
Abstract
The multiple inter-dependent post-translational modifications of histones represent fine regulators of chromatin dynamics. These covalent modifications, including phosphorylation, acetylation, ubiquitination, deimination, and methylation, affect therefore the numerous processes involving chromatin, such as replication, repair, transcription, genome stability, and cell death. Specific enzymes introducing modified residues in histones are precisely regulated, and a single amino acid residue can be subjected to a single or several, independent modifications. Disruption of histone post-translational modifications perturbs the pattern of gene expression, which may result in disease manifestations. It has become evident in recent years that apoptosis-modified histones exert a central role in the induction of autoimmunity, for example in systemic lupus erythematosus and rheumatoid arthritis. Certain histone post-translational modifications are linked to cell death (apoptotic and non-apoptotic cell death) and might be involved in lupus in the activation of normally tolerant lymphocyte subpopulations. In this review, we discuss how these modifications can affect the antigenicity and immunogenicity of histones with potential consequences in the pathogenesis of autoimmune diseases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Histones are a group of highly evolutionarily conserved proteins, which play a critical role in the proper packaging of DNA within the eukaryotic nucleus. Two copies of each of four core histones H2A, H2B, H3, and H4 associated to ~146 base pairs of DNA form the fundamental repeating subunit of chromatin, the so-called core particle. The structure known as the “nucleosome” contains ~180–200 base pairs of DNA and the histone octamer associated to a single copy of the extranucleosomal linker histone H1, which is partially replaced by H5 in nucleated cells of birds, fishes, and reptiles. It has been estimated that there are tens of millions of nucleosomes within a single human nucleus. Histones are not simple structural proteins; for decades, it is known that they actively participate in many different DNA-templated programs, including transcription, replication, recombination, and DNA repair. In these vital cellular pathways, a decisive role is played by post-translational modifications that affect histones and introduce in these proteins a very high degree of diversity. Histone modifications, which occur in highly controlled programs at different phases during the cell cycle, are involved alone or in combination in the fine regulation of distinct nuclear processes. Together with the enzymes that “write” the histone code and the regulatory proteins that “read” the code, histone modifications are thus critical components of eukaryotic development and differentiation pathways [1, 2].
Importantly, it has been shown that specific histone modifications, some of which are associated to apoptosis, might be central in pathological situations such as autoimmunity. Systemic autoimmune diseases, e.g., systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), or Sjögren's syndrome (SS), are characterized by antibodies that recognize a large array of self-components [3, 4]. Detailed studies have attributed a central role for apoptotic and non-apoptotic cell death in mediating tolerance break to these self-autoantigens [5–10]. These distinct forms of cell death occur in normal situations and generate new structures such as cleavage fragments or components containing post-translational modifications against which the immune system has normally been tolerized at the early stage of thymus development. In this context, however, it is anticipated that any defect in apoptosis or non-apoptotic cell death can either generate fragments with abnormal modifications (appearance of neo-epitopes recognized as foreign by the immune system) or delay elimination of modified/cleaved antigens, which might trigger the activation and proliferation of resting autoreactive lymphocytes [11, 12]. These normally anergic lymphocyte subpopulations might thus be involved in the early stages of the autoimmune spiral leading to an autoimmune disease. Post-translational modifications of autoantigens are described below with a focus on histone modifications, which have been claimed to be specifically related to autoimmune situations.
Phosphorylation and dephosphorylation of histones
Post-translational phosphorylation is one of the most common protein modifications found in animal cells. In these cells, serine, threonine, and tyrosine residues are subject to phosphorylation that occurs at the –COOH moiety of these residues and leads to the formation of O-phosphoserine, O-phosphothreonine, and O-phosphotyrosine. The ratio of phosphorylation of the three residues is approximately 1,000:100:1 for serine, threonine, and tyrosine residues, respectively. Phosphorylation/dephosphorylation events are generally transient. They are central in the majority of intracellular signaling pathways leading to stimulation or inhibition cascades, which regulate essential functions of the cell.
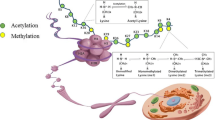
Numerous substrates of kinases and phosphatases represent major antigen targets in autoimmune diseases. Thus, the so-called Goodpasture antigen located in the C-terminus of the human α3 chain of collagen IV, the human ribosomal P0, P1, and P2 proteins, nucleolin/C23 antigen, the U1-70K small nuclear ribonucleoprotein, and SRp72 antigen are phosphoproteins. Histones, which are frequent targets for autoantibodies in SLE and other connective tissue diseases [13], can undergo phosphorylation, especially on their N-terminal tails, at Ser10 and Ser28 of H3, Ser1 of H4, and Ser14 of H2B, for example (Fig. 1). The presence of phosphoserine residues at certain sites in histones has been demonstrated to contribute to the nucleosome or the chromatin fiber remodeling and to positively or negatively influence the modification of other histone sites. H3 Ser10 phosphorylation was linked to mitotic chromosome condensation at the late G2 phase, while H3 Ser28 phosphorylation was associated with the early stage of the M phase (prophase). Interestingly, phosphorylation of histone H2B at Ser14 was shown to correlate with cells undergoing apoptosis [14].

Location of identified post-translational modifications affecting the four core histones and major histone autoepitopes described in the literature. Histone modifications associated with cell death are indicated in white letters; histone modifications identified as critical for autoantibody recognition are indicated on the respected autoepitopes below the black bar
Nothing is known on the possible involvement of histone phosphorylation state in the etiology of systemic autoimmune diseases or in the generation of the anti-histone autoimmune response. However, it is interesting to highlight the fact that the major B cell epitopes recognized by lupus antibodies in histones, and identified in using non-phosphorylated synthetic peptides, are mainly located in histone N-terminal tails, which contain most of the modification sites [15–17]. In addition, a major T cell epitope has been characterized in the N-terminal peptide 10–33 of H2B, which was reported to be tolerogenic in (SWRxNZB)F1 lupus mice [18, 19]. Along the same line, it is interesting to recall that the AHNAK protein, a 700-kD phosphoprotein identified as an antibody target in lupus [20], contains at its C-terminal domain three copies of the sequence Lys-Ser-Pro-Lys, a cdc2 kinase phosphorylation motif, also present in the peptide 170–185 of histone H1, which is frequently recognized by IgG antibodies from patients with SLE and SS [21].
Acetylation of histones and autoimmunity
It is known for a long time that in histones the acetylation/deacetylation of lysine residues plays an important role in the regulation of transcription. Acetylation of histones maintains an open and accessible conformation of chromatin, which is hereby available for the binding of the transcriptional machinery. The four core histones are particularly acetylated on their N-terminal tails, which influences the binding of the latter to DNA. In the N-terminal histone tails, epitopes targeted by lupus autoantibodies have been identified in histones H4 and H2B, which are prone to acetylation (17; Fig. 1). Recently, the involvement of the H4 N-terminal tail acetylation has been shown in lupus [22]. The epitope of KM-2, a monoclonal antibody generated from a MRL/lpr lupus mouse, was mapped by different approaches. KM-2 preferentially recognized H4 peptide 1–22 containing acetylated residues at positions 8 and/or 12 and/or 16. Any peptide containing an acetylated lysine residue at position 5 was not recognized by KM-2 antibody, and the non-acetylated peptide was less well recognized. Importantly, plasmas from both SLE patients and lupus mice showed a superior (but not exclusive) reactivity with acetylated N-terminal H4 peptides as well as with hyperacetylated histones isolated from cells treated with a histone deacetylase inhibitor. In addition, it was shown that the acetylation of histone H4 increased early during apoptosis. Monoclonal antibody KM-2 as well as plasmas from SLE patients and lupus mice showed an increased reactivity with apoptotic cells and histones isolated from apoptotic cells. The H4 peptide 1–22 acetylated on Lys8,12,16 residues was able to modulate autoimmune disease in MRL/lpr lupus-prone mice [22]. The tri-acetylated H4 peptide was administered subcutaneously in Freund’s adjuvant into pre-diseased MRL/lpr mice and was shown to enhance mortality, proteinuria, skin lesions, and glomerular IgG depositions. In contrast, the non-acetylated peptide had no effect. Hyperacetylated histones were able to maturate bone marrow-derived dendritic cells (DCs) in vitro, which produced increased levels of IL-6 and TNF-α compared with DCs cultured in the presence of nucleosomes purified from non-treated cells. In addition, DCs cultured in the presence of hyperacetylated nucleosomes were able to activate syngeneic T cells. Interestingly, Fab fragments that can bind acetylated Lys8 residue of histone H4 were selected from a phage-displayed repertoire of rearranged antibody genes from splenic B cells from a patient with SLE [23].
The enzymes involved in the acetylation and deacetylation of histones, i.e., the acetyltransferases and histone deacetylases, have been identified and play an important role in the regulation of transcription. Inhibitors of histone deacetylases such as trichostatin A and suberoylanilide hydroxamic acid increase the levels of histone acetylation in cells and subsequently alter gene expression. A hypoacetylation of histone H3 and H4 have been found in CD4+ T cells from SLE patients and splenocytes from MRL/lpr lupus mice [24, 25]. Treatment of MRL/lpr mice with histone deacetylase inhibitors resulted in resetting the epigenetic abnormalities to the normal situation [25–27]. In addition, treatment led to the improvement of glomerulonephritis and splenomegaly. The altered histone acetylation in T cells in SLE has been primarily linked to an altered gene expression. However, one can imagine that histones or chromatin released for instance from apoptotic T cells containing an altered modification pattern have an enhanced capability for activation of the immune system.
Histone ubiquitination and autoimmunity
Several proteins that are common targets in SLE and SS have been shown to be ubiquitinated. This is the case of the 52-Kd Ro/SSA protein and histones. Monoubiquitinated histones H2A (UH2A) and H2B (UH2B) modified at Lys119 of H2A and Lys120 in H2B (Fig. 1) represent 10% and 1% of total H2A and H2B proteins, respectively. Disappearance of UH2A has been linked to late apoptotic processes [28, 29]. Although detailed protocols of purification have been described, UH2A and UH2B remain difficult to purify in sufficient amounts for immunological investigations. To circumvent this technical problem, we synthesized chimeric peptides corresponding to the branched moiety of ubiquitinated histone UH2A and demonstrated that autoantibodies reacting with this branched peptide occur in ~60% of lupus patients, whereas they are present in less than 10% of patients with systemic sclerosis (SSc) and are virtually absent in patients with RA and primary SS [30]. This result was in good agreement with other data showing that autoantibodies to ubiquitin are detectable by ELISA in about 80% of lupus patients and in 0–13% only of patients with SS, RA, juvenile chronic arthritis, SSc, sarcoidosis, and normal individuals [31]. Human monoclonal antibodies reacting either with ubiquitin or with the UH2A branched peptide were also successfully generated by fusion of a human cell line with splenocytes from a young girl with active SLE who required splenectomy for severe thrombocytopenia [32]. Furthermore, using rabbit antibodies to UH2A peptide, it was possible to visualize UH2A deposits in renal biopsies from lupus patients [33]. The staining was granular in nature and was present in capillary and mesangial areas. However, deposition of ubiquitin, UH2A, or anti-ubiquitin antibodies was not observed in the kidneys of genetically predisposed or induced mouse models of lupus, and the level of circulating UH2A antibodies in the serum of these mice was apparently not linked to the manifestation of lupus nephritis [34, 35]. These observations reflect important differences in the pathophysiological response to components of the ubiquitin family in murine and human lupus disease. Serum IgG antibodies reacting with the branched UH2A peptide were also found to be present in 52% patients with Crohn’s disease (10% had antibodies reacting with ubiquitin) and 10% of patients with ulcerative colitis [36], two autoimmune bowel diseases with a potential evolution toward colon cancer. All together, these data support an important role of ubiquitinated histones in the onset and/or in the development of lupus. To our knowledge, the possible importance of UH2B in lupus has never been investigated.
Poly(ADP-ribosylation) of histones
Antibodies reacting with whole or fragments of poly(ADP-ribose) polymerase (PARP) and with its product poly(ADP-ribose) have been detected in the serum of patients with systemic autoimmune diseases (notably SLE) and with Crohn's disease and ulcerative colitis [36–39]. PARP uses NAD+ as a substrate to synthesize a linear or multibranched polymer of ADP-ribose on glutamic acid, aspartic acid, and lysine residues on various acceptor proteins, including itself. This drastic modification takes place in response to multiple cellular situations, including DNA damage, inflammation, and cell death induction.
Histones H1 and H2B are known substrates of PARP. Although a number of physiopathological effects mediated through the PARP system have been observed in autoimmune diseases, and more specially in SLE, it is not known whether poly(ADP-ribosyl)ated proteins (and histones in particular) are specific targets for autoantibodies. Since we have observed a rearrangement of histones when nucleosomes are incubated with poly(ADP-ribose) chains [40], it may well be that following certain cellular events (and apoptosis for example), poly(ADP-ribosylation) of histones within the nucleosome greatly affects their antigenicity and immunogenicity and leads to an autoimmune response. Other substrates of PARP such as DNA topoisomerase I, a well-known target of autoantibodies in SSc, might also become immunogenic upon modification.
Histone deimination (citrullination) and methylation
A number of autoantibody targets are subject to enzymatic conversion of arginine residues into citrulline. Citrulline is a non-standard amino acid, generated by deimination of arginine residues by calcium-dependent peptidylarginine deiminases (PAD; [41]). The pioneer observation that in RA a very high proportion of patients possess circulating IgG antibodies reacting with citrullinated antigens [42, 43] has generated considerable interest. An intense research led in a few years to the development of a widely used test based on citrullinated peptides validated for the diagnosis of RA and to the identification of deiminated forms of fibrin as major synovial targets for RA antibodies [44]. These findings also led to the conception of new pathological concepts, which might explain the inflammatory process in RA.
Histones are also subject to citrullination. It was found that in histones, human PAD4 converts methylated arginine residues into citrulline [45, 46] (Fig. 1). Hypercitrullination of histones by PAD4 mediates chromatin decondensation [47]. PAD4 targets multiple sites in H3 and H4, including those sites methylated by coactivators CARM1 (Arg17 of H3) and PRMT1 (Arg3 of H4). PAD4 activity was found to be linked to the transcriptional regulation of estrogen-responsive genes in MCF-7 cells [46]. Knowing that methylation of arginine residues within H3 has been linked to active transcription, deimination of methylated arginine residues has been considered as an important mechanism for antagonizing transcription and gene activity. It is not known, however, whether histone citrullination is important for autoantibody recognition. It was recently shown that neutrophils demiminate histones in response to a wide range of inflammatory stimuli arising in infections (e.g., LPS, TNF, or hydrogen peroxide) and not after treatments that induce apoptosis [48]. In parallel, a massive redistribution of chromatin occurs. It was postulated that deiminated histones and bacterial structures that stimulate signaling pathways might break tolerance and initiate immune response that with time could extend to non-modified form of the antigen [48].
Methylation of lysine residues in histones also plays a role in the regulation of transcription and silencing of genes. However, so far, it remains unclear if a change in the methylation status of histones is linked with apoptosis or autoimmunity.
Incorporation of symmetrical dimethylarginine residues in histones
Several members of the Sm/RNP proteins family contain symmetric ω-N G,N G′-dimethylarginine (sDMA) residues. Brahms et al. [49] were the first to identify that SmD1 and SmD3 autoantigens involved in RNA splicing, are dimethylated in vivo in all nine and four positions, respectively, of their C-terminal RG stretches. A diagnostic test based on a single peptide encompassing residues 108–122 of SmD3 containing an additional cysteine residue at the C-terminus and an sDMA residue at position 112 was developed. This test was found to significantly improve the detection of Sm antibodies in lupus sera [50, 51].
Specific enzymes able to introduce sDMA residues into histone H4 and myelin basic protein have been characterized. It is not established, however, whether this modification occurs in vivo and contributes to antigenicity and/or immunogenicity of these self-proteins.
Other histone modifications
Histones are also subject to other modifications such as transglutamination on Gln and Lys residues [52], peroxynitrite modification that in vivo increase the content of carbonyl, nitrotyrosine and dityrosine in histone [53], and incorporation of isoaspartyl moiety, mostly at Asp25 residue in H2B [54]. Although additional studies are required to establish a clear link between these modifications and autoimmunity, they have been shown, as acetylation for example [55], to alter histone immunogenicity and might therefore play an important role in the anti-histone autoimmune response. Excessive protein nitration and in particular hypernitrotyrosinemia has been reported in different inflammation-associated situations, including SLE, RA, SS, and vasculitis [56]. Overexpression of inducible nitric oxide synthase enzyme has been seen in numerous tissues of active patients with lupus.
Conclusions
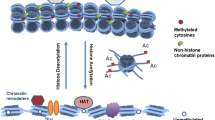
Among the variety of possible histone modifications, only a limited number has directly been associated with autoimmunity (Fig. 1). However, the central question remains how post-translational modified self-proteins can activate the immune system. Evidence shows a strong link between anti-nuclear autoimmunity and cell death. Therefore, several mechanisms, which link cell death to the production of anti-chromatin autoantibodies, have been proposed (Fig. 2). Histone modifications during cell death alter the structure of chromatin. This might lead to the exposure on the outside of chromatin of new epitopes encompassed in histone regions showing an increased immunogenic potential. In addition, histone modifications can enhance the binding of certain molecules. Recently, the chromatin-binding protein HMGB1 has been identified as an important “danger” molecule, involved in the activation of the immune system by dying cells [57]. Chromatin/HMGB1 complexes are found in the serum of patients with SLE and are able to activate immune cells [58]. Cell death is also accompanied by the production of reactive oxygen species (ROS), which directly induce new modifications. ROS-induced histone modifications have not been identified yet, but might increase the antigenicity of histones. Finally, autoimmunity has been associated with an inefficient removal of apoptotic debris [11, 59]. The prolonged survival of apoptotic structures such as (modified) chromatin might lead to the activation of circulating autoreactive immune cells. In total, these mechanisms can lead to the activation of the immune system and lead to the production of anti-chromatin autoantibodies. Finally, as an Editor’s note and for completeness in addition to this special issue on epigenetics, there have been several recent publications, which have focused not only on epigenetics and autoimmunity but also on epigenetics as a developmental origin of a variety of human diseases [60–70].

A schematic overview of the role of histone modifications in the development of anti-chromatin autoimmunity. Shortly, several processes involving cell death by necrosis, apoptosis, or the formation of neutrophil extracellular traps lead to histone modifications, which can subsequently cause an increased immunogenicity of chromatin. Histone modifications change the structure of chromatin, which might result in the exposure of immunogenic epitopes or enhance the binding of danger molecules such as HMGB1. In addition, abnormal modifications may be induced by reactive oxygen species produced during cell death. Finally, a defective/abnormal removal of chromatin released from dying cells could lead to a prolonged persistence of altered material in the circulation. This could all lead to the activation of the immune system and subsequently to the production of anti-chromatin autoantibodies
References
Bhaumik SR, Smith E, Shilatifard A (2007) Covalent modifications of histones during development and disease pathogenesis. Nat Struct Mol Biol 14:1008–1016
Ruthenburg AJ, Li H, Patel DJ, Allis CD (2007) Multivalent engagement of chromatin modifications by linked binding modules. Nat Rev Mol Cell Biol 8:983–994
Mahler M, Blüthner M, Pollard KM (2003) Advances in B-cell epitope analysis of autoantigens in connective tissue diseases. Clin Immunol 107:65–79
Routsias JG, Tzioufas AG, Moutsopoulos HM (2004) The clinical value of intracellular autoantigens B-cell epitopes in systemic rheumatic diseases. Clin Chim Acta 340:1–25
LeFeber WP, Norris DA, Ryan SR et al (1984) Ultraviolet light induces binding of antibodies to selected nuclear antigens on cultured human keratinocytes. J Clin Invest 74:1545–1551
Casciola-Rosen LA, Anhalt G, Rosen A (1994) Autoantigens targeted in systemic lupus erythematosus are clustered in two populations of surface structures on apoptotic keratinocytes. J Exp Med 179:1317–1330
Casiano CA, Tan EM (1996) Recent developments in the understanding of antinuclear autoantibodies. Int Arch Allergy Immunol 111:308–313
Utz PJ, Gensler TJ, Anderson P (2000) Death, autoantigen modifications, and tolerance. Arthritis Res 2:101–114
Rodenburg RJT, Raats HMJ, Pruijn GJM, van Venrooij WJ (2000) Cell death: a trigger of autoimmunity? BioEssay 22:627–636
Rovere P, Sabbadini MG, Fazzini F et al (2000) Remnants of suicidal cells fostering systemic autoaggression. Apoptosis in the origin and maintenance of autoimmunity. Arthritis Rheum 43:1663–1672
Gaipl US, Kuhn A, Sheriff A et al (2006) Clearance of apoptotic cells in human SLE. Curr Dir Autoimmun 9:173–187
Munoz LE, van Bavel C, Franz S, Berden J, Herrmann M, van der Vlag J (2008) Apoptosis in the pathogenesis of systemic lupus erythematosus. Lupus 17:371–375
Dumortier H, Muller S (2007) Histone autoantibodies. In: Shoenfeld Y, Meroni PL, Gershwin E (eds) Textbook of autoantibodies, chapter 22, 2nd edn. Elsevier, Amsterdam, pp 169–176
Cheung WL, Ajiro K, Samejima K et al (2003) Apoptotic phosphorylation of histone H2B is mediated by mammalian sterile twenty kinase. Cell 113:507–517
Monestier M, Fasy TM, Losman MJ, Novick KE, Muller S (1993) Structure and binding properties of monoclonal antibodies to core histones from autoimmune mice. Mol Immunol 30:1069–1075
Monestier M, Decker P, Briand JP, Gabriel JL, Muller S (2000) Molecular and structural properties of three autoimmune IgG monoclonal antibodies to histone H2B. J Biol Chem 275:13558–13563
Fournel S, Muller S (2003) Synthetic peptides in the diagnosis of systemic autoimmune diseases. Curr Prot Peptide Sci 4:261–276
Lu L, Kaliyaperumal A, Boumpas DT, Datta SK (1999) Major peptide autoepitopes for nucleosome-specific T cells of human lupus. J Clin Invest 104:345–355
Kaliyaperumal A, Mohan C, Wu W, Datta SK (1996) Nucleosomal peptide epitopes for nephritis-inducing T helper cells of murine lupus. J Exp Med 183:2459–2469
Sköldberg F, Rönnblom L, Thornemo M et al (2002) Identification of AHNAK as a novel autoantigen in systemic lupus erythematosus. Biochem Biophys Res Commun 291:951–958
Stemmer C, Briand JP, Muller S (1994) Mapping of linear epitopes of human histone H1 recognized by rabbit anti-H1/H5 antisera and antibodies from autoimmune patients. Mol Immunol 31:1037–1046
Dieker JW, Fransen JH, Van Bavel CC et al (2007) Apoptosis-induced acetylation of histones is pathogenic in systemic lupus erythematosus. Arthritis Rheum 56:1921–1933
Batova I, Kowal C, May R, Scharff MD, Diamond B (2008) Human recombinant Fab fragments with sub-nanomolar affinities for acetylated histones. J Immunol Methods 329:1–10
Hu N, Qiu X, Luo Y et al (2008) Abnormal histone modifications patterns in lupus CD4+ T cells. J Rheumatol 35:804–810
Garcia BA, Busby SA, Shabanowitz J, Hunt DF, Mishra N (2005) Resetting the epigenetic histone code in the MRL-lpr/lpr mouse model of lupus by histone acetylase inhibition. J Proteome Res 4:2032–2042
Mishra N, Reilly CM, Brown DR, Ruiz P, Gilkeson GS (2003) Histone deacetylase inhibitors modulate renal disease in the MRL-lpr/lpr mouse. J Clin Invest 111:539–552
Reilly CM, Mishra N, Miller JM et al (2004) Modulation of renal disease in MRL/lpr mice by suberoylanilide hydroxamic acid. J Immunol 173:4171–4178
Marushige Y, Marushige K (1995) Disappearance of ubiquitinated histone H2A during chromatin condensation in TGF beta 1-induced apoptosis. Anticancer Res 15:267–272
Mimnaugh EG, Kayastha G, McGovern NB et al (2001) Caspase-dependent deubiquitination of monoubiquitinated nucleosomal histone H2A induced by diverse apoptogenic stimuli. Cell Death Differ 8:1182–1196
Plaué S, Muller S, Van Regenmortel MHV (1989) A branched, synthetic octapeptide of ubiquinated histone H2A as target of autoantibodies. J Exp Med 169:1607–1617
Muller S, Briand JP, Van Regenmortel MHV (1988) Presence of antibodies to ubiquitin during the autoimmune response associated with systemic lupus erythematosus. Proc Natl Acad Sci USA 85:8176–8180
Ravirajan CT, Kalsi J, Winska-Wiloch H et al (1992) Antigen binding diversity of human hybridoma autoantibodies derived from splenocytes of patients with SLE. Lupus 1:157–165
Stöckl F, Muller S, Batsford S et al (1994) A role for histones and ubiquitin in lupus nephritis? Clin Nephrol 41:10–17
Elouaai F, Lule J, Benoist H et al (1994) Autoimmunity to histones, ubiquitin, and ubiquitinated histone H2A in NZB x NZW an MRL-lpr/lpr mice. Anti-histone antibodies are concentrated in glomerular eluates of lupus mice. Nephrol Dialysis Transpl 9:362–366
Mézière C, Stöckl F, Batsford S, Vogt A, Muller S (1994) Antibodies to DNA, chromatin core particles and histones in mice with graft-versus-host disease and their involvement in glomerular injury. Clin Exp Immunol 98:287–294
Reumaux D, Mézière C, Colombel JF, Duthilleul P, Muller S (1995) Distinct production of antibodies to nuclear components in ulcerative colitis and Crohn’s disease. Clin Immunol Immunopathol 77:349–357
Kanai Y, Kawaminami Y, Miwa M et al (1977) naturally-occurring antibodies to poly(ADP-ribose) in patients with systemic lupus erythematosus. Nature 265:175–177
Muller S, Briand JP, Barakat S et al (1994) Autoantibodies reacting with poly(ADP-ribose) and with a zinc-finger functional domain of poly(ADP-ribose)polymerase involved in the recognition of damaged DNA. Clin Immunol Immunopathol 73:187–196
Decker P, Briand JP, De Murcia G, Pero RW, Isenberg DA, Muller S (1998) Zinc is an essential co-factor for recognition of the DNA-binding domain of poly(ADP-ribose) polymerase by antibodies in autoimmune rheumatic and bowel diseases. Arthritis Rheum 41:918–926
Thibeault L, Hengartner M, Lagueux J, Poirier G, Muller S (1992) Rearrangement of the nucleosome structure in chromatin by poly (ADP-ribose). Biochim Biophys Acta 1121:317–324
Vossenaar ER, Zendman AJ, van Venrooij WJ, Pruijn GJ (2003) PAD, a growing family of citrullinating enzymes: genes, features and involvement in disease. Bioessays 25:1106–1118
Schellekens GA, de Jong BA, van den Hoogen FH, van de Putte LB, van Venrooij WJ (1998) Citrulline is an essential constituent of antigenic determinants recognized by rheumatoid arthritis-specific autoantibodies. J Clin Invest 101:273–281
Girbal-Neuhauser E, Durieux JJ, Arnaud M et al (1999) The epitopes targeted by the rheumatoid arthritis-associated antifilaggrin autoantibodies are posttranslationally generated on various sites of (pro)filaggrin by deimination of arginine residues. J Immunol 162:585–594
Masson-Bessière C, Sebbag M, Girbal-Neuhauser E et al (2001) The major synovial targets of the rheumatoid arthritis-specific antifilaggrin autoantibodies are deiminated forms of the alpha- and beta-chains of fibrin. J Immunol 166:4177–4184
Cuthbert GL, Daujat S, Snowden AW et al (2004) Histone deimination antagonizes arginine methylation. Cell 118:545–553
Wang Y, Wysocka J, Sayegh J et al (2004) Human PAD4 regulates histone arginine methylation levels via demethylimination. Science 306:279–283
Wang Y, Li M, Stadler S et al (2009) Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J Cell Biol 184:205–213
Neeli I, Khan SN, Radic M (2008) Histone deimination as a response to inflammatory stimuli in neutrophils. J Immunol 180:1895–1902
Brahms H, Raymackers J, Union A, de Keyser F, Meheus L, Lührmann R (2000) The C-terminal RG dipeptide repeats of the spliceosomal Sm proteins D1 and D3 contain symmetrical dimethylarginines, which form a major B-cell epitope for anti-Sm autoantibodies. J Biol Chem 275:17122–17129
Mahler M, Stinton LM, Fritzler MJ (2005) Improved serological differentiation between systemic lupus erythematosus and mixed connective tissue disease by use of an SmD3 peptide-based immunoassay. Clin Diagn Lab Immunol 12:107–113
Mahler M, Fritzler MJ, Blüthner M (2005) Identification of a SmD3 epitope with a single symmetrical dimethylation of an arginine residue as a specific target of a subpopulation of anti-Sm antibodies. Arthritis Res Ther 7:R19–R29
Piacentini M, Colizzi V (1999) Tissue transglutaminase: apoptosis versus autoimmunity. Immunol Today 20:130–134
Khan MA, Dixit K, Jabeen S, Moinuddin AK (2009) Impact of peroxynitrite modification on structure and immunogenicity of H2A histone. Scand J Immunol 69:99–109
Young GW, Hoofring SA, Mamula MJ et al (2005) Structural integrity of histone H2B in vivo requires the activity of protein L-isoaspartate O-methyltransferase, a putative protein repair enzyme. J Biol Chem 280:26094–26098
Muller S, Isabey A, Couppez M, Plaué S, Sommermeyer G, Van Regenmortel MHV (1987) Specificity of antibodies raised against triacetylated histone H4. Mol Immunol 24:779–789
Kuo WN, Kreahling JM, Shanbhag VP, Shanbhag PP, Mewar M (2000) Protein nitration. Mol Cell Biochem 214:121–129
Kono H, Rock KL (2008) How dying cells alert the immune system to danger. Nat Rev Immunol 8:279–289
Urbonaviciute V, Fürnrohr BG, Meister S et al (2008) Induction of inflammatory and immune responses by HMGB1-nucleosome complexes: implications for the pathogenesis of SLE. J Exp Med 205:3007–3018
Dieker JW, van der Vlag J, Berden JH (2002) Triggers for anti-chromatin autoantibody production in SLE. Lupus 11:856–864
Arnheim N, Calabrese P (2009) Understanding what determines the frequency and pattern of human germline mutations. Nat Rev Genet 10:478–488
Barros SP, Offenbacher S (2009) Epigenetics: connecting environment and genotype to phenotype and disease. J Dent Res 88:400–408
Figueiredo LM, Cross GA, Janzen CJ (2009) Epigenetic regulation in African trypanosomes: a new kid on the block. Nat Rev Microbiol 7:504–513
Hewagama A, Richardson B (2009) The genetics and epigenetics of autoimmune diseases. J Autoimmun 33:3–11
Invernizzi P (2009) Future directions in genetic for autoimmune diseases. J Autoimmun 33:1–2
Invernizzi P, Pasini S, Selmi C, Gershwin ME, Podda M (2009) Female predominance and X chromosome defects in autoimmune diseases. J Autoimmun 33:12–16
Larizza D, Calcaterra V, Martinetti M (2009) Autoimmune stigmata in Turner syndrome: when lacks an X chromosome. J Autoimmun 33:25–30
Persani L, Rossetti R, Cacciatore C, Bonomi M (2009) Primary Ovarian Insufficiency: X chromosome defects and autoimmunity. J Autoimmun 33:35–41
Sawalha AH, Harley JB, Scofield RH (2009) Autoimmunity and Klinefelter's syndrome: when men have two X chromosomes. J Autoimmun 33:31–34
Wells AD (2009) New insights into the molecular basis of T cell anergy: anergy factors, avoidance sensors, and epigenetic imprinting. J Immunol 182:7331–7341
Zernicka-Goetz M, Morris SA, Bruce AW (2009) Making a firm decision: multifaceted regulation of cell fate in the early mouse embryo. Nat Rev Genet 10:467–477
Acknowledgments
This work was supported by the French Centre National de la Recherche Scientifique (CNRS). JD was supported by a post-doctoral fellowship from CNRS and the French Fondation pour la Recherche Médicale.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dieker, J., Muller, S. Epigenetic Histone Code and Autoimmunity. Clinic Rev Allerg Immunol 39, 78–84 (2010). https://doi.org/10.1007/s12016-009-8173-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12016-009-8173-7