
Abstract
The immune system shapes synaptic transmission and plasticity in experimental autoimmune encephalomyelitis (EAE), the mouse model of multiple sclerosis (MS). These synaptic adaptations are believed to drive recovery of function after brain lesions, and also learning and memory deficits and excitotoxic neurodegeneration; whether inflammation influences synaptic plasticity in MS patients is less clear. In a cohort of 59 patients with MS, we found that continuous theta-burst transcranial magnetic stimulation did not induce the expected long-term depression (LTD)-like synaptic phenomenon, but caused persisting enhancement of brain cortical excitability. The amplitude of this long-term potentiation (LTP)-like synaptic phenomenon correlated with the concentration of the pro-inflammatory cytokine interleukin-1β (IL-1β) in the cerebrospinal fluid. In MS and EAE, the brain and spinal cord are typically enriched of CD3+ T lymphocyte infiltrates, which are, along with activated microglia and astroglia, a major cause of inflammation. Here, we found a correlation between the presence of infiltrating T lymphocytes in the hippocampus of EAE mice and synaptic plasticity alterations. We observed that T lymphocytes from EAE, but not from control mice, release IL-1β and promote LTP appearance over LTD, thereby mimicking the facilitated LTP induction observed in the cortex of MS patients. EAE-specific T lymphocytes were able to suppress GABAergic transmission in an IL-1β-dependent manner, providing a possible synaptic mechanism able to lower the threshold of LTP induction in MS brains. Moreover, in vivo blockade of IL-1β signaling resulted in inflammation and synaptopathy recovery in EAE hippocampus. These data provide novel insights into the pathophysiology of MS.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Changes in synaptic excitability underlie several crucial brain functions, including learning and memory, and adaptive and maladaptive plasticity in response to experience and disease processes (Nisticò et al. 2012). In experimental autoimmune encephalomyelitis (EAE), a mouse model of multiple sclerosis (MS), the immune system shapes synaptic transmission and plasticity, driving recovery of function after a brain lesion, but also learning and memory deficits and excitotoxic neurodegeneration (Barbizan et al. Barbizan and Oliveira 2010; Centonze et al. 2009; Jenkins et al. 2010; Kim do et al. 2012; Mandolesi et al. 2010; Mori et al. 2011). The immune-neuronal interaction involves the responsiveness of neurons and glia to inflammatory cytokines released by infiltrating lymphocytes and activated microglia, such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) (Di Filippo et al. 2013; Haji et al. 2012; Mandolesi et al. 2012; Musumeci et al. 2011; Nisticò et al. 2013; Rossi et al. 2010, 2012a, b; Ziehn et al. 2012).
MS is an inflammatory, demyelinating and neurodegenerative disease of white and gray matter, and a major cause of disability in young adults. Recent studies highlight the importance of inflammation-induced synaptic dysfunction since the very early phases of MS (Mori et al. 2011; Rossi et al. 2012a, b), suggesting that not only in EAE, but also in human disease inflammatory molecules secreted by autoreactive T lymphocytes or activated microglia interfere with the physiological mechanisms of neuronal communication.
Two main forms of long-lasting adaptations of neuronal transmission have been described: long-term potentiation (LTP) and long-term depression (LTD), which consist, respectively, in enduring enhancement or weakening of synaptic strength in response to repeated synaptic activation (Malenka and Bear 2004). Both LTP and LTD can be explored safely and non-invasively in humans by means of, respectively, intermittent (iTBS) or continuous theta-burst stimulations (cTBS), delivered through a transcranial magnetic stimulation (TMS) device and developed to mimic the physiological activity of hippocampal neurons during learning episodes (Di Lazzaro et al. 2005; Huang et al. 2005). We have recently shown that iTBS-induced LTP is abnormal in the acute inflammatory phases of MS, suggesting that an impairment of cortical synaptic plasticity plays a substantial role in cognitive dysfunction associated with MS (Mori et al. 2011). Cortical responses to cTBS in MS, however, are still unclear, as well as the potential role of proinflammatory cytokines and of infiltrating lymphocytes on synaptic plasticity in this disorder.
Here, we show that LTP appearance is favored over LTD in response to repetitive synaptic activation in MS brains, and that IL-1β and infiltrating T lymphocytes likely play a substantial role in this alteration.
Methods
The study involving human subjects was approved by the Ethics Committee of the University Hospital Tor Vergata, Rome (Italy). All experiments in mice were carried out in accordance with the Guide for the Care and Use of Laboratory Animals and the European Communities Council Directive of November 24, 1986 (86/609/EEC).
Subjects
A total of 59 central-southern Italian relapsing–remitting (RR) MS patients were included in this study (47 females and 12 males, aged 17–58 years). The diagnosis of MS, in the remitting (n = 32) or in the relapsing phase (n = 27), was established by clinical, laboratory and MRI parameters, and matched published criteria (Polman et al. 2011). Seventeen of these subjects (14 females and 3 males, aged 17–45 years) were hospitalized at the neurological clinic of the University Hospital Tor Vergata of Rome (Italy) and underwent, within 24 h of admittance, CSF withdrawal for diagnostic purposes. They were later diagnosed as suffering from RRMS. Corticosteroids or other MS-specific immunoactive therapies were initiated later if appropriate. In all instances, patients underwent detection of oligoclonal banding in the CSF (positive in 90 % of cases). Expanded Disability Status Scale (EDSS) scores of MS subjects were between 0 and 5.0.
The 17 control subjects were healthy volunteers. Controls (13 females, 4 males, age 20–45) were matched to cases on age, gender and ethnicity. MS specialist neurologists performed clinical assessment of MS patients.
Transcranial Magnetic Stimulation
Transcranial magnetic stimulation was performed in the 59 RRMS patients and in the 17 age- and gender-matched healthy controls. All patients were asymptomatic in the upper right limb. The primary motor cortex of the dominant hemisphere was stimulated in all experiments. A monophasic Magstim 2002 connected to a figure of eight, 70-mm diameter coil, was used to define the motor hot spot and to assess MEP size. The motor hot spot was defined as the location where TMS consistently produced the largest MEP size in the target muscle. A second coil was connected to a biphasic stimulator, a Rapid2 Magstim package (Magstim Co., UK), and was used to deliver rTMS. The coils were held tangential to the scalp, at an angle of 45° away from the mid-sagittal line with the handle pointing backwards. According to the guidelines of the International Federation of Clinical Neurophysiology (Rothwell et al. 1999), we defined the resting motor threshold (RMT) as the minimum stimulation intensity over the motor hot spot, which can elicit an MEP of no <50 μV in 5 out of 10 trials. Active threshold (AMT) was defined as the intensity necessary to evoke a 200-μV, peak-to-peak amplitude, MEP while subjects maintained approximately 10 % contraction of the target muscle. The change in corticospinal excitability produced by each intervention was assessed by measuring the amplitude of the MEP response to a standard test pulse that remained constant throughout the experiment. In each subject, the intensity of this pulse was individually adjusted at the start of the experiment to produce a stable MEP (of 0.5–1 mV) with the subject at rest.
During the experiments, subjects were sitting comfortably in an armchair with their eyes open. EMGs were recorded via Ag/AgCl electrodes placed over the FDI of the dominant hand using a belly-tendon montage. Signals were filtered (30 Hz–2 kHz) and amplified (Digitimer 360, Digitimer Ltd., Welwyn Garden City, Herts, UK) and then stored on computer via a Power 1401 data acquisition interface (Cambridge Electronic Design Ltd., Cambridge, UK). Analysis was carried out using Signal Software (Cambridge Electronic Design). cTBS was applied over the motor cortex hot spot as described by Huang et al. (2005). Each burst consisted of 3 stimuli (80 % AMT) given at 50 Hz. cTBS, which usually suppresses corticospinal excitability, was delivered as a sequence of 200 bursts (600 stimuli) given at a rate of 5 Hz (total duration of 40 s). The effect of cTBS on corticospinal excitability was quantified by measuring the amplitude of MEPs evoked in the right first dorsal interosseous (FDI) by a constant intensity TMS pulse given over the contralateral motor cortex. Sixty MEPs were collected before cTBS (baseline) and at two different time points (0 and 15 min) after the end of cTBS. Stimulation intensity was set to induce a stable MEP of approximately 1 mV peak-to-peak amplitude in the relaxed right FDI at baseline and remained unchanged until end of recordings. MEP’s amplitudes were then averaged at each time point and normalized to the mean baseline amplitude.
Differences between groups for cTBS after-effects were evaluated using a repeated measures analysis of variance with between-subjects GROUP (MS vs. controls) and within-subjects TIME (baseline, 0 and 15 min after cTBS) main factors. Correlations between IL-1β CSF levels and cTBS effects on MEP size were analyzed through the Spearman correlation coefficient.
Cytokine Determination in the CSF
For measurements of inflammatory cytokines, the CSF was centrifuged and immediately stored at −80 °C until analyzed using Bio-Plex Multiplex Cytokine Assay (Bio-Rad Laboratories), according to manufacturer instructions. Concentrations of IL-1β and of TNFα (Bio-Rad Catalog #171-A11127, #171-305000) were calculated according to a standard curve generated for each target and expressed as pg/ml. When the concentrations of the cytokines were below the detection threshold, they were assumed to be 0 pg/ml.
EAE Induction
Experimental autoimmune encephalomyelitis was induced in 6–8 weeks old female C57BL/6 mice purchased from Charles-River (Italy). Mice were randomly assigned to standard cages, with four to five animals per cage, and kept under standard housing conditions with a light/dark cycle of 12 h and free access to food and water.
After 1 week of acclimatization, mice were injected subcutaneously at the flanks with 200 μg of myelin oligodendrocyte glycoprotein p35–55 (MOG35–55) emulsion for the induction of EAE by active immunization. The emulsion was prepared under sterile conditions using MOG35–55 (>85 % purity, Espikem, Florence, Italy) in complete Freund’s adjuvant (CFA, Difco), and Mycobacterium tuberculosis H37Ra (8 mg/ml; strain H37Ra, Difco, Lawrence, KS, USA) emulsified with phosphate buffered saline (PBS). The control emulsion was prepared in the same way without MOG35–55 for the control group (CFA group). All animals were injected with 500 ng of pertussis toxin (Sigma, St. Louis, MO, USA) intravenously on the day of immunization and 2 days later according to standard protocols of EAE induction. Animals were scored daily for clinical symptoms of EAE, according to the following scale: 0, no clinical signs; 1, flaccid tail; 2, hind limb weakness; 3, hind limb paresis; 4, complete bilateral hind limb paralysis; 5, death due to EAE; intermediate clinical signs were scored adding 0.5 value (Aktas and Zipp 2003; Centonze et al. 2009).
Preparation of CD3+ Cells from EAE Mice
After aseptically removing spleens from donor EAE and CFA mice (25 dpi), splenocyte suspensions were prepared by mechanically teasing the spleen with the blunt end of a 10-ml plastic syringe plunger in a Petri dish, then passing suspensions through a BD Falcon 70-μm cell strainer (BD Biosciences-Discovery Labware, Bedford, MA, USA) to remove debris and finally suspending the cells in 1 % fetal bovine serum, Roswell Park Memorial Institute 1640. From unfractionated splenocytes, CD3+ T cell lymphocytes were obtained by magnetic-activated cell sorting (MACS; Miltenyi Biotec, Bergisch Gladbach, Germany): mCD3 isolation kit. 5 × 103 pure T cells were then placed onto a single hippocampal slice and incubated for 30–60 min before the electrophysiological experiments.
Electrophysiology
Slices from mouse hippocampus (250–400 μm) were cut in ice-cold artificial cerebrospinal fluid (ACSF) composed of (in mM): NaCl (124), KCl (3.0), MgCl2 (1.0), CaCl2 (2.0), NaH2PO4 (1.25), NaHCO3 (26), glucose (10); saturated with 95 % O2, 5 % CO2, as previously described (Errico et al. 2011; Balducci et al. 2011). After an incubation period of at least 1 h slices were submerged in a recording chamber and perfused continuously with ACSF saturated with 95 % O2, 5 % CO2 at approximately 3 ml/min. Field excitatory post-synaptic potentials (fEPSP) were evoked with a bipolar electrode stimulating the Schaeffer collaterals. Extracellular recording electrodes filled with 3 M NaCl were placed in the stratum radiatum of CA1 region. The initial slope of the fEPSP was used as a measure of the evoked population excitatory synaptic response. A baseline was established with test pulses adjusted to evoke 30–40 % of the maximal response. LTP and LTD were induced by high-frequency stimulation (HFS; 100 Hz for 1 s) or paired-pulse low-frequency stimulation (PP-LFS, 900 pulses at 1 Hz), respectively.
Results are reported as mean ± SEM. Statistical significance was evaluated by Student’s t test between 50 and 60 min following delivery of the conditioning train. Statistical significance was set at p < 0.05. All experiments and the analysis of data were performed in a blind manner. For all statistical comparisons, the n used was the number of animals rather than number of slices.
Visually guided whole-cell recordings were obtained at room temperature (RT) from the soma of CA1 neurons using patch electrodes (3–4 MΩ) that contained (in mM): KCl (145), CaCl2 (0.05), EGTA (0.1), Hepes (10), Na3-GTP (0.3), Mg-GTP (4.0), pH adjusted to 7.3 with KOH. Unless otherwise stated, neurons were held at −70 mV. Current signals were filtered at 3 kHz and digitized at 10 kHz using a Multiclamp 700B operated by the pClamp10 software (Molecular Devices, Sunnyvale, CA, USA). No series resistance compensation was implemented, in order to keep a low signal-to-noise ratio, however, recordings were discarded if series resistance changed by more than 15 % from control.
The spontaneous events were detected from 3 min trace records and analyzed with Clampfit (Molecular Devices, Sunnyvale, CA, USA). Changes in amplitude or inter-event interval of the spontaneous events were compared according to their cumulative distributions, using the Kolmogorov–Smirnov (K–S) test, or their median values, using the Student’s t test.
Drugs were applied by dissolving them to the desired final concentration in the bathing solution and were as follows: CNQX (10 μM) from Tocris Cookson (Bristol, UK); IL-1 receptor antagonist (IL-1ra; 10 μg/ml) from R&D Systems (Minneapolis, USA).
Minipump Implantation and Continuous Intracranial Infusion of IL-1ra
One week before immunization, mice were implanted with a minipump in order to allow continuous intracerebroventricular (icv) infusion of either vehicle (n = 8) or IL-1ra (150 ng/die; R&D Systems) (n = 8) for 4 weeks. Alzet osmotic minipumps (model 1004; Durect Corporation, Cupertino, CA, USA) connected via catheter tube to intracranial cannula (Alzet Brain Infusion Kits 3) delivered vehicle or IL-1ra into the right lateral ventricle at a continuous rate of 0.11 μl/h. The coordinates used for icv minipump implantation were: antero–posterior = −0.4 mm from bregma; lateral = −1 mm; depth: 2.5 mm from the skull (Haji et al. 2012; Mandolesi et al. 2013).
Immunohistochemistry and Microscopy
Immunohistochemistry, microscopy and image analysis were performed as described previously (Nisticò et al. 2013). Mice were killed at the peak of the acute phase (20 dpi). They were deeply anesthetized with avertine and perfused through the aorta with ice-cold 4 % paraformaldehyde. Brains were post-fixed for at least 4 h at 4 °C and equilibrated with 30 % sucrose overnight (ON). Coronal sections (30 μm) were cut with a freezing microtome and permeabilized in PBS with Triton-X 0.25 % (TPBS). All following incubations were performed in TPBS.
Sections were pre-incubated with 10 % normal donkey serum solution for 1 h at RT and incubated with the following primary antibodies: rat anti CD3 (1:250, AbD Serotec); rabbit anti GFAP (1:500, DAKO); rabbit anti Iba1 (1:750, Wako). After being washed for three times (10 min each), sections were incubated with the following secondary antibodies: Cy3-conjugated donkey anti-rat (1:200, Jackson) or Alexa-488-conjugated donkey anti-rabbit (1:200; Invitrogen), for 2 h at RT and rinsed (last wash in Dapi 0.01 mg/ml). Serial sections were mounted with Vectashield (Vector Labs, USA) on poly-l-lysine-coated slides, air-dried and coverslipped. Images from immunolabeled samples were acquired with a Zeiss LSM700 confocal imaging system using 10× and 60× objectives (pixel resolution 1,024 × 1,024, digital zoom 0.5×). Section images in the z dimension (z spacing, 1 μm) were acquired with 60× objective. The confocal pinhole was kept at 1, the gain and the offset were adjusted to prevent saturation in the brightest signals. Subtraction of the background and smooth filter were applied on stacks to reduce noise by NIH ImageJ software (http://rsb.info.nih.gov/ij/). Each image stack was z projected and exported in TIFF file format.
Hippocampal Total Extract Preparations and Western Blot
Intracerebroventricular-implanted EAE (n = 5; 21 dpi), CFA (n = 4) and EAE-IL-1ra (n = 4) mice were sacrificed through cervical dislocation and the hippocampi were quickly removed and frozen until use. Tissues were homogenized in RIPA buffer plus protease inhibitor cocktail (Sigma). After sonication, the homogenates were centrifuged at 16,000×g for 15 min and supernatant collected. Protein content was quantified according to Bradford method. 10–20 μg of proteins were loaded onto 10 % polyacrylamide gel and then blotted onto nitrocellulose membrane (Protran, Whatman). This was blocked for 1 h at RT by 5 % non-fat dry milk in 0.1 % Tween20-PBS (T-PBS). All the following incubations were performed in T-PBS. Primary antibodies were used as following: rabbit anti-Iba1 (1:500, Wako, 1 over night) and rabbit anti synapsin (1:20,000, Novus Biologicals, 1 h RT). The mouse anti-beta-actin (1:20,000, Sigma-Aldrich, 1 h RT) or rabbit anti-GAPDH (1:20,000, cell signaling, 1 h RT) were used for normalization of protein contents. Membranes were incubated with secondary HRP-conjugated IgG (Millipore, 1:10,000) in 5 % milk for 1 h at RT. After washing, immunodetection was performed by ECL-Plus reagent (Amersham) and membrane was exposed to film (Sigma). Densitometric analysis of protein levels was performed by NIH ImageJ software (http://rsb.info.nih.gov/ij/).
Western blot (WB) results are presented as mean ± SEM and the data are normalized to control CFA values. Statistical analysis was performed using a paired or unpaired Student’s t test. Multiple comparisons were analyzed by one-way ANOVA followed by Tukey HSD. The significance level was established at p < 0.05.
Results
Aberrant Cortical Plasticity in MS
To study how inflammation impacts on cTBS-induced brain plasticity, 59 MS patients and 17 healthy subjects underwent the TMS procedure. Repeated measures ANOVA revealed a significant effect of group (F = 6.56; p < 0.05) and a significant time × group interaction (F = 3.82; p < 0.05). Post hoc contrasts showed that effects on MEP amplitude were remarkably different in the two experimental groups 15 min after cTBS (F = 6.40; p < 0.05), because this stimulation protocol caused the expected LTD-like effect in controls, and resulted in no effect or in frank long-lasting potentiation of MEP amplitude in MS subjects. Post hoc contrasts failed to reach significance at the 0.05 level immediately after cTBS (F = 3.48; p = 0.075), however, there was a similar trend (Fig. 1a).

Abnormal enhancement of cortical excitability in MS patients. a cTBS induced-effects differ between 59 MS patients and 17 healthy subjects (HS). The control group exhibits the expected LTD-like effect, while MS patients show LTP-like changes. Differences between the two groups were significantly different 15 min after cTBS. Correlation plots between b TNF-α and c IL-1β levels in the CSF of 17 MS subjects and the MEP amplitude changes, expressed as percentage of baseline 15 min after TBS. LTP-like changes in MS patients correlate with IL-1β but not with TNF-α CSF levels. *Indicates p < 0.05, ANOVA
To detail the role of central inflammation in the abnormal synaptic plasticity of MS subjects, correlations between CSF concentrations of two major pro-inflammatory cytokines, TNF-α and IL-1β and cTBS-induced MEP changes were explored in a subsample of 17 MS subjects undergoing CSF withdrawal for diagnostic purposes. No correlation was found between TNFα CSF levels and cTBS effect in these subjects (Fig. 1b), while IL-1β levels were significantly and directly correlated (ρ = 0.53; p < 0.05) with the cTBS-induced changes in MEP amplitude (Fig. 1c).
Infiltrating T Lymphocytes and Astrogliosis in the Hippocampus of EAE Mice
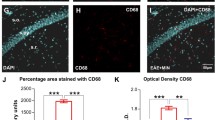
We have recently shown that LTP induction is favored over LTD also in the hippocampus of EAE mice, and this effect is associated with IL-1β release (Nisticò et al. 2013), indicating similar mechanisms of synaptic plasticity alterations in both disorders. We observed that the more abundant sources of IL-1β in EAE hippocampal slices were sparse lesion sites characterized by the presence of activated microglia and localized in proximity of the dentate gyrus, CA3 region and fimbria (Nisticò et al. 2013). In MS and EAE, lesion sites in the brain and spinal cord are typically enriched of CD3+ T lymphocyte infiltrates which are, along with activated microglia and astroglia, a major cause of inflammatory cytokine release (Centonze et al. 2009; Mandolesi et al. 2012; Nisticò et al. 2013). Therefore, we investigated by immunofluorescence and confocal imaging whether in lesion sites of EAE hippocampus (20 dpi) CD3+ lymphocytes were present. As shown in Fig. 2a, CD3+ cells were clearly present in EAE hippocampal lesions (Fig. 2a′, a″) relative to CFA hippocampus, which did not present any infiltrating lymphocyte (Fig. 2b′, b″).

Presence of CD3+ lymphocytes in EAE hippocampus and astrogliosis. a, b Immunostaining of hippocampal coronal sections, counterstained with the nuclear marker dapi (white), showing CD3-positive (red) lymphocytes (a″) abundantly present in the lesion sites of EAE mice (a, a′, dapi staining) (20 dpi; score ≥ 2). No infiltrates and lesion sites were detectable in CFA hippocampus (b, b′, b″). a′, a″ and b′, b″ are high magnifications of the white boxes in a and b, respectively. c, d GFAP immunostaining of coronal hippocampal slices counterstained with dapi (white) showing a consistent astroglia activation in EAE hippocampus (c) relative to the CFA group (d). c′, c″ and d′, d″ show high magnifications of the CA1 regions in c and d, respectively. Scale bars in a, b, c, d are 200 μm; in a′, a″, b′, b″, c′, c″, d′, d″ are 20 μm (Color figure online)
To provide a complete characterization of the inflammatory reaction occurring in the EAE hippocampus during the acute phase of the disease, we performed astrocyte immunostaining. As shown in Fig. 2c and d, a widespread increase in the intermediate filament protein GFAP with accompanying cellular hypertrophy was evident in EAE hippocampal slices (Fig. 2c′, c″) compared to CFA (Fig. 2d′, d″). Upregulation of GFAP by astrocytes is regarded as a hallmark of astrogliosis (Zhang et al. 2010).
T Lymphocytes from EAE Mice Alter Bidirectional Synaptic Plasticity in the Hippocampus
Thus, to assess the potential involvement of CD3+ T cells in the synaptic alterations found in EAE mice and possibly in MS patients, we pre-incubated (30 min to 1 h) hippocampal slices from C57BL/6 mice in the presence of CD3+ cells extracted from the spleens of EAE (25 dpi) or CFA mice. We found that PP-LFS at 1 Hz for 15 min, a protocol known to induce LTD in adult rodents (Kemp et al. 2000), triggered LTD in C57BL/6 slices pre-incubated with CFA-specific T lymphocytes (n = 7), whereas synaptic responses returned to baseline levels following incubation with EAE-specific T lymphocytes (n = 9) (C57BL/6 + lymph CFA, 0.62 ± 0.05; C57BL/6 + lymph EAE, 1.03 ± 0.04; p < 0.05) (Fig. 3a). Along the same line, LTP magnitude was significantly enhanced following incubation with EAE-specific T lymphocytes (n = 5) compared to CFA-specific T lymphocytes (n = 5) (C57BL/6 + lymph CFA, 1.24 ± 0.07; C57BL/6 + lymph EAE, 1.75 ± 0.08; p < 0.05) (Fig. 3b). Overall, our results demonstrate that LTP appearance is favored over LTD in the presence of EAE-specific T lymphocytes.

Bidirectional synaptic plasticity is altered in the hippocampus by EAE lymphocytes. a Sample traces (above) and superimposed graph (below) showing the normalized changes in field potential slope (mean ± SEM) induced by PP-LFS in slices from C57BL/6Jmice incubated with CFA (n = 7) or EAE (n = 9) lymphocytes. Insets show fEPSPs from representative experiments during a baseline interval and 60 min after delivery of conditioning train. b Sample traces (above) and superimposed graph (below) showing the normalized changes in field potential slope (mean ± SEM) induced by HFS in slices from C57BL/6Jmice incubated with CFA (n = 5) or EAE (n = 5) lymphocytes
Remarkably, co-incubation with IL-1ra was able to antagonize the effect mediated by EAE T cell lymphocytes (n = 6) (C57BL/6 + lymph EAE + IL-1ra, 0.82 ± 0.06; p < 0.05) (Fig. 4).

Sample traces (above) and summary graph (below) of the averaged time course of LTD induced by PP-LFS in C57BL/6J slices pre-incubated with EAE lymphocytes with or without IL-1ra application (10 μg/ml, n = 6)
T Lymphocytes from EAE Mice Alter Hippocampal GABA Transmission via IL-1β
Defective GABA transmission is responsible for reduced LTP threshold in EAE (Nisticò et al. 2013). Therefore we studied the effects of CD3+ T cells on GABAergic sIPSCs from CA1 pyramidal neurons in the continuous presence of CNQX (10 μM). In slices pre-incubated with EAE-specific T lymphocytes, we found a reduced efficacy of GABAergic transmission compared to slices incubated with T lymphocytes taken from CFA mice. Accordingly, we found a reduced sIPSC amplitude (p < 0.001, K–S test on the cumulative distributions; p < 0.05, t test on the median values; Fig. 5a, left) and larger inter-event interval (p < 0.001, K–S test on the cumulative distributions; p < 0.001, t test on the median values; Fig. 5a, right) caused by EAE-specific T lymphocytes, which release higher amount of IL1-β compared to CFA-CD3+ cells (Mandolesi et al. 2013). Moreover, such effect of lymphocytes on GABAergic transmission was reminiscent of the alterations seen in EAE slices or in response to bath-applied IL-1β (Nisticò et al. 2013).

IL-1β-dependent inhibition of the GABAergic transmission by EAE lymphocytes. Pooled (mean ± SEM) cumulative distributions of sIPSCs amplitude (left; bin size 10 pA) and inter-event interval (right; bin size 50 ms) recorded from neurons of C57BL/6 mice pre-incubated in CFA lymphocytes (n = 9) of EAE lymphocytes (n = 10) mice. Histograms in the insets are averages (mean ± SEM) of the corresponding median values. On top are trace records from typical neurons exposed to CFA (left) or EAE (right) lymphocytes. b Pooled (mean ± SEM) cumulative distributions of sIPSCs amplitude (left; bin size 10 pA) and inter-event interval (right; bin size 50 ms) recorded from neurons of C57BL/6 mice pre-incubated in EAE lymphocytes alone (n = 10) or EAE lymphocytes with the IL-1β receptor antagonist IL-1ra (10 μg/ml; n = 11). Histograms in the insets are averages (mean ± SEM) of the corresponding median values. On top are trace records from typical neurons exposed to EAE lymphocytes alone (left) or EAE lymphocytes with IL-1ra (right). * and ***Indicate p < 0.05 and 0.001 t test, respectively
The decrease in the efficacy of GABAergic transmission by EAE lymphocytes was prevented by co-incubation with the IL-1β receptor antagonist IL-1ra (10 μg/ml). In the presence of IL-1ra, we recorded sIPSCs of higher amplitude (p < 0.01, K–S test on the cumulative distributions; p < 0.05, t test on the median values; Fig. 5b, left) and lower inter-event interval (p < 0.01, K–S test on the cumulative distributions; p < 0.05, t test on the median values; Fig. 5b, right).
In vivo Inhibition of IL-1β Rescues Inflammatory and Synaptic Damage in EAE Hippocampus
Besides an acute inhibitory effect of IL-1β on GABAergic signaling, we wanted to investigate the long-term effect of IL-1β on hippocampal synaptopathy correlated with GABAergic degeneration in EAE mice (Ziehn et al. 2010, 2012; Nisticò et al. 2013).
To this aim, we blocked IL-1β signaling in vivo by chronic icv infusion of IL-1ra, the physiological antagonist of the proinflammatory cytokine IL-1β. The treatment consisted in a continuous delivery for four weeks of IL-1ra (EAE-IL-1ra) or vehicle (EAE vehicle) through osmotic minipumps starting 1 week before immunization. In correspondence of the acute phase of the disease (20–25 dpi) and at the end of the treatment, we killed the mice of both experimental groups and performed immunofluorescence and WB experiments. Of note, during this phase of the disease, EAE-IL-1ra mice presented a mild EAE severity in comparison to their relative controls (data not shown) in accordance to our previous observations (Mandolesi et al. 2013).
We studied the inflammatory reaction in these mice by quantification of Iba-1 protein, a marker of microglial activation which is abundantly expressed in EAE hippocampus during the acute phase (Nisticò et al. 2013). As shown in Fig. 6a–c, the Iba-1 immunostaining in EAE-IL-1ra hippocampal slices (Fig. 6b) was less prominent relatively to the EAE-vehicle group (Fig. 6a) and the amoeboid-activated morphology typical of microglia cells in EAE hippocampus (Fig. 6a′) was less pronounced in EAE-IL-1ra group (Fig. 6b′). Such observation was supported by WB quantification of the Iba-1 protein (Fig. 6c), which was significantly decreased in EAE-IL-1ra (Iba1/GAPDH ratio: 0.87 ± 0.145, n = 4) protein extracts relatively to the EAE vehicle (1.46 ± 0.058, n = 5), but not significantly different from the CFA group (CFA vehicle 1 ± 0.079, n = 4; one-way ANOVA p = 0.0021: CFA vehicle vs. EAE vehicle p < 0.01, CFA vehicle vs. EAE-IL-1ra p > 0.05, EAE vehicle vs. EAE-IL-1ra p < 0.01).

In vivo inhibition of IL-1β signaling ameliorates synaptic alterations and inflammatory reaction in EAE hippocampus. a–c Immunostaining and WB quantification of Iba-1 protein in EAE-IL-1ra and EAE-vehicle treated mice. Double staining of Iba1-positive microglia/macrophage cells (green) and of the cell nuclei (white) shows less inflammatory reaction in EAE-IL-1ra slices (b, b′) in comparison to EAE mice (a, a′). More resting “ramified” cells are present in EAE-IL-1ra hippocampi (b′), while in EAE-vehicle Iba1 positive cells show a typical amoeboid-activated cell morphology (a′). c Quantitative WB analysis shows a significant reduction of Iba1 in EAE-IL-1ra hippocampal protein extracts relative to the EAE-vehicle group, reaching a mean value similar to that of CFA-vehicle mice. d WB quantification of synapsin levels in the hippocampi of EAE-IL-1ra, EAE-vehicle and CFA-vehicle mice (22 dpi). The IL-1ra treatment induced a significant preservation of the synaptic inputs. WB data were normalized to β-tubulin. Data were presented as the mean ± SEM; one-way ANOVA: EAE vehicle vs. EAE-IL1ra *p < 0.05, **p < 0.01, CFA vehicle vs. EAE vehicle # p < 0.05. Scale bars in a and b: 200 μm; in a′ and b′: 10 μm (Color figure online)
Due to the strong impact of the icv IL-1ra treatment in triggering the inflammatory response, we analyzed the expression level of the pre-synaptic marker synapsin to evaluate the integrity of hippocampal circuitry in EAE mice (Ziehn et al. 2012). Quantitative WB analysis (Fig. 6d) showed that EAE-vehicle hippocampal protein extracts presented a significant reduction in the synaptic marker during the acute phase of the disease (synapsin/β tubulin ratio: 0.641 ± 0.068, n = 5) while in IL-1ra-treated mice, the synapsin expression reached normal values (1.04 ± 0.071, n = 4) in comparison to CFA mice (1 ± 0.131, n = 4; one-way ANOVA p = 0.0175: CFA vehicle vs. EAE vehicle p < 0.05, CFA vehicle vs. EAE-IL-1ra p > 0.05, EAE vehicle vs. EAE-IL-1ra p < 0.05).
Such results provide an in vivo demonstration of the crucial role played by IL-1β signaling in hippocampal synaptic degeneration in EAE mice.
Discussion
In the present study, we showed a reversed polarity of cortical synaptic plasticity in response to cTBS in MS patients, and found that CSF levels of the pro-inflammatory cytokine IL-1β correlated with the amplitude of this aberrant LTP-like phenomenon. Together with our recent finding that also iTBS-induced plasticity is altered in MS (Mori et al. 2011), these results indicate a profound subversion of plasticity rules and mechanisms in the MS brain, which is likely caused by the inflammatory milieu generated by infiltrating autoreactive T lymphocytes in the CNS during immune-mediated attacks. In this respect, a previous study reported the expected LTD-like plasticity in response to cTBS in a sample of 14 stable MS patients (Zeller et al. 2012), while about half of our studied population was in relapsing phase.
The mechanism by which cTBS enhanced cortical excitability in our sample of MS patients is unknown. It has been postulated that TBS induces a mixture of excitatory and inhibitory long-lasting effects, the direction of which being determined in physiological conditions by the excitation/inhibition balance (Huang et al. 2011). Thus, either a reduced GABA inhibition (Caramia et al. 2004; Rossi et al. 2012b) or an increased glutamate release (Rossi et al. 2012a), which have been both reported in the MS brain, may likely play significant roles in the LTD deficit seen in MS patients in response to cTBS. Indeed, a magnetic resonance spectroscopy study in healthy subjects showed that cTBS over M1 increases GABA concentration without affecting glutamate levels (Stagg et al. 2009) suggesting that GABA may play an important role in the generation of cTBS after-effects in humans. Implication of GABA in the generation of LTD phenomena in the neocortex has been observed also in animals (Hess and Donoghue 1996; Komaki et al. 2007; Trepel and Racine 2000). For example, attenuation of LTP by GABAA receptor agonists has been reported (Chapman et al. 1998; Grover and Yan 1999; Levkovitz et al. 1999; Lu et al. 2000; Martin et al. 2010; Wigström and Gustafsson 1983; Yoshiike et al. 2008), while low-to-moderate concentrations of GABAB receptor antagonists are able to facilitate LTP (Hess and Donoghue 1996; Stäubli et al. 1999).
Both LTP and LTD of excitatory synaptic transmission can be induced experimentally at a same synapse in response to repetitive synaptic activation, being LTP threshold generally higher than LTD threshold, and therefore requiring more intense stimulation for its induction (Gambrill et al. 2011; Kirkwood et al. 1996). Inhibition of GABA synapses has already been found to play a major role in lowering LTP threshold in EAE mice (Nisticò et al. 2013), and here we recognized for the first time the ability of EAE-specific activated T lymphocytes in promoting LTP appearance over LTD likely by suppressing GABAergic tone. We indeed observed in the hippocampus of EAE mice the presence of infiltrating T lymphocytes, which are mainly localized in the lesion sites. Interestingly, it has been shown that during EAE not only Th1 lymphocytes, but also Tregs and naïve T cells cross the blood brain barrier (BBB) (Korn et al. 2007; Cose 2007; Herz et al. 2011). In particular, naïve T cells migrating in the brain may contribute to re-establish the correct immune tolerance, by differentiating into T regulatory cells. However, in this study, we did not investigate the exact nature of the infiltrates and their potentially anti-inflammatory effects.
Conversely, it is known that CD3+ infiltration combined with microglia activation promotes a prominent inflammatory reaction during the acute phase of the disease, including astrogliosis. We indeed observed also a strong astroglia activation, a hallmark of EAE brain (Grasselli et al. 2013; Mandolesi et al. 2013) and of MS (Lycke et al. 1998). Important triggers and modulators of astrogliosis are proinflammatory cytokines released by activated microglia (Zhang et al. 2010). The function of reactive astrocytes is not well understood and both harmful and beneficial activities were reported (Zhang et al. 2010). Moreover, astrocytes produce cytokines that can act on microglia, thus creating a paracrine and autocrine feedback loop whereby microglia-derived factors and astroglial-derived factors regulate each other (Lotan and Schwartz 1994) and the neuronal compartment.
Lymphocytes and activated microglia are sources of IL-1β (Nisticò et al. 2013). Here, we focused our attention on CD3+ cells and found that T lymphocytes from EAE-spleens, but not from control mice, release IL-1β and alter bidirectional synaptic plasticity in normal hippocampus, mimicking the facilitated LTP induction observed in the cortex of MS patients with high CSF levels of IL-1β. The specific molecular mechanisms that mediate IL-1β effect is still largely unknown, however, its acute effect on synaptic transmission has been observed in different brain regions and seems to involve both pre-synaptic and post-synaptic mechanisms (Musumeci et al. 2011; Mandolesi et al. 2012, 2013; Rossi et al. 2012a, b). In addition, literature provides evidence for both acute and long-lasting effects, requiring post-translational modifications and altered gene expression of channel proteins (Schäfers and Sorkin 2008). At this regard, IL-1β influences the expression of ionotropic receptors involved in synaptic function and LTP induction and maintenance, such as NMDA (Coogan and O’Connor 1997; Viviani et al. 2006) and AMPA receptors (Lai et al. 2006).
The effects induced by IL-1β on the cTBS protocol resemble the metaplastic changes described by Gentner et al. (2008). These authors in fact found that 300 stimuli of cTBS without prior voluntary motor activation facilitate MEP amplitude, but turn to the expected inhibition when preceded by sustained voluntary muscle activity. Also phasic voluntary movements reverse subsequent cTBS 600 after-effects into an LTP-like effect (Iezzi et al. 2008). Homeostatic synaptic plasticity relies on the Bienenstock–Cooper–Munro theory (Bienenstock et al. 1982) proposing that integrated post-synaptic activity modulates the threshold for LTP/LTD induction after a stimulation protocol. Therefore, low post-synaptic activity increases the probability for LTP induction, while high levels increase the probability for LTD induction. Similar homeostatic mechanisms may regulate synaptic plasticity also in the human motor cortex. Indeed, the normally induced inhibitory after-effects of 1-Hz rTMS can be reversed to facilitation when preceded by another inhibitory protocol such as cathodal transcranial DC stimulation (Siebner et al. 2004).
A growing body of experimental and clinical studies in MS proposes that the immune-mediated enhancement of glutamatergic synaptic transmission and the unbalance between GABAergic and glutamatergic transmission may cause degenerative neuronal damage through excitotoxicity (Rossi et al. 2012a). On the other hand, it is believed that after a brain lesion LTP mechanisms increase synaptic excitability of denervated neurons to functionally compensate neuronal loss and mediate improvements of neurological defects. Thus, although neuronal and synaptic degeneration occur, their clinical consequences are initially well compensated in MS and EAE, likely due to plastic adaptations of surviving neurons (Schirmer et al. 2013). The precise nature of the adaptation mechanisms taking place in MS brains is still unknown, however, LTP occurrence in spared neurons might well compensate for neuronal loss in acute and chronic neurological diseases. Functional MRI studies in MS showed that in the presence of severe CNS injury, more extended neuronal activation in the cortex was associated with less disability (Reddy et al. 2000; Rocca et al. 2002), suggesting that the capacity to recruit more extended neuronal activation may attenuate the effect of CNS injury. On the other hand, after a prolonged time, an abnormal enhancement of neuronal excitability could become in turn detrimental.
Finally, we evaluated a potential long-term effect of IL-1β on synaptic and neuronal degeneration occurring in EAE hippocampus. A hallmark of the EAE hippocampus during the acute phase of the disease is indeed a neuronal degeneration, in particular of GABAergic neurons, accompanied by a robust decrease in synaptic inputs (Nisticò et al. 2013; Ziehn et al. 2012; Ziehn et al. 2010). We observed that the preventive IL-1ra treatment was able to ameliorate the inflammatory reaction in the EAE hippocampus and also the clinical disability of the mice (Mandolesi et al. 2013). These results highlight the importance of IL-1β as a suitable therapeutic target in MS, as already suggested by a previous work based on a gene therapy strategy (Furlan et al. 2007). In addition, we demonstrated effectiveness of the icv treatment on hippocampal synaptopathy in correlation with a strongly reduced inflammatory reaction. Such observations strengthen the idea that IL-1β, by acting directly on the neuronal system, can induce also degenerative mechanisms potentially causing detrimental effects on synaptic homeostasis.
In conclusion, the present work suggests a role of IL-1β as a mediator of synaptic plasticity alterations in MS and its experimental model EAE. Facilitated LTP during immune attacks of the CNS might play a role in the recovery of function after a lesion, but also in cognitive deficits and excitotoxic neurodegeneration in this disorder.
References
Aktas, O., & Zipp, F. (2003). Regulation of self-reactive T cells by human immunoglobulins-implications for multiple sclerosis therapy. Current Pharmaceutical Design, 9(3), 245–256.
Balducci, C., Mehdawy, B., Mare, L., Giuliani, A., Lorenzini, L., Sivilia, S., et al. (2011). The γ-secretase modulator CHF5074 restores memory and hippocampal synaptic plasticity in plaque-free Tg2576 mice. Journal of Alzheimers Disease, 24(4), 799–816.
Barbizan, R., & Oliveira, A. L. (2010). Impact of acute inflammation on spinal motoneuron synaptic plasticity following ventral root avulsion. Journal of Neuroinflammation, 7, 29.
Bienenstock, E. L., Cooper, L. N., & Munro, P. W. (1982). Theory for the development of neuron selectivity: Orientation specificity and binocular interaction in visual cortex. The Journal of Neuroscience, 2(1), 32–48.
Caramia, M. D., Palmieri, M. G., Desiato, M. T., Boffa, L., Galizia, P., Rossini, P. M., et al. (2004). Brain excitability changes in the relapsing and remitting phases of multiple sclerosis: A study with transcranial magnetic stimulation. Clinical Neurophysiology, 115(4), 956–965.
Centonze, D., Muzio, L., Rossi, S., Cavasinni, F., De Chiara, V., Bergami, A., et al. (2009). Inflammation triggers synaptic alteration and degeneration in experimental autoimmune encephalomyelitis. The Journal of Neuroscience, 29(11), 3442–3452.
Chapman, C. A., Perez, Y., & Lacaille, J. C. (1998). Effects of GABA(A) inhibition on the expression of long-term potentiation in CA1 pyramidal cells are dependent on tetanization parameters. Hippocampus, 8(3), 289–298.
Coogan, A., & O’Connor, J. J. (1997). Inhibition of NMDA receptor-mediated synaptic transmission in the rat dentate gyrus in vitro by IL-1β. Neuroreport, 8(9–10), 2107–2110.
Cose, S. (2007). T-cell migration: A naive paradigm? Immunology, 120(1), 1–7.
Di Filippo, M., Chiasserini, D., Gardoni, F., Viviani, B., Tozzi, A., Giampà, C., et al. (2013). Effects of central and peripheral inflammation on hippocampal synaptic plasticity. Neurobiology of Disease,. doi:10.1016/j.nbd.2012.12.009.
Di Lazzaro, V., Pilato, F., Saturno, E., Oliviero, A., Dileone, M., Mazzone, P., et al. (2005). Theta-burst repetitive transcranial magnetic stimulation suppresses specific excitatory circuits in the human motor cortex. The Journal of Physiology, 565(Pt3), 945–950.
Errico, F., Nisticò, R., Napolitano, F., Oliva, A. B., Romano, R., Barbieri, F., et al. (2011). Persistent increase of D-aspartate in D-aspartate oxidase mutant mice induces a precocious hippocampal age-dependent synaptic plasticity and spatial memory decay. Neurobiology of Aging, 32(11), 2061–2074.
Furlan, R., Bergami, A., Brambilla, E., Butti, E., De Simoni, M. G., Campagnoli, M., et al. (2007). HSV-1-mediated IL-1 receptor antagonist gene therapy ameliorates MOG(35–55)-induced experimental autoimmune encephalomyelitis in C57BL/6 mice. Gene Therapy, 14(1), 93–98.
Gambrill, A. C., Storey, G. P., & Barria, A. (2011). Dynamic regulation of NMDA receptor transmission. Journal of Neurophysiology, 105(1), 162–171.
Gentner, R., Wankerl, K., Reinsberger, C., Zeller, D., & Classen, J. (2008). Depression of human corticospinal excitability induced by magnetic theta-burst stimulation: Evidence of rapid polarity-reversing metaplasticity. Cerebral Cortex, 18(9), 2046–2053.
Grasselli, G., Rossi, S., Musella, A., Gentile, A., Loizzo, S., Muzio, L., et al. (2013). Abnormal NMDA receptor function exacerbates experimental autoimmune encephalomyelitis. Britsh Journal of Pharmacology, 168(2), 502–517.
Grover, L. M., & Yan, C. (1999). Blockade of GABAA receptors facilitates induction of NMDA receptor-independent long-term potentiation. Journal of Neurophysiology, 81(6), 2814–2822.
Haji, N., Mandolesi, G., Gentile, A., Sacchetti, L., Fresegna, D., Rossi, S., et al. (2012). TNF-α-mediated anxiety in a mouse model of multiple sclerosis. Experimental Neurology, 237(2), 296–303.
Herz, J., Paterka, M., Niesner, R. A., Brandt, A. U., Siffrin, V., Leuenberger, T., et al. (2011). In vivo imaging of lymphocytes in the CNS reveals different behaviour of naïve T cells in health and autoimmunity. Journal of Neuroinflammation, 8, 131.
Hess, G., & Donoghue, J. P. (1996). Long-term depression of horizontal connections in rat motor cortex. The European Journal of Neuroscience, 8(4), 658–665.
Huang, Y. Z., Edwards, M. J., Rounis, E., Bhatia, K. P., & Rothwell, J. C. (2005). Theta burst stimulation of the human motor cortex. Neuron, 45(2), 201–206.
Huang, Y. Z., Rothwell, J. C., Chen, R. S., Lu, C. S., & Chuang, W. L. (2011). The theoretical model of theta burst form of repetitive transcranial magnetic stimulation. Clinical Neurophysiology, 122(5), 1011–1018.
Iezzi, E., Conte, A., Suppa, A., Agostino, R., Dinapoli, L., Scontrini, A., et al. (2008). Phasic voluntary movements reverse the aftereffects of subsequent theta-burst stimulation in humans. Journal of Neurophysiology, 100(4), 2070–2076.
Jenkins, T. M., Toosy, A. T., Ciccarelli, O., Miszkiel, K. A., Wheeler-Kingshott, C. A., Henderson, A. P., et al. (2010). Neuroplasticity predicts outcome of optic neuritis independent of tissue damage. Annals of Neurology, 67(1), 99–113.
Kemp, N., McQueen, J., Faulkes, S., & Bashir, Z. I. (2000). Different forms of LTD in the CA1 region of the hippocampus: Role of age and stimulus protocol. The European Journal of Neuroscience, 12(1), 360–366.
Kim do, Y., Hao, J., Liu, R., Turner, G., Shi, F. D., & Rho, J. M. (2012). Inflammation-mediated memory dysfunction and effects of a ketogenic diet in a murine model of multiple sclerosis. PLoS One, 7(5), e35476.
Kirkwood, A., Rioult, M. C., & Bear, M. F. (1996). Experience-dependent modification of synaptic plasticity in visual cortex. Nature, 381(6582), 526–528.
Komaki, A., Shahidi, S., Lashgari, R., Haghparast, A., Malakouti, S. M., & Noorbakhsh, S. M. (2007). Effects of GABAergic inhibition on neocortical long-term potentiation in the chronically prepared rat. Neuroscience Letters, 422, 181–186.
Korn, T., Reddy, J., Gao, W., Betelli, E., Awasthi, A., Petersen, T. R., et al. (2007). Myelin-specific regulatory T cells accumulate in the CNS but fail to control autoimmune inflammation. Nature Medicine, 13, 423–431.
Lai, A. Y., Swayze, R. D., El-Husseini, A., & Song, C. (2006). Interleukin-1 beta modulates AMPA receptor expression and phosphorylation in hippocampal neurons. Journal of Neuroimmunology, 175(1–2), 97–106.
Levkovitz, Y., Avignone, E., Groner, Y., & Segal, M. (1999). Upregulation of GABA neurotransmission suppresses hippocampal excitability and prevents long-term potentiation in transgenic superoxide dismutase-overexpressing mice. The Journal of Neuroscience, 19(24), 10977–10984.
Lotan, M., & Schwartz, M. (1994). Cross talk between the immune system and the nervous system in response to injury: Implications for regeneration. The FASEB Journal, 8, 1026–1033.
Lu, Y. M., Mansuym, I. M., Kandel, E. R., & Roder, J. (2000). Calcineurin-mediated LTD of GABAergic inhibition underlies the increased excitability of CA1 neurons associated with LTP. Neuron, 26(1), 197–205.
Lycke, J. N., Karlsson, J. E., Andersen, O., & Rosengren, L. E. (1998). Neurofilament protein in cerebrospinal fluid: A potential marker of activity in multiple sclerosis. Journal of Neurology, Neurosurgery and Psychiatry, 64, 402–404.
Malenka, R. C., & Bear, M. F. (2004). LTP and LTD: An embarrassment of riches. Neuron, 44(1), 5–21.
Mandolesi, G., Grasselli, G., Musella, A., Gentile, A., Musumeci, G., Sepman, H., et al. (2012). GABAergic signaling and connectivity on Purkinje cells are impaired in experimental autoimmune encephalomyelitis. Neurobiology of Disease, 46(2), 414–424.
Mandolesi, G., Grasselli, G., Musumeci, G., & Centonze, D. (2010). Cognitive deficits in experimental autoimmune encephalomyelitis: Neuroinflammation and synaptic degeneration. Neurological Sciences, 31(Suppl 2), S255–S259.
Mandolesi, G., Musella, A., Gentile, A., Grasselli, G., Haji, N., Sepman, H., et al. (2013). Interleukin-1β alters glutamate transmission at Purkinje cell synapses in a mouse model for multiple sclerosis. The Journal of Neuroscience, 33(29), 12105–12121.
Martin, L. J., Zurek, A. A., MacDonald, J. F., Roder, J. C., Jackson, M. F., & Orser, B. A. (2010). Alpha5GABAA receptor activity sets the threshold for long-term potentiation and constrains hippocampus-dependent memory. The Journal of Neuroscience, 30(15), 5269–5282.
Mori, F., Rossi, S., Sancesario, G., Codecà, C., Mataluni, G., Monteleone, F., et al. (2011). Cognitive and cortical plasticity deficits correlate with altered amyloid-β CSF levels in multiple sclerosis. Neuropsychopharmacology, 36(3), 559–568.
Musumeci, G., Grasselli, G., Rossi, S., De Chiara, V., Musella, A., Motta, C., et al. (2011). Transient receptor potential vanilloid 1 channels modulate the synaptic effects of TNF-α and of IL-1β in experimental autoimmune encephalomyelitis. Neurobiology of Disease, 43(3), 669–677.
Nisticò, R., Mango, D., Mandolesi, G., Piccinin, S., Berretta, N., Pignatelli, M., et al. (2013). Inflammation subverts hippocampal synaptic plasticity in experimental multiple sclerosis. PLoS One, 8(1), e54666.
Nisticò, R., Pignatelli, M., Piccinin, S., Mercuri, N. B., & Collingridge, G. (2012). Targeting synaptic dysfunction in Alzheimer’s disease therapy. Molecular Neurobiology, 46(3), 572–587.
Polman, C. H., Reingold, S. C., Banwell, B., Clanet, M., Cohen, J. A., Filippi, M., et al. (2011). Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Annals of Neurology, 69(2), 292–302.
Reddy, H., Narayanan, S., Arnoutelis, R., Jenkinson, M., Antel, J., Matthews, P. M., et al. (2000). Evidence for adaptive functional changes in the cerebral cortex with axonal injury from multiple sclerosis. Brain, 123(11), 2314–2320.
Rocca, M. A., Falini, A., Colombo, B., Scotti, G., Comi, G., & Filippi, M. (2002). Adaptive functional changes in the cerebral cortex of patients with non disabling multiple sclerosis correlate with the extent of brain structural damage. Annals Neurology, 51(3), 330–339.
Rossi, S., De Chiara, V., Furlan, R., Musella, A., Cavasinni, F., Muzio, L., et al. (2010). Abnormal activity of the Na/Ca exchanger enhances glutamate transmission in experimental autoimmune encephalomyelitis. Brain, Behavior, and Immunity, 24(8), 1379–1385.
Rossi, S., Furlan, R., De Chiara, V., Motta, C., Studer, V., Mori, F., et al. (2012a). Interleukin-1β causes synaptic hyperexcitability in multiple sclerosis. Annals of Neurology, 71(1), 76–83.
Rossi, S., Studer, V., Motta, C., De Chiara, V., Barbieri, F., Bernardi, G., et al. (2012b). Inflammation inhibits GABA transmission in multiple sclerosis. Multiple Sclerosis, 18(11), 1633–1635.
Rothwell, J. C., Hallett, M., Berardelli, A., Eisen, A., Rossini, P., & Paulus, W. (1999). Magnetic stimulation: motor evoked potentials. The International Federation of Clinical Neurophysiology. Electroencephalography and Clinical Neurophysiology. Supplement, 52, 97–103.
Schäfers, M., & Sorkin, L. (2008). Effect of cytokines on neuronal excitability. Neuroscience Letters, 437(3):188–193. Erratum in: Neuroscience Letters. 2010 481(3):191.
Schirmer, L., Merkler, D., König, F. B., Brück, W., & Stadelmann, C. (2013). Neuroaxonal regeneration is more pronounced in early multiple sclerosis than in traumatic brain injury lesions. Brain Pathology, 23(1), 2–12.
Siebner, H. R., Lang, N., Rizzo, V., Nitsche, M. A., Paulus, W., Lemon, R. N., et al. (2004). Preconditioning of low-frequency repetitive transcranial magnetic stimulation with transcranial direct current stimulation: Evidence for homeostatic plasticity in the human motor cortex. The Journal of Neuroscience, 24(13), 3379–3385.
Stagg, C. J., Wylezinska, M., Matthews, P. M., Johansen-Berg, H., Jezzard, P., Rothwell, J. C., et al. (2009). Neurochemical effects of theta burst stimulation as assessed by magnetic resonance spectroscopy. Journal of Neurophysiology, 101(6), 2872–2877.
Stäubli, U., Scafidi, J., & Chun, D. (1999). GABAB receptor antagonism: Facilitatory effects on memory parallel those on LTP induced by TBS but not HFS. The Journal of Neuroscience, 19(11), 4609–4615.
Trepel, C., & Racine, R. J. (2000). GABAergic modulation of neocortical long-term potentiation in the freely moving rat. Synapse (New York, N.Y.), 35(2), 120–128.
Viviani, B., Gardoni, F., Bartesaghi, S., Corsini, E., Facchi, A., Galli, C. L., et al. (2006). Interleukin-1 beta released by gp120 drives neural death through tyrosine phosphorylation and trafficking of NMDA receptors. The Journal of Biological Chemistry, 281(40), 30212–30222.
Wigström, H., & Gustafsson, B. (1983). Heterosynaptic modulation of homosynaptic long-lasting potentiation in the hippocampal slice. Acta Physiologica Scandinavica, 119(4), 455–458.
Yoshiike, Y., Kimura, T., Yamashita, S., Furudate, H., Mizoroki, T., Murayama, M., et al. (2008). GABA(A) receptor-mediated acceleration of aging-associated memory decline in APP/PS1 mice and its pharmacological treatment by picrotoxin. PLoS One, 3(8), e3029.
Zeller, D., Dang, S. Y., Weise, D., Rieckmann, P., Toyka, K. V., & Classen, J. (2012). Excitability decreasing central motor plasticity is retained in multiple sclerosis patients. BMC Neurology, 12, 92.
Zhang, D., Hu, X., Qian, L., O’Callaghan, J. P., & Hong, J. S. (2010). Astrogliosis in CNS pathologies: Is there a role for microglia? Molecular Neurobiology, 41(2–3), 232–241.
Ziehn, M. O., Avedisian, A. A., Dervin, S. M., Umeda, E. A., O’Dell, T. J., & Voskuhl, R. R. (2012). Therapeutic testosterone administration preserves excitatory synaptic transmission in the hippocampus during autoimmune demyelinating disease. The Journal of Neuroscience, 32(36), 12312–12324.
Ziehn, M. O., Avedisian, A. A., Tiwari-Woodruff, S., & Voskuhl, R. R. (2010). Hippocampal CA1 atrophy and synaptic loss during experimental autoimmune encephalomyelitis, EAE. Laboratory Investigation, 90(5), 774–786.
Acknowledgments
This investigation was supported by Fondazione Italiana Sclerosi Multipla (Progetto Speciale FISM), by the Italian National Ministry of Health and by the Italian National Ministry of University to DC. We are indebted to Dr. Mauro Federici and Vladimiro Batocchi for excellent technical assistance.
Conflict of interest
Diego Centonze acted as an Advisory Board member of Merck Serono, Teva, Bayer Schering, Biogen Idec, Novartis, and received funding for traveling and honoraria for speaking or consultation fees from Merck Serono, Teva, Novartis, Bayer Schering, Sanofi-aventis, Biogen Idec. He is also the principal investigator in clinical trials for Novartis, Merck Serono, Teva, Bayer Schering, Sanofi-aventis, Biogen Idec, Roche. The other authors have no disclosures to report.
Author information
Authors and Affiliations
Corresponding author
Additional information
Francesco Mori and Robert Nisticò are equally contributing first authors.
Rights and permissions
About this article
Cite this article
Mori, F., Nisticò, R., Mandolesi, G. et al. Interleukin-1β Promotes Long-Term Potentiation in Patients with Multiple Sclerosis. Neuromol Med 16, 38–51 (2014). https://doi.org/10.1007/s12017-013-8249-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12017-013-8249-7