
Abstract
Voltage-gated N-type Ca2+ channels play a central role in regulation of neurotransmitter release. The II–III linker of N-type calcium channel α1B subunit (exons 17–21 in rats) contains synaptic protein interaction (synprint) site, which plays a pivotal role in efficient neurotransmission. Using RT-PCR analysis of rat hippocampus and lumbar spinal cord transcripts, the expressions of Cav2.2 exon 18a and a novel Cav2.2 splice variant, which were detected in this study, have been investigated, and they showed tissue-specific pattern. The new variant contains a large deletion in II–III linker that produces a stop codon, which is predicted to produce a two-domain-truncated channel. To study the effect of inflammatory pain on the expression pattern of these variants, animals were treated with intraplantar formalin, and the amount of splice variants in lumbar spinal cord was measured. The results showed that chronic inflammatory pain increases the Cav2.2 mRNA levels lacking exon 18a and decreases the amount of full-length variants, which do not have any deletion. Determining different Cav2.2 splice variants in rat nervous system and the impact of inflammatory pain on the splicing pattern suggest a possible regulatory role for calcium channel alternative splicing.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Voltage-gated calcium channels mediate rapid and voltage-dependent entry of calcium into many types of neural, muscle, and endocrine cells. Calcium entry contributes to a number of physiological functions, including neurotransmitter and hormone secretion, muscle contraction, and release of and modulation of other membrane ion channels (Reuter 1983; Tsien 1983).
Electrophysiological analysis categorizes native calcium currents into two major classes that differ in their voltage activation properties: high-voltage-activated calcium channels (L-, N-, P/Q- and R-types) and low-voltage-activated calcium channels (T-type; Hofmann et al. 1994). An individual channel is composed of multiple subunits (α1, β, α2/δ, γ). The α1 subunit, known as the main pore-forming subunit, determines the major functional properties of the channel (Dunlap et al. 1995). The pore-forming α1 subunit contains as a basic motif a tetrameric association of four domains each containing a series of six transmembrane α-helical segments, numbered S1–S6, which are connected by both intra- and extracellular loops (Dubel et al. 1992). Ten genes encoding Ca2+ channel α1 subunits have been cloned and classified systematically according to their characteristics (Ertel et al. 2000).
Splicing of α1B mRNA also represents an important mechanism for optimization of neurotransmitter release in different regions of the nervous system (Lin et al. 1997, 1999; Pan and Lipscombe 2000). Alternative splicing occurs most often in cytoplasmic domains of Cav2 channels, where the effects on channel gating are hard to predict (Thaler et al. 2004).
N-type Ca2+ channels are essential for transmission of nociception. These channels are highly concentrated in presynaptic terminals of small-diameter myelinated and unmyelinated nociceptors that synapse in laminae I and II of the dorsal horn where they control neurotransmitter release (Holz et al. 1988; Maggi et al. 1990). Because of the importance of N-type calcium channels in the release of pronociceptive neurotransmitters and pain transmission in spinal cord, they are attractive targets for therapeutic intervention concerning chronic and neuropathic pain conditions (Snutch 2005).
There are several reports about the importance of exon 37a/ exon 37b splice site in the activity of N-type calcium channels in nociceptors (Bell et al. 2004; Castiglioni et al. 2006; Altier et al. 2007). Here, we focus on the alternative splicing in the linker of domains II and III of Cav2.2 and its role in pain transmission. An important part of this linker is called synaptic protein interaction site (synprint site), which associates with proteins of the presynaptic vesicle docking and release machinery (Sheng et al. 1994, 1996; Catterall 1999). An exon cassette called exon 18a in the Cav2.2 gene encoding 21 amino acids in the synprint site preferentially expresses in monoamine neurons in the rat brain (Ghasemzadeh et al. 1999). The insertion of this exon cassette induces a shift in the steady-state inactivation curve to more depolarized potentials without affecting the kinetics or voltage dependence of channel activation (Coppola et al. 1994).
Kaneko and colleagues in 2002 reported two novel human Cav2.2 calcium channel variants (Cav2.2 Δ1and Cav2.2 Δ2). These two splice variants contain large sequence deletions in the linker of domains II and III, including the synprint site. The voltage dependence of steady-state inactivation of these variants was shifted to more depolarized potentials, and recovery from inactivation occurred more rapidly than that of the full-length channel.
In the present study, the expression pattern of Cav2.2 synprint site splice variants in rat hippocampus and lumbar spinal cord is described. A novel splice variant, which contains a stop codon as a result of the deletion of large portions of Cav2.2 cytoplasmic II–III linker, was detected. Besides, the impact of chronic inflammatory pain on the alternative splicing of Cav2.2 II–III linker in lumbar spinal cord is reported.
Materials and Methods
Animals
In this study, male Wistar rats weighing 200–250 g were used. Animals were housed in groups of four and had free access to food and drinking water with a 12-h light/dark cycle. The injections were performed between 10:00 and 12:00 each day to reduce any variation between groups. Animal research protocols were approved by the Committee of Ethics in Research, Shahid Beheshti University (M.C.) and conformed to NIH guidelines for use of animals in research. All efforts were made to minimize animal suffering and to reduce the number of animals used.
Animals were grouped in four groups of rats (four to six in each). To induce chronic inflammatory pain, 50 μl of 2% formalin was injected subcutaneously (s.c.) into the hind paws (Javan et al. 2005). The first group received single-dose formalin in the dorsal region of right paw. In the second group, injections were repeated for 4 days in different regions of the paws (first day: right/dorsal; second day: left/ventral; third day: right/ventral; fourth day: left/dorsal). For the other two control groups (single dose and repeated one), saline was injected to hind paw with the same protocol for formalin injections.
Gene Expression Study
One hour after the single-dose injections (formalin or saline) and 24 h after the last injections in the second and fourth groups (repeated ones), animals were anesthetized using CO2 and then decapitated. Extracted samples of hippocampus and lumbar spinal cord were preserved in liquid nitrogen and then used for isolation of total cellular RNAs. The protocol was a modification of guanidine isothiocyanate–phenol–chloroform method using RNX+ reagent (CinnaGen, Iran). Samples were used immediately in reverse transcription reaction using oligo-dT primer and M-MuLV reverse transcriptase (Fermentas) based on the manufacturer’s protocol. The reactions were incubated at 42°C for 60 min and then inactivated at 70°C for 10 min.
To determine the possible position of Δ1 deletion (a 1,146-base-pair deletion between nucleotides 2412 and 3559 in humans, which corresponds to a 382-amino-acid deletion between Arg756 and Leu1139 in rats), two human (GeneBank accession number M94172) and rat (GeneBank accession number M92905) mRNAs were aligned, and the location of this deletion in the rats was guessed according to its site in humans. The Cav2.2 e18a and Cav2.2 Δ1 primers were designed flanking the regions of e18a insertion and Δ1 deletion, respectively. In addition, two other primers were used specifically for β-actin as an internal standard. The sequence of primers used was as follows: Cav2.2 e18a forward: 5′-GTCAGCAAAGGCATGTTTTCATC-3′; Cav2.2 e18a reverse: 5′-GTGGTGATTTCGAGTTTCCTTATC-3′; Cav2.2 Δ1 forward: 5′-GCCAATCAGAAG CTTGCTCTTC-3′; Cav2.2 Δ1 reverse: 5′-CGATGACCACAAGAATCACCATC-3′; β-actin forward: 5′-CCCAGAGCAAGAGAGGCATC-3′; β-actin reverse: 5′-CTCAGGAGGAGCAATGATCT-3′.
A semi-quantitative PCR method (Maron et al. 2002) was used for studying the expression of Cav2.2 splice variants in rat hippocampus and lumbar spinal cord. The reaction parameters and number of PCR cycles were adjusted to obtain a condition with linear relation between the number of PCR cycles and PCR products and with linear relation between the initial amount of cDNA template and PCR product. The PCR reactions were incubated at 95°C for 5 min, followed by 35 thermal cycles (60 s at 94°C, 60 s at 58°C, 60 s at 72°C) for Cav2.2 e18a and Cav2.2 Δ1 primers and 25 ones for β-actin primers. The final cycle was followed by a 10-min extension step at 72°C. Upon performing PCR reaction, PCR products were analyzed on 2% low melting point agarose (Roche, Germany) gel and bands were quantified by densitometry using Lab works analyzing software (UVP, UK).
To determine the Δ1 deletion position in rats, band was sequenced and compared with the NCBI genomic DNA reference sequence (NC-005120), mRNA reference sequence (NM-147141), and protein reference sequence (NP-671482) using Sequencher software (Gene Code Corporation, Ann Arbor, MI, USA).
Data Analysis
The values of Cav2.2 containing and lacking e18a along with Cav2.2 full length and deleted band densities and β-actin ones were obtained from gel analysis using Lab works software. Band densitometry was calculated and expressed as Cav2.2 18a/β-actin and Cav2.2 Δ1/ β-actin, respectively. Data have been presented as mean ± SEM. The averages of different groups were compared using one-way ANOVA followed by Tukey’s post test to check statistical significance. P < 0.05 was considered as significant difference.
Results
Determination of Splice Variants in Cav2.2 Synprint Site
After performing PCR using specific primers for detection of e18a insertion and Δ1 deletion, two amplified fragments in each reaction were separated by gel electrophoresis.
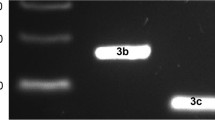
As is shown in Fig. 1a, two 343 and 406 bp bands were observed in the PCR products of Cav2.2 e18a primers. In the PCR products of Cav2.2 Δ1 primers, two different bands were detected and showed a novel deletion in this site. The bands separated by gel electrophoresis were 1,232 and 278 bp in size (Fig. 1b).

Gel electrophoresis pattern of PCR products for two different alternative splicing. a PCR products of Cav2.2 e18a primers. Lane 1 = 100 bp DNA ladder, lane 2 = Cav2.2 containing (406 bp) and lacking (343 bp) e18a variants in the intact rats. b PCR products of Cav2.2 Δ1 primers. Lane 1 = 100 bp DNA ladder, lane 2 = Cav2.2 full length (1,232 bp) and Δ1 deleted (278 bp) variants in the intact rats
The Δ1 deletion band detected in gel electrophoresis was different in size with what we expected according to the Δ1 deletion previously reported in human (Kaneko et al. 2002). Sequencing of Δ1 deletion band determined the deletion site as shown in Fig. 2.

The sequence of Δ1 deletion in the rat synprint site. Δ1 deletion occurs after the Cc.2270 and Gc.3281, and a 21-kb intron is spliced out. As a result, a stop codon arises, which was predicted to produce a truncated protein with 758 amino acids
The sequence of deleted variant showed a stop codon in the synprint site as a result of Δ1 deletion, which is predicted to produce a truncated protein with only 758 amino acids.
Tissue-Specific Expression Pattern of Cav2.2 Splice Variants
As shown in Fig. 3, there are significant differences between expression pattern of 18a exon cassette in hippocampus and lumbar spinal cord. In hippocampus, most of the α1B subunits lack e18a, but in lumbar spinal cord, approximately half of the channels contain this exon encoding 21 amino acids. The expression pattern of Δ1 deletion is also different in these two regions. Cav2.2 full-length variants without Δ1 deletion in the lumbar spinal cord are more abundant than hippocampus. In other words, deletion occurred in few Cav2.2 variants in the lumbar spinal cord, but it happened in hippocampus more frequently. As can be observed, the β-actin band densities did not show a significant difference in these two regions.

Comparative expression pattern of different Cav2.2 splice variants in rat hippocampus and lumbar spinal cord. a Gel electrophoresis of β-actin, Cav2.2 e18a, and Cav2.2 Δ1 PCR products. Lane 1 = 100 bp DNA ladder, lane 2 = β-actin in hippocampus, lane 3 = β-actin in lumbar spinal cord, lane 4 = Cav2.2 e18a containing (406 bp) and lacking (343 bp) variants in hippocampus, lane 5 = Cav2.2 e18a containing and lacking variants in lumbar spinal cord, lane 6 = Cav2.2 full length (1,232 bp) and Δ1 deletion (278 bp) variants in hippocampus, lane 7 = Cav2.2 full length and Δ1 deletion variants in lumbar spinal cord. b Quantitative comparison of β-actin, Cav2.2 e18a, and Cav2.2 Δ1 PCR products’ band densities in hippocampus and lumbar spinal cord. Data have been shown as mean ± SEM; *P < 0.05
Effects of Inflammatory Pain on the Expression Pattern of Cav2.2 Splice Variants in Lumbar Spinal Cord
Following formalin-induced chronic inflammatory pain, some changes were detected in the expression patterns of Cav2.2 splice variants. Although β-actin band density did not change after saline or formalin treatment (Fig. 4a), the band density of e18a lacking variant increased after single-dose administration of formalin (P < 0.05; Fig. 4b). The amount of Cav2.2 e18a containing variant was unchanged after the single-dose or chronic administration of formalin.

Effects of chronic inflammatory pain on Cav2.2 alternative splicing in rat lumbar spinal cord. a Effects of chronic inflammatory pain on β-actin expression. As it can be observed, no significant changes occur after saline or formalin treatment. b Effects of chronic inflammatory pain on the Cav2.2 e18a alternative splicing. The Cav2.2 e18a lacking variants levels increase after single-dose injection of formalin. No changes can be detected in the other situations. c Effects of chronic inflammatory pain on the Cav2.2 Δ1 deletion alternative splicing. The Cav2.2 full-length variants levels decrease both after single-dose and chronic administration of formalin but not after saline injections. N normal, AS acute saline, AF acute formalin, CS chronic saline, CF chronic formalin. Data have been shown as mean ± SEM; *P < 0.05 when compared with normal rats
As shown in Fig. 4c, both single-dose and chronic administration of formalin reduced the amount of full-length variant without Δ1 deletion but did not have any effect on Δ1 deletion variant (P < 0.05).
Discussion
The consequences of regulated alternative splicing among the genes that comprise voltage-gated calcium channels permits specialization of channel function, optimizing calcium signaling in different regions of the brain and in different cellular compartments (Lipscombe et al. 2002). Recent studies focus on N-type Ca2+ channels because of their importance in control of neurotransmitter release in different types of neurons.
In mammalian neurons, the synprint site, located within domain II–III linker of Cav2.2 subunits, associates with proteins of presynaptic vesicle docking and release machinery such as syntaxin1, SNAP25, Synaptotagmin 1, and CSP (Catterall 1999). The focus of this study was on exon cassette 18a encodes 21 aa located between exons 18 and 19 in LII–III of Cav2.2 genes, and the Δ1 deletion occurs at this site. Besides the expression pattern of these variants in hippocampus and lumbar spinal cord, the effects of chronic inflammatory pain on their expression levels have been investigated.
Novel Cav2.2 Splice Variant in Rats with a Stop Codon in Synprint Site
To study whether the synprint site deletion reported previously in humans (Kaneko et al. 2002) also exists in rats, human and rat cDNAs were aligned, and the possible region of Δ1 deletion was marked in the rat cDNA. The deleted band, which was amplified by the PCR, was sequenced and showed a novel deletion in rat synprint site (Fig. 2).
As is shown in Fig. 5a, as a result of Δ1 deletion, a large portion of Cav2.2 II–III linker is deleted, which is predicted to produce a truncated protein in rats. This deletion is similar to one previously reported in human by Kaneko et al. (2002). Full-length II–III linker region of Cav2.2 variants is encoded by six exon cassettes in humans (17 to 22) and five in rats (17 to 21; Fig. 5b). In the case of Δ1 deletion in rats, an intron with GT-AG sites (nucleotides between Cc.2270 and Gc.3281) is spliced out and deletes a large part of synprint site (exon 19 and 20). Interestingly a stop codon arises as a result of this deletion, which is predicted to produce a truncated protein with 758 amino acids containing two domain (the complete α1B subunit contains 2,336 amino acids in rats). To the best of our knowledge, there is just one report (published in abstract form) of N-type calcium channel splice variant, which has a stop codon in the middle of the structure (Mittman and Agnew 2000). So, it will be very interesting to detect the protein that resulted from Δ1 deletion variants in different region of CNS and investigate whether such a structure, which is similar to half part of the complete α1B subunit, is functional or not.

Alternative splicing in the cytoplasmic linker of domains II and III of rat Cav2.2. a An alternative splicing in II–III linker of Cav2.2, which produces a full-length variant; a1 N-type calcium channel alpha 1B subunit (Cav2.2) gene structure; a2 schematic diagram of the gene region encoding for II–III linker. Full-length II–III linker is encoded by exons 17 to 21; a3 mRNA sequence corresponding to full-length II–III linker of Cav2.2; a4 schematic structure of rat N-type calcium channel with full-length II–III linker showing the parts encoded by exons 17 to 21. b Alternative splicing in II–III linker of Cav2.2 that deletes a large part of mRNA encoding for II–III linker and leads to an expected two-domain calcium channel; b1 schematic diagram of the gene region encoding for II–III linker; b2 mRNA sequence corresponding to II–III linker, which is produced as a result of Δ1 deletion. Exons 19 and 20 with the flanking introns are spliced out, and a stop codon arises; b3 two-domain calcium channel structure, which is expected to be produced as a result of Δ1 deletion in rats. c Genomic DNA sequence at boundary sites of Cav2.2 full length and deleted variants. In the case of Δ1 deletion, a large intron (29 kb) with consensus gt-ag terminals is spliced out. As a result, a stop codon arises, which is expected to produce a truncated protein with 758 amino acids. IS1 First α-helical segment of domain I, IS6 sixth α-helical segment of domain I, IIS1 first α-helical segment of domain II, IIS6 sixth α-helical segment of domain II, IIIS1 first α-helical segment of domain III, IIIS6 sixth α-helical segment of domain III, IVS1 first α-helical segment of domain IV, IVS6 sixth α-helical segment of domain IV
There are some reports about recombinant truncated voltage-gated Ca2+ channels. Ahern et al. (2001) cut the four-domain DHPR α1S in the cytoplasmic linker between domain II and III and expressed fragments separately. Although excitation–contraction was not coupled, charge movement was detected in the I–II domain that is expressed alone. A two-domain I–II construct of neuronal α1A Ca2+ channels also recovered charge movements like α1S I–II domain. Moreover, they characterized the recombinant two-domain form (α1A I–II) and showed that co-expression of the α1A I–II significantly reduces the current amplitude of α1A/β1a/α2δ channel via competition for β subunit (Arikkath et al. 2002). Similar results about α1A were reported by Raike et al. (2007). Interestingly, it has been demonstrated that a truncated form of α1B protein, similar in structure to the α1A I–II, inhibits the current generated by the full-length α1B protein (Raghib et al. 2001). Suppression requires interaction between a truncated construct and a related full-length channel, which activates a component of the unfolded protein response to suppress translation (Page et al. 2004). However, little is known about the tissue specificity and expression regulation of these truncated variants.
Region-Specific Expression Pattern of Cav2.2 Splice Variants
Alternative splicing in cytoplasmic II–III loop of the α1B mRNAs is differentially regulated in hippocampus and lumbar spinal cord. As it has been shown in Fig. 3, β-actin expresses in these two regions equally, but there are some differences in the Cav2.2 splice variants levels. In the hippocampus, most of the channels lack e18a, and exon skipping is a frequent event in this region, while approximately half of the channels contain this exon in lumbar spinal cord. Because of the physiological, biophysical, and pharmacological differences of e18a containing and e18a lacking variants (Pan and Lipscombe 2000; Rott and Horn 2003; Thaler et al. 2004), these variations can cause tissue specificity through the nervous system.
Δ1 deletion variants levels are similar in hippocampus and lumbar spinal cord, but full-length variants are more abundant in lumbar spinal cord in comparison with hippocampus. If the expected Δ1 deletion variant produces a truncated protein, which has negative effect on full-length channel synthesis (Raghib et al. 2001; Page et al. 2004), more full-length variants in lumbar spinal cord means more functional channels in this region. These data suggest that regional regulation of Cav2.2 alternative splicing can cause structural and functional diversity in CNS, and it will be a good target for designing new drugs, which affect specific region of CNS with attenuated side effects on the other parts.
Alternative Splicing Modulation by Chronic Inflammatory Pain
Generally speaking, little is known about the mechanisms which regulate expression of alternative spliced exons of α1B in different regions of nervous system. To study the impact of chronic inflammatory pain on Cav2.2 alternative splicing, formalin pain was selected as a commonly used model of persistent pain (Dubuisson and Dennis 1977; Abbott et al. 1995; Damas and Liegeois 1999; Fu et al. 2001), and lumbar spinal cord was extracted from treated animals as an important site in pain transmission to analyze the changes in Cav2.2 mRNA levels. Besides, lumbar spinal cord contains N-type channels in its dorsal horn terminals associated with medium-sized Aδ fibers and small-diameter C-type fibers, both of which are responsible for pain perception (Gribkoff 2006). There were several reports about behavioral and histological consequences of formalin as pain inducer (Matthias and Franklin 1992; Damas and Liegeois 1999; Fu et al. 2001) and its mechanism of function (Lee et al. 2008), so we just focused on molecular events that occurred after chronic inflammatory pain induction.
As is shown in Fig. 4, several changes were observed in the expression pattern of e18a variants and Δ1 deletion variants in formalin-treated animals, while β-actin band densities, which were used as an internal standard, did not change following formalin administration (Fig. 4a).
Single-dose formalin injection caused an increase in Cav2.2 mRNA levels lacking exon 18a but no changes in the level of Cav2.2 e18a containing variants. According to the electrophysiological differences between these two variants (Pan and Lipscombe 2000; Thaler et al. 2004), this alteration may lead to channels with attenuated excitability and fewer exocytosis events. The other alterations that have been investigated were reduction in the full-length variant levels, both after single and repeated injections of formalin (Fig. 4c). These alterations may be the result of change in splicing or overall decrease in Cav2.2 mRNA levels, but in both cases, they cause reduction in the amount of functional channels necessary for neurotransmitter release. It will be useful to investigate changes in other important sites for pain transmission and perception such as dorsal root ganglia to determine the exact impact of chronic inflammatory pain on N-type calcium channels. Expanding our knowledge about the molecular effects of chronic inflammatory pain in nervous system can help us to design new drugs to control the pain.
References
Abbott, F. V., Franklin, K. B. J., & Westbrook, R. F. (1995). The formalin test: scoring properties of the first and second phases of the pain response in rats. Pain, 65, 91–102. doi:10.1016/0304-3959(94)00095-V.
Ahern, C. A., Arikkath, J., Vallejo, P., Gurnett, C. A., Powers, P. A., Campbell, K. P., et al. (2001). Intramembrane charge movements and excitation-contraction coupling expressed by two-domain fragments of the Ca2+ channels. Proceedings of the National Academy of Sciences of the United States of America, 98, 6935–6940. doi:10.1073/pnas.111001898.
Altier, C., Dale, C. S., Kisilevsky, A. E., Chapman, K., Castiglioni, A. J., Matthews, E. A., et al. (2007). Differential role of the N-type calcium channel splice isoforms in pain. The Journal of Neuroscience, 27, 6363–6373. doi:10.1523/JNEUROSCI.0307-07.2007.
Arikkath, J., Felix, R., Ahern, C., Chen, C. C., Mori, Y., Song, I., et al. (2002). Molecular characterization of a two-domain form of the neuronal voltage-gated P/Q-type calcium channel α12.1 subunit. FEBS Letters, 532, 300–308. doi:10.1016/S0014-5793(02)03693-1.
Bell, J. T., Thaler, C., Castiglioni, A. J., Helton, T. D., & Lipscombe, D. (2004). Cell-specific alternative splicing increases calcium channel current density in the pain pathway. Neuron, 41, 127–138. doi:10.1016/S0896-6273(03)00801-8.
Castiglioni, A. J., Raingo, J., & Lipscombe, D. (2006). Alternative splicing in the C-terminus of Cav2.2 controls expression and gating of N-type calcium channels. The Journal of Physiology, 576(1), 119–134. doi:10.1113/jphysiol.2006.115030.
Catterall, W. A. (1999). Interactions of presynaptic Ca2+ channels and snare proteinsin neurotransmitter release. Annals of the New York Academy of Sciences, 868, 144–159. doi:10.1111/j.1749-6632.1999.tb11284.x.
Coppola, T., Waldmann, R., Borsotto, M., Hurteaux, C., Romy, G., & Lazdunski, M. (1994). Molecular cloning of a murine N-type calcium channel α1 subunit evidence for isoforms, brain distribution, and chromosomal localization. FEBS Letters, 338, 1–5. doi:10.1016/0014-5793(94)80105-3.
Damas, J., & Liegeois, J. F. (1999). The inflammatory reaction induced by formalin in the rat paw. Naunyn-Schmiedeberg’s Archives of Pharmacology, 359, 220–227. doi:10.1007/PL00005345.
Dubel, S. J., Starr, T. V. B., Hell, J., Ahluanian, M. K., Enyeart, J. J., Catterall, W. A., et al. (1992). Molecular cloning of the α-1 subunit of an ω-conotoxin-sensitive calcium channel. Proceedings of the National Academy of Sciences of the United States of America, 89, 5058–5062. doi:10.1073/pnas.89.11.5058.
Dubuisson, D., & Dennis, S. G. (1977). The formalin test: a quantitative study to the analgesic effects of the morphine, meperidine and brain stem stimulation in rats and cats. Pain, 4, 161–174. doi:10.1016/0304-3959(77)90130-0.
Dunlap, K., Luebke, J. I., & Turner, T. J. (1995). Exocytotic Ca2+ channels in mammalian central neurons. Trends in Neurosciences, 18, 89–98. doi:10.1016/0166-2236(95)93882-X.
Ertel, E. A., Campbell, K. P., Harpold, M. M., Hofmann, F., Mori, Y., Perez-Reyes, E., et al. (2000). Nomenclature of voltage-gated calcium channels. Neuron, 25, 533–535. doi:10.1016/S0896-6273(00)81057-0.
Fu, K. Y., Light, A. R., & Maixner, W. (2001). Long lasting inflammation and long-term hyperalgesia after subcutaneous formalin injection into the rat hindpaw. Pain, 2, 2–11. doi:10.1054/jpai.2001.9804.
Ghasemzadeh, M. B., Pierce, R. C., & Kalivas, P. W. (1999). The monoamine neurons of the rat brain preferentially express a splice variant of α1B subunit of the N-type calcium channel. Journal of Neurochemistry, 73, 1718–1723. doi:10.1046/j.1471-4159.1999.731718.x.
Gribkoff, V. K. (2006). The role of voltage-gated calcium channels in pain and nociception. Seminars in Cell & Developmental Biology, 17, 555–564. doi:10.1016/j.semcdb.2006.09.002.
Hofmann, F., Biel, M., & Flockerzi, V. (1994). Molecular basis for Ca2+ channel diversity. Annual Review of Neuroscience, 17, 399–418. doi:10.1146/annurev.ne.17.030194.002151.
Holz, G. G. T., Dunlap, K., & Kream, R. M. (1988). Characterization of the electrically evoked release of substance P from dorsal root ganglion neurons: methods and dihydropyridine sensitivity. The Journal of Neuroscience, 8, 463–471.
Javan, M., Ahmadiani, A., Motamedi, F., & Kazemi, B. (2005). Changes in G proteins genes expression in rat lumbar spinal cord support the inhibitory effect of chronic pain on the development of tolerance to morphine analgesia. Neuroscience Research, 53, 250–256. doi:10.1016/j.neures.2005.06.020.
Kaneko, S., Cooper, C. B., Nshioko, N., Yamasaki, H., Suzuki, A., Jarvis, S. E., et al. (2002). Identification and characterization of novel human Cav2.2 (α1B) calcium channel variants lacking the synaptic protein interaction site. The Journal of Neuroscience, 22, 82–92.
Lee, A. T. H., Shah, J. J., Li, L., Cheng, Y., Moore, P. K., & Khanna, S. (2008). A nociceptive-intensity-dependent role for hydrogen sulphide in the formalin model of persistent inflammatory pain. Neuroscience, 152, 89–96. doi:10.1016/j.neuroscience.2007.11.052.
Lin, Z., Haus, S., Edgerton, J., & Lipscombe, D. (1997). Identification of functionally distinct isoforms of the N-type Ca2+ channel in rat sympathetic ganglia and brain. Neuron, 18, 153–166. doi:10.1016/S0896-6273(01)80054-4.
Lin, Z., Lin, Y., Schorge, S., Pan, J. Q., Beierlein, M., & Lopscombe, D. (1999). Alternative splicing of a short cassette exon in α1B generates functionally distinct N-type calcium channels in central and peripheral neurons. The Journal of Neuroscience, 19, 5322–5331.
Lipscombe, D., Pan, J. Q., & Gray, A. C. (2002). Functional diversity in neuronal voltage-gated calcium channels by alternative splicing of Cavα1. Molecular Neurobiology, 26, 21–44. doi:10.1385/MN:26:1:021.
Maggi, C. A., Tramontana, M., Cecconi, R., & Santicioli, P. (1990). Neurochemical evidence for the involvement of N-type calcium channels in transmitter secretion from peripheral endings of sensory nerves in guinea pigs. Neuroscience Letters, 114, 203–206. doi:10.1016/0304-3940(90)90072-H.
Maron, M., Mozzetti, S., Ritis, D. D., Pirelli, L., & Scambia, G. (2002). Semiquantitative RT-PCR analysis to assess the expression levels of multiple transcripts from the same sample. Biological Procedures Online, 3, 19–25. doi:10.1251/bpo20.
Matthias, B. K., & Franklin, K. B. J. (1992). Formalin pain is expressed in decerebrate rats but not attenuated by morphine. Pain, 51, 199–206. doi:10.1016/0304-3959(92)90261-9.
Mittman, S., & Agnew, W. S. (2000). Three new alternative spliced exons of the calcium channel 1B subunit gene CACNA1B. Soc. Neurosci. Abstr. 26, 40.4.
Page, M. K., Heblich, F., Davies, A., Butcher, A. J., Leroy, J., Bertaso, F., et al. (2004). Dominant-negative calcium channel suppression by truncated constructs involves a kinase implicated in the unfolded protein response. The Journal of Neuroscience, 24, 5400–5409. doi:10.1523/JNEUROSCI.0553-04.2004.
Pan, J. Q., & Lipscombe, D. (2000). Alternative splicing in the cytoplasmic II–III loop of the N-type Ca channel α1B subunit: functional differences are β subunit-specific. The Journal of Neuroscience, 20, 4769–4775.
Raghib, A., Bartaso, F., Davies, A., Page, K. M., Meir, A., Bogdanov, Y., et al. (2001). Dominant-negative synthesis suppression of voltage-gated calcium channel Cav2.2 induced by truncated constructs. The Journal of Neuroscience, 21, 8495–8504.
Raike, R. S., Kordasiewicz, H. B., Thompson, R. M., & Gomez, C. M. (2007). Dominant-negative suppression of Cav2.1 currents by α12.1 truncations requires the conserved interaction domain for β subunits. Molecular and Cellular Neurosciences, 34, 168–177. doi:10.1016/j.mcn.2006.10.011.
Reuter, H. (1983). Calcium channel modulation by neurotransmitters, enzymes and drugs. Nature, 301, 569–574. doi:10.1038/301569a0.
Rott, K. J., & Horn, F. L. (2003). The impact of splice isoforms on voltage-gated calcium channel α1 subunits. The Journal of Physiology, 554(3), 609–613. doi:10.1113/jphysiol.2003.052712.
Sheng, Z. H., Rettig, J., Takahashi, M., & Catterall, W. A. (1994). Identification of a syntaxin-binding site on N-type calcium channels. Neuron, 13, 1303–1313. doi:10.1016/0896-6273(94)90417-0.
Sheng, Z. H., Rettig, J., Cook, T., & Catterall, W. A. (1996). Calcium dependent interaction of N-type calcium channels with the synaptic core complex. Nature, 379, 451–454. doi:10.1038/379451a0.
Snutch, T. P. (2005). Targeting chronic and neuropathic pain: the N-type calcium channel comes of age. Neurorx, 2, 662–670. doi:10.1602/neurorx.2.4.662.
Thaler, C., Gray, A. C., & Lipscombe, D. (2004). Cumulative inactivation of N-type Cav2.2 calcium channels modified by alternative splicing. Proceedings of the National Academy of Sciences of the United States of America, 101, 5675–5679. doi:10.1073/pnas.0303402101.
Tsien, R. W. (1983). Calcium channels in excitable cell membranes. Annual Review of Physiology, 45, 341–358. doi:10.1146/annurev.ph.45.030183.002013.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Funding: This research was funded by the Neuroscience Research Center, Shahid Beheshti University (M.C.), Tehran, Iran.
Rights and permissions
About this article
Cite this article
Asadi, S., Javan, M., Ahmadiani, A. et al. Alternative Splicing in the Synaptic Protein Interaction Site of Rat Cav2.2 (α1B) Calcium Channels: Changes Induced by Chronic Inflammatory Pain. J Mol Neurosci 39, 40–48 (2009). https://doi.org/10.1007/s12031-008-9159-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12031-008-9159-2