132, miR-146a, and miR-155 on MRP8/TLR4-Induced Astrocyte-Related Inflammation. J Mol Neurosci 57, 28–37 (2015). https://doi.org/10.1007/s12031-015-0574-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12031-015-0574-x