
Abstract
Traditionally, tendons are considered to only contain tenocytes that are responsible for the maintenance, repair and remodeling of tendons. Stem cells, which are termed tendon-derived stem cells (TDSCs), have recently been identified in tendons. This review aims to summarize the current information about the in vitro characteristics of TDSCs, including issues related to TDSC isolation and culture, their cell morphology, immunophenotypes, proliferation and differentiation characteristics and senescence during in vitro passaging. The challenges in studying the functions of these cells are also discussed. The niche where TDSCs resided essentially provides signals that are conducive to the maintenance of definitive stem cell properties of TDSCs. Yet the niche may also induce pathologies by imposing an aberrant function on TDSCs or other targets. The possible niche factors of TDSCs are herein discussed. We presented current evidences supporting the potential pathogenic role of TDSCs in the development of tendinopathy with reference to the recent findings on the altered biological responses of these cells in response to their potential niche factors. The use of resident stem cells may promote engraftment and differentiation of transplanted cells in tendon and tendon–bone junction repair because the tendon milieu is an ideal and familiar environment to the transplanted cells. Evidences are presented to show the potential advantages and results of using TDSCs as a new cell source for tendon and tendon–bone junction repair. Issues pertaining to the use of TDSCs for tissue repair are also discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
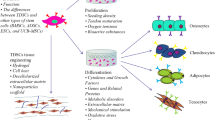
Tendons are traditionally considered to contain only tenocytes, for the maintenance, repair and remodeling of tendons. Despite this, scientific studies have long suggested that there might be a special cell population inside tendons that possesses self-renewal and multi-lineage differentiation potentials [1–5]. However, it was not until 2007 that Bi et al. [6] directly demonstrated the existence of stem cells in tendon tissue. Bi et al. [6] showed that human and mouse tendons contained a minor cell population that exhibited universal stem cell characteristics, including clonogenicity, self-renewal and multi-lineage differentiation capacities even after extended expansion in vitro and in vivo. Since then, there has been extensive progress and interest in the study of these cells in tendon physiology, pathology and tendon tissue engineering. This review therefore aims to present the current evidences about the in vitro characteristics of TDSCs and the challenges in studying the functions of these cells. TDSCs, similar to other stem cells, require the interplay of their local micro-environment (‘niche’) to participate in tissue regeneration, maintenance and repair. The in vivo niche of TDSCs is not clearly understood at present. Evidences about the possible niche factors of TDSCs are herein presented. The niche may go wrong and induce aberrant functions of TDSCs. Evidences supporting the potential pathological role(s) of TDSCs and their potential in vivo niche factors in the development of tendinopathy are discussed. The potential use and issues pertaining to the use of TDSCs as a new cell source for tendon and tendon–bone junction repair are also presented.
Slightly different terms have been used in the literature to describe these stem and progenitor cells isolated from tendons. In this review, we will use the term tendon-derived stem cells (TDSCs) to represent this cell population.
In Vitro TDSC Characteristics
TDSC Isolation and Culture
TDSCs have been isolated and cultured from adult human [6], mouse [6], rabbit [7], rat [8] and fetal human [9] tendons. To isolate TDSCs, the tendon tissue is minced and digested with collagenase type I. The isolated nucleated cells are then plated at an optimal low cell density. TDSCs attach to the plate and remain quiescent for several days before they divide rapidly to form colonies [9]. The current stem cell culture method depends on the selection and enrichment of stem cells during sub-culture. Zhang and Wang et al. [7, 10, 11] removed tenocytes by picking TDSC colonies by local application of trypsin. The TDSC colonies were pooled and sub-cultured for experiments. This method has the advantage of reducing the contamination by tenocytes but the efficiency of getting a pure TDSC culture needs more experimentation. The selection of an optimal seeding density is a critical issue in successful TDSC culture. TDSCs, like other stem cells, grow faster at a very low seeding density [8, 12, 13]. There are species-specific and tissue origin-specific differences in the optimal initial seeding density [8, 14, 15].
The size and density of the TDSCs colonies were mixed, suggesting a large variation of cell proliferation among colonies [7, 8]. The chondro-, osteo- and adipogenic differentiation potentials of TDSC colonies also varied [6]. The TDSCs obtained from tendon tissues are therefore most likely a mixture of stem cells and progenitor cells, which are heterogeneous in self-renewal potential, proliferation and multi-lineage differentiation potentials.
The age of the animal or the individual might affect the success of obtaining TDSCs; indeed, Zhou et al. [16] reported that there was a depletion of the TDSCs pool in aged tendon tissues, with a 70% reduction in the number of TDSCs isolated from aged (24–26 months) (0.171 ± 0.06%) compared to young (3–4 months) tendons (6.26 ± 0.55%). As a result, the isolation of TDSCs from young individuals is preferred. Human hamstring tendons and mouse patellar tendons contain 3–4% of TDSCs [6]. Rat flexor tendons contain about 1–2% of TDSCs [8]. About 5–6% of human fetal Achilles TDSCs at passage 2 were able to form colonies [9]. While it only represents a very small fraction of total nucleated cells, the value has already exceeded the percentage of stem cells in human bone marrow aspirates by at least three orders of magnitude (0.001–0.01% of total nucleated cells) [17, 18]. There is currently a lack of uniformity on the appearance of TDSCs, and the shape is largely dependent on the species, tissue origins, cell passages and confluence of the culture. Figure 1 summarizes the different morphologies of TDSCs isolated from different species and tissue origins.

Photographs showing the different morphologies of TDSCs isolated from different species and tissue origins. a Rabbit patellar TDSCs and b rabbit Achilles TDSCs at P10 were reported to be cobblestone-like in confluence culture (Bar: 50 μm). (Reprinted from Zhang. J,, Wang, J. H. C. Characterization of differential properties of rabbit tendon stem cells and tenocytes. BMC Musculoskeletal Disorder, 11, 10, 2010, with permission under terms of the Creative Commons Attribution License for Open Access article). c Human fetal hamstring TDSCs at P2 were reported to be fibroblast-like in culture (Bar: 200 μm) (Reprinted from Yin, Z., Chen, X., Chen, J. L., et al. The regulation of tendon stem cell differentiation by the alignment of nanofibers. Biomaterials, 31(8), 2163–2175, 2010, with permission from Elsevier). d Rat flexor TDSCs at P0-P3 were reported to exhibit different morphologies at different passages. At P0, large polygonal and star-shaped cells were observed while flat cells and slender fibroblast-like cells were observed at P1. At P3, homogeneous populations of fibroblast-like cells were observed. Magnification: ×100; Stain: crystal violet (Reprinted from Rui, Y. F., Lui, P. P. Y., Li, G., Fu, S. C., Lee, Y. W., Chan, K. M. Isolation and characterization of multi-potent rat tendon-derived stem cells. Tissue Engineering Part A, 16(5), 1549–1558, 2010. No permission required from Mary Ann Liebert, Inc. publishers for reusing authors’ own content). e Mouse patellar TDSCs (called mTSPCs in the original article [6]) were reported to be displayed five different morphologies and were morphologically different from that of BMSCs, whereas human hamstring TDSCs (called hTSPCs in the original article [6]) were relatively homogeneous and were morphologically similar to that of human BMSCs (Bar: 100 μm) (Reprinted by permission from Macmillan Publishers Ltd: Nature Medicine, Bi, Y., Ehirchiou, D., Kilts, T. M., et al. Identification of tendon stem / progenitor cells and the role of the extracellular matrix in their niche. Nature Medicine, 13(10), 1219–1227, 2007.) f Two types of cells were identified during in vitro culture of patellar TDSCs from P5 to P30. One of them was spindle-shaped, showing a fibroblastic appearance while the other one displayed an enlarged and triangular morphology. Magnification: ×100; Stain: crystal violet (unpublished result)
Immunophenotypic Prolife
Phenotypically, TDSCs share some common stem cell markers as identified on the cell surface of other MSCs. They were reported to express Sca-1, CD44, CD90, CD90.1, CD90.2, CD105, Stro-1, CD146, nucleostemin, Oct-4 and SSEA-4, but were negative for CD31, CD34, CD18, CD117, CD45, Flk-1, CD144 and CD106 [6–11, 16]. There were differences in the expression of stem cell markers in young and aged rat TDSCs [16]. Whereas nearly 100% both of young and aged rat TDSCs expressed nucleostemin, Oct-4 and SSEA-4, aged TDSCs expressed a lower level of CD90.1 than young cells but a higher level of CD44 [16]. The authors hypothesized that the increase in CD44 expression in aged TDSCs might contribute to reduced repair capacity of TDSCs with age as CD44 knock-down was reported to improve healing tendon mechanics, and to increase matrix and cytokine expression in a mouse patellar tendon injury model [16]. Table 1 summarizes the expression of phenotypic markers in TDSCs isolated from different species.
Although MSCs expressed a number of phenotypic markers such as CD44 and CD90, none of which unfortunately is specific to MSCs. The expression of a unique and definitive surface antigen on MSCs that supports the specific identification of MSCs is yet to be identified. The isolation of MSCs, including TDSCs, remains largely dependent on a selection and enrichment approach. Whereas TDSCs expressed many of the same markers as BMSCs, the expression patterns were not identical (see also Section “Potential Applications of TDSCs in Musculoskeletal Tissue Repair”). Both mouse and human TDSCs lacked CD18 which was reported to be expressed by BMSCs [6, 9]. On the other hand, over 60% of mouse TDSCs expressed CD90.2, whereas CD90.2 was absent in mouse BMSCs in Bi et al.’s study [6]. CD106 was reported not to express in human and rat TDSCs [6, 8, 9, 16] but it was reported to express in human and mouse BMSCs in other studies [19, 20]. These data suggested that TDSCs are closely related to BMSCs, but not identical to them. However, the possibility that TDSC and BMSC are different stages of a common MSC cannot be disregarded.
The markers that can differentiate TDSCs and tenocytes also remain largely undefined. Zhang and Wang [7, 10] reported that TDSCs expressed Oct-4, SSEA-4 and nucleostemin, whereas tenocyes expressed none of these markers. While TDSCs possess multi-lineage differentiation potential, tenocytes did not show such differentiation potential [7]. The identification of a minimum set of criteria that can differentiate TDSCs from BMSCs and tenocytes would facilitate the study of their in vivo niche, roles in tendon physiology and pathology as well as applications in tissue engineering.
Cell Proliferation
TDSCs, similar to other MSCs, proliferated faster than the terminally differentiated cells in culture [7]. Young TDSCs were also reported to proliferate faster than aged TDSCs, which might be caused by cell cycle arrest at the G2/M phase in aged TDSCs [16].
Differentiation Potential
TDSCs were able to differentiate into adipocytes, chondrocytes and osteocytes in vitro [6–9] and formed tendon-like, cartilage-like, bone-like and tendon–bone junction-like tissues in nude mouse/rat models [6, 7]. There were age-related declines in the mRNA expressions of scleraxis (Scx) and tendmodulin (Tnmd), two tendon lineage-related molecular markers in TDSCs both at basal level and upon induction by TGF-beta3 [16]. As the specificity of these markers for tenocyte identification remains unclear, whether their decreased expression indicated reduced tenogenic differentiation ability of aged TDSCs to tenocytes compared to young TDSCs needs further study. While aged TDSCs showed no difference in osteogenic and chondrogenic differentiation potentials compared to their young counterpart, they formed adipocytes more readily and expressed higher levels of adipogenic markers including PPARγ2, C/EBPa and leptin than the young TDSCs following induction [16].
MSCs are initially defined as multi-potent adult stem cells that can differentiate into different tissues originated from the mesoderm. However, recent data showed that MSCs could be pluripotent and could differentiate into tissues and cells of non-mesodermic origin, such as hepatocytes, neurons, cardiomyocytes and epithelial cells [21–27]. Whether TDSCs can also differentiate into cells of non-mesodermic origin or are developmentally adapted to differentiate into tissue in which they resided need further study. This information is important for gaining further insight into the lineage hierarchy of stemness property of TDSCs and hence has important implications in the application of this new cell type for tissue engineering.
Senescence During In Vitro Passaging
Although MSCs show remarkable self-renewal potential, and can be readily expanded in vitro by serial passaging, they are vulnerable to replicative senescence [28–35]. The population doubling time of rabbit patellar and Achilles TDSCs increased at the late passages (>P12), suggesting senescence of the cells during sub-culture [7]. Although the colony numbers and the proliferative rate of rat patellar TDSCs increased during sub-culture (up to P30), their senescence associated β-galactosidase activity increased while their stem cell-related marker expression and multi-lineage differentiation potential decreased with in vitro passaging [36, unpublished result].
Physiological Niche
Adult stem cells have limited functions without the niche. An example is haematopoietic stem cells (HSCs) which function to regenerate the blood and the immune system. They have little function outside their specific anatomic locations and their functions are constantly modulated by niche signals under conditions of physiological challenge to meet the needs of an organism [37]. The interplay between stem cells and their niche creates a dynamic system necessary for sustaining tissues. However, the niche may also induce pathologies by imposing an aberrant function on stem cells or on other targets. By modifying the stem cell niche or by replicating the in vivo stem cell niche during in vitro stem cell culture, we may be able to design new stem cell therapeutics for the treatment of disease. Understanding the stem cell niche is therefore crucial in stem cell biology because it contributes both to tissue health and diseases.
The same probably also applies to TDSCs which also require the niche to maintain their definitive stem cell properties. As stem cells residing in tendons, TDSCs are expected to play a primary role in maintaining tissue homeostasis and repair after injury. Diseases, such as chronic tendinopathy, therefore, may occur if the control of renewal and differentiation of these resident stem cells becomes aberrant. The organization and exact niche signals which regulate TDSC fate are not clearly understood at present. However, mechanical loading, composition and nano-/micro-structure of extracellular matrix (ECM), biological factors, tenocytes, neural input, vascular input, physiological factors such as oxygen tension and metabolic products of tissue activity, among others, are likely to be important niche components which regulate TDSC fate (Fig. 2). Studies on the effects of niche factors on TDSC’s renewal, proliferation and differentiation will therefore provide important information about tendon physiology, pathology and also allow us to recapitulate the condition for expansion of TDSCs in vitro for tissue engineering. In this regard, TDSCs are sensitive to mechanical loading, which features an inherent part of the tendon environment [14, 38, 39]. While the exact composition of ECM of TDSCs remained unclear, a previous study has shown that slow cycling cells, supposed to be TDSCs, are resident in-between long parallel collagen fibrils containing biglycan and fibromodulin [6] because the depletion of biglycan and fibromodulin in a double knock-out animal model impaired tendon formation, which was more translucent, significantly thinner and more cellular than the wild-type patellar tendon. The tendon tissue also showed disorganized collagen fibers with large gaps and pathological ossification within the tendon tissue [6]. The interaction of TDSCs with an aligned versus random nanofibrous scaffold was reported to control their tenogenic versus osteogenic lineage commitment [9], suggesting that TDSCs are sensitive to topographical cues from the ECM. The cytoskeletal structure of TDSC may mediate its interaction with the ECM, as shown in Yin et al.’s study [9]. We have previously reported the clustering of oval-shaped cells in tendons which displayed high proliferative index and apoptotic index as well as expressed high levels of pro-collagen type 1, TGF beta 1, hsp47, BMP-12 and BMP-13 [40]. We termed this cell cluster as an active remodeling site in tendons; and we also suggested that it served as the nucleus for intrinsic healing responses in normal tendons in response to micro-injuries from daily activities. Whether this active remodeling site is TDSCs needs further study [40]. The TDSC fate is also likely to be controlled by the BMP signaling pathway; this is because TDSCs isolated from the biglycan and fibromodulin double knock-out animal model displayed increased sensitivity to BMP-2 signaling [6]. A molecular defense mechanism against the BMP signaling pathway to prevent chondro-osteogenesis in tendon mid-substance is therefore likely to be in place [41, 42]. Tenocytes are the major cell type in tendon and form a three-dimensional network of cell processes throughout tendon [43]. It is therefore likely that TDSCs interact with tenocytes which modulate their cell fate. Indirect co-culture of rat BMSCs and tendon fibroblasts was reported to promote the proliferation and mRNA expression of tendon/ligament-related genes in BMSCs [44]. Whether tenocytes will also regulate the self-renewal, proliferation and differentiation of TDSCs needs further study. The tendon milieu is hypoxic. Comparison of rat TDSCs cultured in hypoxic (2%) versus normoxic conditions (20% oxygen) showed that their clonogenicity and cell number were higher in the former, suggesting that hypoxia enhanced the recruitment and proliferation of TDSCs (unpublished results). Previous studies suggested that the perivascular niche is a source of stem cells. Perivascular cells were reported to express both pericyte and MSC markers in situ, exhibit clonogenicity and multi-lineage differentiation potential [45]. Some researchers suggested that all MSCs were pericytes that gradually assumed tissue-specific phenotypes under the influence of the local niche [46] because the characteristics of MSCs originating from different tissues were found to be very similar but with varied growth kinetics and functional roles [47]. Perivascular cells of human supraspinatus tendon capillaries expressed both tendon cell markers (Scx, collagen type I, Collagen type III, smad8) and stem/precursor cell markers (CD133, Musashi-1, Nestin, CD44, CD29) in addition to the pericyte-associated marker α-SMA, suggesting that the perivascular niche is a source for tendon precursor cells [48]. BMP-13, a growth factor that could induce the formation of tendon and ligament tissues in an animal study [49], was detected exclusively in active tenoblasts in active remodeling sites and perivascular mesenchymal cells, but not in interstitial tenocytes of human patellar tendons, and it showed similar distribution as proliferating cell nuclear antigen (PCNA) and pro-collagen type I in the tissue [50]. Whether TDSCs are pericytes or resident at the perivascular niche need further study. Rat TDSCs expressed α-SMA, suggesting their possible relationship to perivascular cells [8]. However, it should be noted that tendon is relatively avascular and receives its blood supply mainly from the endotendon and paratendon [51], whereas TDSCs were isolated from the tendon mid-substance after removing all the peritendinous connective tissues [8, 14]. Indeed, the stem/precursor cell-related markers such as nestin and Musashi-1 were expressed in tendon cells embedded in the dense ECM besides the perivascular region [48].

Schematic diagram showing the potential niche factors regulating TDSC fate. Erroneous differentiation of TDSC as a result of change in its niche may detour the fate of TDSC from tenogenesis and affect the size of TDSC pool for tendon repair
Below we will present the current evidences supporting the role(s) of TDSCs and their potential niche factors in chronic tendinopathy, the application of these cells in tissue engineering and the opportunities for mass expansion or tenogenic differentiation of TDSCs through the incorporation of these potential in vivo niche factors in the in vitro culture system.
TDSCs in Chronic Tendinopathy
Evidence for Erroneous Differentiation of TDSCs in the Pathogenesis of Chronic Tendinopathy
Chronic tendinopathy refers to a broad spectrum of pathological conditions in tendons and their insertions, with symptoms including tenderness, swelling and activity-related chronic pain. Its underlying pathogenesis is poorly understood and treatment is usually palliative. A change of tendon loading caused by mechanical overload, compression or disuse has been implicated as the possible etiologies [52], but they do not completely explain the failed healing responses, cellular and molecular alternations seen in the diseased tendon, including hypercellularity, hypervascularity, matrix disturbance with an increase in proteoglycan deposition, particularly the oversulfated form, ECM degradation, rounding of cell nuclei and acquisition of chondrocyte phenotypes, occasional adipose and bony metaplasia [53–61]. The production of abnormal matrix components (e.g., fatty degeneration, glycosaminoglycan accumulation with cell rounding and acquisition of chondrocyte phenotypes and calcification) in tendinopathic tendons suggested either that non-tenocytes migrated to the injury site; or that endogeneous or exogeneous stem cells possessing multi-lineage differentiation potential differentiated into non-tenocytes. The current evidence supported the latter because stem cells possess migratory abilities. Because TDSCs are isolated from tendons and tenocytes were demonstrated not to possess differentiation potential [7], the erroneous differentiation of TDSCs, because of alteration of their in vivo niche as a result of changes of tendon loading, to non-tenocytes, might result in lipid accumulation, ectopic chondro-ossification and mucoid degeneration as well as depletion of the TDSCs pool for tendon repair. Consequently, it might result in pain and failed healing, and it might also predispose the injured tendon to rupture, as seen in tendinopathy. This hypothesis is accepted by the scientific community [62] and is also supported by other groups [6, 7, 38]. Indeed, the ectopic chondro-ossified deposits in the tendon mid-substance appeared in a process resembling endochondral ossification as shown by immunohistochemical staining of collagen type X in our collagenase-induced failed healing animal model of tendinopathy [63]. This is similar to the endochondral ossification process observed in Achilles and patellar tendinopathies [61]. We also observed an earlier expression of chondrogenic markers (collagen type II and Sox9) in the healing tendon fibroblasts prior to their expression in the chondrocyte-like cells and ossified deposits which appeared later in the tendon mid-substance in our animal model [63]. The hypothesis was further supported by the fact that TDSCs which were isolated from the tendon mid-substance in our animal model at week 2 exhibited lower proliferative potential, higher yield and higher basal and induced osteogenic differentiation potentials when compared to TDSCs isolated from healthy tendons [64], although the source of stem cells contributing to the altered functions can be endogeneous or exogeneous as there was no specific markers for TDSCs. Bi et al. [6] also reported that TDSCs isolated from the biglycan and fibromodulin double knock-out animal model showed higher clonogeneity and proliferation. TDSCs isolated from the double knock-out animals expressed collagen type II and aggrecan, which were not observed in wild-type TDSCs [6]. There was decreased protein expression of collagen type I and mRNA expression of Scx in TDSCs isolated from double knock-outs compared to TDSCs isolated from wild-type animals. Moreover, TDSCs isolated from the double knock-out animals formed bone in addition to tendon-like tissue in vivo, whereas wild-type TDSCs only formed tendon-like tissue [6]. Overuse was also reported to upregulate the expression of cartilage-associated genes and downregulate the expression of tendon-associated genes in rat supraspinatous tendons [65] and in horse superficial digital flexor tendons [66]. Indeed, aberrant differentiation of stem cells is also seen in the pathogenesis of arterial calcification [67], skin calcification [68], skeletal calcification [69] and fracture nonunion [70].
Alteration of Potential TDSC Niche Factors
Other supportive evidences for the role(s) of erroneous differentiation of TDSCs in the pathogenesis of chronic tendinopathy come from altered biological responses of these cells in response to mechanical loading, biological factors, composition and micro-/nanostructure of ECM, which are potential important niche factors of TDSCs in vivo.
Mechanical Loading
TDSCs were sensitive to repetitive tensile mechanical loading in vitro. Cyclic uniaxial stretching was reported to align the cells along the direction of mechanical load dose-dependently [14, 38] and increase the proliferation of TDSCs seeded in microgrooves in a stretching magnitude-dependent manner [38]. Treadmill running at 13 m/minute for 50 min a day, 5 days a weeks for three weeks doubled the proliferation rates (shortened population doubling time and increased nucleostemin expression) and hence expanded the pool both of mouse Achilles TDSCs and patellar TDSCs as well as increased the production of collagen in Achilles TDSCs- and patellar TDSCs-tenocytes co-cultures [10]. However, large mechanical loading might be detrimental because 8%, but not 4%, stretching directed the differentiation of TDSCs into non-tenocyte lineages [10]. This finding provides a possible mechanistic link between erroneous differentiation of TDSCs and mechanical overloading, a commonly suggested etiological factor of chronic tendinopathy. However, the loading conditions tested were limited in all the previous studies. Whether mechanical strain, frequency or duration is more important in defining mechanical overload and hence more detrimental to tendon homeostasis remains unclear. Future studies should test a wider range of loading conditions to define more precisely how different loading regimens influence TDSCs proliferation, differentiation and self-renewal. There is a lateral compressive component while applying uniaxial tensile loading during stretching of plates or tendon loading. Therefore, it is not absolutely clear if it is the tensile or compressive strain which TDSCs are responding. Future study is required to find out the exact signal that TDSCs are responding. The response of stem cells in 3D culture was different [71]. The stretching of TDSCs in 3D culture embedded in different extracellular matrices should be investigated in order to more closely examine the mechanobiological responses of TDSCs. The inclusion of tenocyes will produce a more physiologically relevant experimental condition for evaluating the responses of TDSCs in response to mechanical loading. The mechanotransduction mechanism leading to altered TDSC differentiation should also be investigated.
Biological Factors
Previous literature has shown that there was an increased expression of COX-2 and PGE2 in clinical samples of tendinopathy [55]. Moreover, the production of PGE2 increased in tendons during exercise in animals [72, 73] and the production of COX-1, -2 and PGE2 also increased in human tendon fibroblasts after repetitive mechanical loading in vitro [74, 75]. Repeated intratendinous injection of PGE2 was reported to lead to focal hypercellularity and matrix degeneration in the patellar tendon of rabbits similar to those seen in tendinopathy [75]. As such, PGE2 might be a possible contributor to the pathogenesis of tendinopathy. Indeed, a recent study has shown that PGE2 dose-dependently reduced the proliferation while it increased both adipogenesis and osteogenesis of TDSCs, as shown by the accumulation of lipid droplets and calcium deposits, respectively [72]. However, the fact that PGE2 production was increased by physical activity does not necessarily imply that it plays a role in the pathogenesis of tendinopathy. In Zhang and Wang’s study [72], although there was an increased production of PGE2 in tendons after exercise, the mice were run only once until exhausted (mean time: 212 ± 50 min.) and hence could not represent the chronic mechanical loading condition as observed in tendinopathy. Moreover, running at the same speed for 3 weeks was reported to show beneficial effects on TDSCs in another study by the same group [10].
Chondro-osteogenic BMPs, such as BMP-2, BMP-4, BMP-6 and BMP-7, are growth factors belonging to the TGF-β superfamily and have been used in many studies to promote cartilage, bone and tendon–bone junction repair [5, 76–81]. Because of the role of these BMPs in bone and cartilage formation, it is usually found in bone and cartilage and is generally absent in soft tissue such as tendons. However, ectopic expression of chondro-osteogenic BMPs could occur in pathological conditions and lead to chondro-ossification of soft tissues including ligaments and tendons [67, 82–85]. Injection of rhBMP-2 induced ectopic bone formation in tendons [5]. By using the collagenase-induced failed healing animal model of tendinopathy, we reported formation of ectopic tendon–bone junction in the tendon mid-substances with high levels of expression of chondro-osteogenic BMP-2, -4, -7 [63, 86, 87]. Ectopic overexpression of these BMPs was also observed in our clinical samples of patellar tendinopathy [88]. Indeed, the chondro-osteogenic BMPs appeared at the healing tendon fibroblasts prior to their expression at the pathological chondro-ossification sites in the tendon mid-substance, similar to the spatial and temporal expression patterns of chondrogenic markers in the same animal model [87]. When mouse TDSCs were treated with BMP-2 and then transplanted subcutaneously into immunocompromised mice, structures similar to osteotendinous junctions (termed entheses) were formed [6], which were similar to the ectopic chondro-ossified structures observed both in our animal model [63] and in clinical samples of tendinopathy [88]. These data suggested a potential role of these chondro-osteogenic BMPs in chondro-ossification and failed healing process of tendinopathy although no causal relationship could be demonstrated, similar to the case of PGE2. The change of tendon loading might contribute to the ectopic expression of chondro-osteogenic BMPs in tendinopathy. In vitro cyclic tensile loading increased the expression of BMP-2 which promoted osteogenic differentiation of TDSCs [14]. This study provided insights about the potential targets and actions of chondro-osteogenic BMPs in response to mechanical load in the pathogenesis of tendinopathy [14]. In addition to tensile loading, the expression of BMPs was reported to be regulated by hydrostatic pressure, shear stress and compression in other cell types [89–91]. Whether there will be increased ectopic expression of chondro-osteogenic BMPs in TDSCs under compressive loading in vitro, which has also been suggested as an etiological factor of tendinopathy as a result of tendon overuse [52], needs further study.
Composition and Micro-/Nanostructure of Extracellular Matrix
The changes in the composition of ECM might also affect the cellular response of TDSCs and promote their aberrant differentiation into non-tenocytes. We have reported changes in the ECM composition with an increase in proteogylcans (PG) content in the collagenase-induced failed healing animal model of tendinopathy and an increase in sulphated glycosaminoglycans (GAGs) content in clinical samples of tendinopathy [60, 92]. GAGs have been implicated in the modulation of osteogenic bioactivity of BMPs and the reduction of sulphated GAGs in PG on BMSCs surface reduced the effect of endogenous BMPs and exogenous BMP-2 on osteogenic gene expression and alkaline phosphatase (ALP) activity of BMSCs [93]. Whether the increase in GAG content in tendinopathy will increase the effect of endogenous and exogenous BMPs on osteo-chondrogenic differentiation and reduce their effect on tenogenic differentiation of TDSCs need to be investigated. TDSCs isolated from biglycan and fibromodulin double knock-out mice which showed tendon ossification were more sensitive to BMP-2 stimulation with increased phosphorylation of Smad1/5/8 and more abundant nuclear localization of phosphorylated Smad1/5/8 than that in wild type cells [6]. It is known that decorin, fibromodulin and biglycan could modulate the activity of resident cell population by binding to and sequestering growth factors such as TGF-beta [94, 95] and there was an increase in the expression of TGF-beta in clinical samples of patellar tendinopathy [55]. The release of growth factors from the damaged ECM may potentially affect TDSC fate. ECM degradation is observed in tendinopathy [96]. Homeostasis of the native mechanical properties and composition of the matrix are important in maintaining stemness properties of stem cells. Whether and how the loss of ECM alignment and hence mechanical loading on the cells will negatively affect the self-renewal and differentiation of TDSCs need further research [9].
Other Potential Niche Factors
TDSCs are unlikely to be the only cell type involved in the pathogenesis of tendinopathy. Despite the fact that tenocytes were not capable of differentiating into non-tenocyte lineages [7], they might also be involved in the development of tendinopathy through direct interaction with TDSCs or production of inflammatory mediators [74, 97, 98] and tissue degradative enzymes (matrix metalloproteinases, MMPs) [99], which might in terms affect TDSC functions. The interaction between TDSCs and tenocytes remains undefined. Better understanding about their interaction, under various conditions, will improve our understanding of tendon homeostasis and the development of tendinopathy.
Nociceptive substances including substance P (SP) and calcitonin gene-related peptide (CGRP) were reported to increase in clinical specimens; and they were implicated in the pathogenesis and origin of pain in tendinopathy [100–102]. We reported increased expression of SP and CGRP in chondrocyte-like cells and ossified deposits, in addition to healing tendon fibroblasts, in our collagenase-induced failed healing animal model of tendinopathy. Therefore, both might have roles in ectopic chondro-ossification in addition to pain mediation [94]. This was supported by previous studies showing that SP and CGRP promoted osteogenic differentiation of bone marrow cells and osteoblasts [95, 103]. Whether they have any direct effect on TDSCs differentiation requires further experimentation.
TDSCs for Tissue Repair
Potential Applications of TDSCs in Musculoskeletal Tissue Repair
Both acute and chronic tendon injuries are very common during sports activities [104]. Injured tendons heal slowly and often result in the formation of inferior fibrotic scar tissue or fibrous adhesions, which compromise the structure and function of healed tendons and increase the risk of re-injury at the repair site [105–107]. The restoration of normal tendon structure and function after injury represents a challenging area in orthopedic and sports medicine. Recent studies have used stem cell therapy for tendon repair and have shown positive results [108–117]. However, the use of embryonic stem-cells-derived MSCs (ESCs-MSCs) [117] for tendon repair may have the risk of terotama formation, while the use of BMSCs for tendon repair may have the risk of ectopic bone [109, 118] and tumor formation under special circumstances [119].
Although stem cells isolated from different tissues share many important stem cell characteristics including adherence to plastics, clonogenicity and multi-lineage differentiation potentials, the degrees of proliferation and differentiation potentials of stem cells can be affected by their origins [120, 121]. This has implications in the selection of an appropriate cell source and conditions for engineering of specific tissue type [122–131]. Indeed, TDSCs retain some tissue-specific differentiation properties in addition to the universal stem cell characteristics, which may make them more advantageous than BMSCs in musculoskeletal tissue engineering. Bi et al. [6] reported that mouse TDSCs expressed higher mRNA levels of Scx, Comp, Sox9 and Runx2 as compared to mouse BMSCs, whereas human TDSCs expressed a higher level of tenomodulin (TNMD) than human BMSCs. We also reported higher mRNA expression of tenogenic, chondrogenic and osteogenic markers in rat TDSCs compared to paired rat BMSCs at basal level [132].
We further showed that TDSCs excelled BMSCs in osteogenesis, chondrogenesis and tenogenesis upon induction [132, unpublished results]. Bi et al. [6] also reported that murine and human TDSCs accumulated Ca2+ more rapidly (about four times) and formed more calcium nodules compared to BMSCs upon osteogenic induction. Oil red O staining of lipid droplets within the adiopocytes was also greater in human and mouse TDSCs compared to that in BMSCs after adipogenic induction.
Moreover, TDSCs exhibited higher clonogenicity and proliferated faster compared to paired BMSCs [132]. Bi et al. [6] also reported that both human and mouse TDSCs proliferated faster than BMSCs isolated from the same patient or animal. The number of population doublings in mouse TDSCs was also higher than that in BMSCs, but this was not observed for human TDSCs [6].
These results thus suggested that TDSCs may be a promising therapeutic cell source for better and earlier musculoskeletal repair, including tendon repair, compared to BMSCs. Indeed, we found that TDSCs promoted earlier and better tendon repair histologically, biomechanically and ultrasonographically, compared to the fibrin-glue only controls, up to week 4 in a patellar tendon window injury rat model [133]. No ectopic bone formation was observed up to week 4 [133]. Preliminary results showed that the histological and biomechanical healing outcomes at week 4 were at least as good as the case with BMSC transplantation [134]. More studies are required to understand the long-term effect of TDSCs compared to BMSCs and also the fate of the transplanted TDSCs during tendon repair. Because TDSCs showed higher levels of chondrogenic and osteogenic markers as well as chondrogenic and osteogenic differentiation potentials, compared to BMSCs, they are also a potential attractive candidate for tendon–bone junction regeneration. We reported that TDSCs expressed higher protein and mRNA levels of BMP receptors IA, IB and II compared to BMSCs [15]. They also exhibited higher osteogenic activity with and without BMP-2 induction compared to BMSCs [15]. Therefore, TDSCs with/without BMP-2 might be an appealing approach for tendon–bone junction repair. Bi et al. [6] reported that treatment of mouse TDSCs with BMP-2 followed by subcutaneous transplantation of the cells in hydroxylapatite/tricalcium phosphate (HA/TCP) scaffold into immuno-compromised mice led to the formation of tendon–bone junction-like structure. The effect of TDSCs, with or without BMP-2, in the promotion of tendon–bone junction repair should be studied in an injury animal model.
Tendons are relatively cell-poor. An efficient, high-yield process is required to obtain enough TDSCs for tissue engineering. Strategies to promote the in vitro expansion of TDSCs are therefore essential. Exploring the TDSC niche factors will provide opportunities for replicating the in vivo niche for the optimal culture of TDSCs in vitro for regenerative therapies.
Tenogenic Differentiation of TDSCs
Because ectopic bone and tumor formation were reported in previous studies with BMSC transplantation [109, 118, 119], the in vitro differentiation of stem cells towards tenogenic lineage before transplantation might be a good strategy to promote tendon healing while minimizing the chance of ectopic bone and tumor formation in tendons. However, a method of controlling the differentiation of MSCs to tendon-forming cells and avoiding ectopic bone and tumor formation remains a great challenge which hinders their application. Platelet-rich plasma-clot releasate (PRCR) was reported to dose-dependently promote the tenogenic differentiation of rabbits TDSCs and activate the resultant cells as shown by the decrease in nucleostemin protein expression, the increase in mRNA and protein expressions of tenocyte-related markers (collagen type I, collagen type III and tenascin C), the increase in total collagen production as well as the increase in mRNA and protein expressions of α-SMA [11]. The culture of human fetal TDSCs in an aligned nanofibrous scaffold drove their tenogenic commitment through an integrin- and myosin-mediated mechanotransduction pathway while suppressed their osteogenic differentiation [9]. It is not clear whether tenogenic differentiation of TDSCs in vitro will promote better tendon healing compared to undifferentiated TDSCs; experiments are now underway.
Conclusion
In conclusion, we have summarized the in vitro characteristics of TDSCs. The identification of a panel of unique and definitive markers for TDSCs is essential to differentiate them from other stem cells and tenocytes. The spectrum of differentiation potential of TDSCs needs to be further investigated, which may intensify their use in tissue engineering. TDSCs are unlikely to function alone. The stem cell niche regulates how TDSCs participate in tissue regeneration, maintenance and repair. Yet the niche may also induce pathologies by imposing aberrant functions on TDSCs or other targets. The organization and exact niche signals regulating TDSC function are not clearly understood at present. We presented evidences suggesting the erroneous differentiation of TDSCs, as a result of change in the potential niche signals, may contribute to the pathogenesis of chronic tendinopathy. Further investigations of what is the anatomical location of TDSCs, what are the niche signals and how do the niche signals regulate the fate of TDSCs in normal condition and in response to tendon injury will confirm this speculation and provide new treatment directions. TDSCs possess many excellent properties compared to BMSCs. They may be used as a new cell source for musculoskeletal repair, including tendon and tendon-bone junction repair. More studies are required to understand the healing outcome and fate of TDSCs after transplantation in different animal models. Issues pertaining to the use of TDSCs for tissue engineering including cell number and tenogenic differentiation need to be tackled. By replicating the in vivo TDSC niche during in vitro cell culture, we may be able to optimize the culture of TDSCs in vitro for regenerative therapies.
References
Salingcarnboriboon, R., Yoshitake, H., Tsuji, K., et al. (2003). Establishment of tendon-derived cell lines exhibiting pluripotent mesenchymal stem-like property. Experimental Cell Research, 287, 289–300.
Scutt, N., Rolf, C. G., & Scutt, A. (2006). Glucocorticoids inhibit tenocyte proliferation and tendon progenitor cell recruitment. Journal of Orthopaedic Research, 24(2), 173–182.
Strassburg, S., Smith, R., Goodship, A., Hardingham, T., & Clegg, P. (2006). Adult and late fetal equine tendons contain cell populations with weak progenitor properties in comparison to bone marrow derived mesenchymal stem cells. Transection of Orthopaedic Research Society Annual Meeting, 31, 1113.
de Mos, M., Koevoet, W. J., Jahr, H., et al. (2007). Intrinsic differentiation potential of adolescent human tendon tissue: an in-vitro cell differentiation study. BMC Musculoskeletal Disorder, 8, 16.
Hashimoto, Y., Yoshida, G., Toyoda, H., & Takaoka, K. (2007). Generation of tendon-to-bone interface ‘enthesis’ with the use of recombinant BMP-2 in a rabbit model. Journal of Orthopaedic Research, 25, 1415–1424.
Bi, Y., Ehirchiou, D., Kilts, T. M., et al. (2007). Identification of tendon stem / progenitor cells and the role of the extracellular matrix in their niche. Nature Medicine, 13(10), 1219–1227.
Zhang, J., & Wang, J. H. C. (2010). Characterization of differential properties of rabbit tendon stem cells and tenocytes. BMC Musculoskeletal Disorder, 11, 10.
Rui, Y. F., Lui, P. P. Y., Li, G., Fu, S. C., Lee, Y. W., & Chan, K. M. (2010). Isolation and characterization of multi-potent rat tendon-derived stem cells. Tissue Engineering Part A, 16(5), 1549–1558.
Yin, Z., Chen, X., Chen, J. L., et al. (2010). The regulation of tendon stem cell differentiation by the alignment of nanofibers. Biomaterials, 31(8), 2163–2175.
Zhang, J., Pan, T., Liu, Y., & Wang, J. H. (2010). Mouse treadmill running enhances tendons by expanding the pool of tendon stem cells (TSCs) and TSC-related cellular production of collagen. Journal of Orthopaedic Research, 28(9), 1178–1183.
Zhang, J., & Wang, J. H. (2010). Platelet-rich plasma releasate promotes differentiation of tendon stem cells into active tenocytes. American Journal of Sports Medicine, 38(12), 2477–2486.
Colter, D. C., Clas, R., DiGirolamo, C. M., & Prockop, D. J. (2000). Rapid expansion of recycling stem cells in cultures of plastic-adherent cells from human bone marrow. Proceedings of the National Academy of Sciences of the United States of America, 97(7), 3213–3218.
Dezawa, M., Ishikawa, H., Itokazu, Y., et al. (2005). Bone marrow stromal cells generate muscle cells and repair muscle degeneration. Science, 309(5732), 314–317.
Rui, Y. F., Lui, P. P. Y., Ni, M., Chan, L. S., Lee, Y. W., & Chan, K. M. (2011). Mechanical loading increased BMP-2 expression which promoted osteogenic differentiation of tendon-derived stem cells. Journal of Orthopaedic Research, 29(3), 390–396.
Rui, Y. F., Lui, P. P. Y., Ni, M., et al. (2010). Higher expression of BMP receptors and BMP-2-induced osteogenesis in tendon-derived stem cells compared to bone marrow-derived stem cells. 7th Combined Meeting of the Orthopaedic Research Societies. 16th–20th Oct, 2010, Kyoto International Conference Centre, Kyoto, Japan.
Zhou, Z., Akinbiyi, T., Xu, L., et al. (2010). Tendon-derived stem / progenitor cell aging: defective self-renewal and altered fate. Aging Cell, 9(5), 911–915.
Stenderup, K., Justesen, J., Eriksen, E. F., Rattan, S. I., & Kassem, M. (2001). Number and proliferative capacity of osteogenic stem cells are maintained during aging and in patients with osteoporosis. Journal of Bone and Mineral Research, 16, 1120–1129.
Pittenger, M. F., Mackay, A. M., Beck, S. C., et al. (1999). Multilineage potential of adult human mesenchymal stem cells. Science, 284, 143–047.
Halfon, S., Abramov, N., Grinblat, B., & Ginis, I. (2011). Markers distinguishing mesenchymal stem cells from fibroblasts are downregulated with passaging. Stem Cells and Development, 20(1), 53–66.
Rombouts, W. J., & Ploemacher, R. E. (2003). Primary murine MSC show highly efficient homing to the bone marrow but lose homing ability following culture. Leukemia, 17(1), 160–170.
Azizi, S. A., Stokes, D., Augelli, B. J., DiGirolamo, C., & Prockop, D. J. (1998). Engraftment and migration of human bone marrow stromal cells implanted in the brains of albino rats—similarities to astrocyte grafts. Proceedings of the National Academy of Sciences of the United States of America, 95, 3908–3913.
Kopen, G. C., Prockop, D. J., & Phinney, D. G. (1999). Marrow stromal cells migrate throughout forebrain and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse brains. Proceedings of the National Academy of Sciences of the United States of America, 96, 10711–10716.
Fukuda, K. (2002). Molecular characterization of regenerated cardiomyocytes derived from adult mesenchymal stem cells. Congenital Anomalies (Kyoto), 42, 1–9.
Makino, S., Fukuda, K., Miyoshi, S., et al. (1999). Cardiomyocytes can be generated from marrow stromal cells in vitro. Journal of Clinical Investigation, 103, 697–705.
Kuo, T. K., Ho, J. H., & Lee, O. K. (2009). Mesenchymal stem cell therapy for nonmusculoskeletal diseases: emerging applications. Cell Transplantation, 18, 1013–1028.
Wang, J. S., Shum-Tim, D., Galipeau, J., Chedrawy, E., Eliopoulos, N., & Chiu, R. C. (2000). Marrow stromal cells for cellular cardiomyoplasty: feasibility and potential clinical advantages. The Journal of Thoracic and Cardiovascular Surgery, 120, 999–1005.
Lee, K. D., Kuo, T. K., Whang-Peng, J., et al. (2004). In vitro hepatic differentiation of human mesenchymal stem cells. Hepatology, 40, 1275–1284.
Izadpanah, R., Kaushal, D., Kriedt, C., et al. (2008). Long-term in vitro expansion alters the biology of adult mesenchymal stem cells. Cancer Research, 68(11), 4229–4238.
Izadpanah, R., Trygg, C., Patel, B., et al. (2006). Biologic properties of mesenchymal stem cells derived from bone marrow and adipose tissue. Journal of Cellular Biochemistry, 99, 1285–1297.
Jiang, Y., Jahagirdar, B. N., Reinhardt, R. L., et al. (2002). Pluripotency of mesenchymal stem cells derived from adult marrow. Nature, 418(6893), 41–49.
Bonab, M. M., Alimoghaddam, K., Talebian, F., Ghaffari, S. H., Ghavamzadeh, A., & Nikbin, B. (2006). Aging of mesenchymal stem cell in vitro. BMC Cell Biology, 7, 14.
Muraglia, A., Cancedda, R., & Quarto, R. (2000). Clonal mesenchymal progenitors from human bone marrow differentiate in vitro according to a hierarchical model. Journal of Cell Science, 113(Pt. 7), 1161–1166.
Banfi, A., Muraglia, A., Dozin, G., Mastrogiacomo, M., Cancedda, R., & Quarto, R. (2000). Proliferation kinetics and differentiation potential of ex vivo expanded human bone marrow stromal cells: implications for their use in cell therapy. Experimental Hematology, 28, 707–715.
Bruder, S. P., Jaiswal, N., & Haynesworth, S. E. (1997). Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. Journal of Cellular Biochemistry, 64, 278–294.
Baxter, M. A., Wynn, R. F., Jowitt, S. N., Wraith, J. E., Fairbairn, L. J., & Bellantuono, I. (2004). Study of telomere length reveals rapid aging of human marrow stromal cells following in vitro expansion. Stem Cells, 22, 675–682.
Tan, Q., Rui, Y. F., Lee, Y. W., & Lui, P. P. Y. (2011). Effect of in vitro passages on the biological properties of tendon-derived stem cells (TDSCs)—Implication in musculoskeletal tissue engineering. In Proceedings of International Symposium of Ligament and Tendon (ISL&T) XI, 12th January, 2011, Long Beach, California, USA.
Levesque, J. P., Helwani, F. M., & Winkler, I. G. (2010). The endosteal ‘osteoblastic’ niche and its role in hematopoietic stem cell homing and mobilization. Leukemia, 24(12), 1979–1992.
Zhang, J., & Wang, J. H. C. (2010). Mechanical response of tendon stem cells: implications of tendon homeostasis and pathogenesis of tendinopathy. Journal of Orthopaedic Research, 28(5), 639–643.
Wang, J. C., & Thampatty, B. P. (2008). Mechanobiology of adult of stem cells. International Review of Cell and Molecular Biology, 271, 297–342.
Chuen, F. S., Chuk, C. Y., Ping, W. Y., Nar, W. W., Kim, H. L., & Ming, C. K. (2004). Immunohistochemical characterization of cells in adult human patella tendons. Journal of Histochemisty & Cytochemistry, 52(9), 1151–1157.
Yoshizawa, T., Takizawa, F., Iizawa, F., et al. (2004). Homeobox protein MSX2 acts as a molecular defense mechanism for preventing ossification in ligament fibroblasts. Molecular and Cellular Biology, 24, 3460–3472.
Hoffmann, A., Pelled, G., Turgeman, G., et al. (2006). Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells. Journal of Clinical Investigation, 116, 940–952.
McNeilly, C. M., Banes, A. J., Benjamin, M., & Ralphs, J. R. (1996). Tendon cells in vivo form a three dimensional network of cell processes linked by gap junctions. Journal of Anatomy, 189(Pt 3), 593–600.
Luo, Q., Song, G., Song, Y., Xu, B., Qin, J., & Shi, Y. (2009). Indirect co-culture with tenocytes promotes proliferation and mRNA expression of tendon/ligament related genes in rat bone marrow mesenchymal stem cells. Cytotechnology, 61(1–2), 1–10.
Crisan, M., Yap, S., Casteilla, L., et al. (2008). A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell, 3(11), 301–313.
Caplan, A. I. (2008). All MSCs are pericytes? Cell Stem Cell, 3(3), 229–230.
da Silva Meirelles, L., Chagastelles, P. C., & Nardi, N. B. (2006). Mesenchymal stem cells reside in virtually all post-natal organs and tissues. Journal of Cell Science, 119(Pt 11), 2204–2213.
Tempfer, H., Wagner, A., Gehwolf, R., et al. (2009). Perivascular cells of the supraspinatus tendon express both tendon- and stem cell-related markers. Histochemistry and Cell Biology, 131, 733–741.
Helm, G. A., Li, J. Z., Alden, T. D., et al. (2001). A light and electron microscopic study of ectopic tendon and ligament formation induced by bone morphogenetic protein-13 adenoviral gene therapy. Journal of Neurosurgery, 95, 298–307.
Wong, Y. P., Fu, S. C., Cheuk, Y. C., Lee, K. M., Wong, M. W., & Chan, K. M. (2005). Bone morphogenetic protein 13 stimulates cell proliferation and production of collagen in human patellar tendon fibroblasts. Acta Orthopaedica, 76(3), 421–427.
Theobald, P., Benjamin, M., Nokes, L., & Pugh, N. (2005). Review of the vascularisation of the human Achilles tendon. Injury, 36(11), 1267–1272.
Arnoczky, S. P., Lavagnino, M., & Egerbacher, M. T. (2007). The response of tendon cells to changing loads: Implications in the etiopathogenesis of tendinopathy. In S. L. Y. Woo, P. A. F. H. Renstrom, & S. P. Arnoczky (Eds.), Tendinopathy in athletes (pp. 46–59). USA: Blackwell.
Riley, G. P., Harrall, R. L., Constant, C. R., Cawston, T. E., & Hazlman, B. L. (1996). Prevalence and possible pathological significance of calcium phosphate salt accumulation in tendon matrix degeneration. Annals of the Rheummatic Diseases, 55, 109–115.
Fu, S. C., Chan, B. P., Wang, W., Pau, H. M., Chan, K. M., & Rolf, C. G. (2002). Increase expression of matrix metalloproteinase 1(MMP1) in 11 patients with patellar tendinosis. Acta Orthopaedica Scandinavica, 73(6), 658–662.
Fu, S. C., Wang, W., Pau, H. M., Wong, Y. P., Chan, K. M., & Rolf, C. G. (2002). Increased expression of transforming growth factor-beta1 in patellar tendinosis. Clinical Orthopaedics and Related Research, 400, 174–183.
Jones, G. C., Corps, A. N., Pennington, C. J., et al. (2006). Expression profiling of metalloproteinases and tissue inhibitors of metalloproteinases in normal and degenerate human Achilles tendon. Arthritis & Rheumatism, 54(3), 832–842.
Maffulli, N., Reaper, J., Ewen, S. W. B., Waterston, S. W., & Barrass, V. (2006). Chondral metaplasia in calcific tendinopathy of the Achilles tendon. Clinical Journal of Sport Medicine, 16, 329–334.
Karousou, E., Ronga, M., Vigetti, D., Passi, A., & Maffulli, N. (2008). Collagens, proteoglycans, MMP-2, MMP-9 and TIMPs in human Achilles tendon rupture. Clinical Orthopaedics and Related Research, 466, 1577–1582.
Karousou, E., Ronga, M., Vigetti, D., Passi, A., & Maffulli, N. (2010). Molecular interactions in extracellular matrix of tendon. Frontiers in Bioscience (Elite Edition), 2, 1–12.
Fu, S. C., Chan, K. M., & Rolf, C. G. (2007). Increased deposition of sulfated glycosaminoglycans in human patellar tendinopathy. Clinical Journal of Sport Medicine, 17(2), 129–134.
Fenwick, S., Harrall, R., Hackney, R., et al. (2002). Endochondral ossification in Achilles and patella tendinopathy. Rheumatology (Oxford), 41, 474–476.
Rui, Y. F., Lui, P. P. Y., Chan, L. S., Chan, K. M., Fu, S. C., & Li, G. (2011). Does erroneous differentiation of tendon-derived stem cells contribute to the pathogenesis of calcifying tendinopathy? Chinese Medical Journal (English), 124(4), 606–610.
Lui, P. P. Y., Fu, S. C., Chan, L. S., Hung, L. K., & Chan, K. M. (2009). Chondrocyte phenotype and ectopic ossification in collagenase-induced tendon degeneration. Journal of Histochemistry & Cytochemistry, 57(2), 91–100.
Rui, Y. F., Lui, P. P. Y., Wong, Y. M., et al. (2011). Impaired differentiation and proliferation of tendon-derived stem cells (TDSCs) isolated from collagenase-induced tendon injury model—A potential mechanism for the chondro-ossification and failed healing tendinopathy. In Proceedings of 57th Annual Meeting of the Orthopaedic Research Society, 13th–16th January, 2011, Long Beach, California, USA.
Archambault, J. M., Jelinsky, S. A., Lake, S. P., Hill, A. A., Glaser, D. L., & Soslowsky, L. J. (2007). Rat supraspinatus tendon expresses cartilage markers with overuse. Journal of Orthopaedic Research, 25, 617–624.
Clegg, P. D., Strassburg, S., & Smith, R. K. (2007). Cell phenotypic variation in normal and damaged tendons. International Journal of Experimental Pathology, 88, 227–235.
Speer, M. Y., Yang, H. Y., Brabb, T., et al. (2009). Smooth muscle cells give rise to osteochondrogenic precursors and chondrocytes in calcifying arteries. Circulation Research, 104(6), 733–741.
Kim, S. Y., Choi, H. Y., Myung, K. B., & Choi, Y. W. (2008). The expression of molecular mediators in the idiopathic cutaneous calcification and ossification. Journal of Cutaneous Pathology, 35(9), 826–831.
Lounev, V. Y., Ramachandran, R., Wosczyna, M. N., et al. (2009). Identification of progenitor cells that contribute to heterotopic skeletogenesis. The Journal of Bone and Joint Surgery American, 91(3), 652–663.
Bajada, S., Marshall, M. J., Wright, K. T., Richardson, J. B., & Johnson, W. E. (2009). Decreased osteogenesis, increased cell senescence and elevated Dickkopf-1 secretion in human fracture nonunion stromal cells. Bone, 45(4), 726–735.
Langberg, H., Skovgaard, D., Karamouzis, M., Bulow, J., & Kjaer, M. (1999). Metabolism and inflammatory mediators in the peritendinous space measured by microdialysis during intermittent isometric exercise in humans. The Journal of Physiology, 515(Pt 3), 919–927.
Zhang, J., & Wang, J. H. (2010). Production of PGE(2) increases in tendons subjected to repetitive mechanical loading and induces differentiation of tendon stem cells into non-tenocytes. Journal of Orthopaedic Research, 28(2), 198–203.
Kuo, C. K., & Tuan, R. S. (2008). Mechanoactive tenogenic differentiation of human mesenchymal stem cells. Tissue Engineering Part A, 14(10), 1615–1627.
Almekinders, L. C., Banes, A. J., & Ballenger, C. A. (1993). Effects of repetitive motion on human fibroblasts. Medicine & Science in Sports & Exercise, 25, 603–607.
Wang, J. H., Jia, F., Yang, G., et al. (2003). Cyclic mechanical stretching of human tendon fibroblasts increases the production of prostaglandin E(2) and levels of cyclooxygenase expression: a novel in vitro model study. Connective Tissue Research, 44, 128–133.
Rodeo, S. A., Suzuki, K., Deng, X. H., Wozney, J., & Warren, R. F. (1999). Use of recombinant human bone morphogenetic protein-2 to enhance tendon healing in a bone tunnel. American Journal of Sports Medicine, 27, 476–488.
Ma, C. B., Kawamura, S., Deng, X. H., et al. (2007). Bone morphogenetic proteins-signaling plays a role in tendon-to-bone healing: a study of rhBMP-2 and noggin. American Journal of Sports Medicine, 35, 597–604.
Tang, Y., Tang, W., Lin, Y., et al. (2008). Combination of bone tissue engineering and BMP-2 gene transfection promotes bone healing in osteoporotic rats. Cell Biology International, 32, 1150–1157.
Martinek, V., Latterman, C., Usas, A., et al. (2002). Enhancement of tendon-bone integration of anterior cruciate ligament grafts with bone morphogenetic protein-2 gene transfer: a histological and biomechanical study. Journal of Bone & Joint Surgery American, 84, 1123–1131.
Chen, C. H., Liu, H. W., Tsai, C. L., Yu, C. M., Lin, I. H., & Hsiue, G. H. (2008). Photoencapsulation of bone morphogenetic protein-2 and periosteal progenitor cells improve tendon graft healing in a bone tunnel. American Journal of Sports Medicine, 36, 461–473.
Jiang, Y., Chen, L. K., Zhu, D. C., et al. (2010). The inductive effect of bone morphogenetic protein-4 on chondral-lineage differentiation and in situ cartilage repair. Tissue Engineering Part A, 16(5), 1621–1632.
Neuwirth, J., Fuhrmann, R. A. E., Veit, A., et al. (2006). Expression of bioactive bone morphogenetic proteins in the subacromial bursa of patients with chronic degeneration of the rotator cuff. Arthritis Research & Therapy, 8, R92.
Sato, R., Uchida, K., Kobayashi, S., et al. (2007). Ossification of the posterior longitudinal ligament of the cervical spine: histopathological findings around the calcification and ossification front. Journal of Neurosurgery: Spine, 7, 174–183.
Li, H., Jiang, L. S., & Dai, L. Y. (2007). Hormones and growth factors in the pathogenesis of spinal ligament ossification. European Spine Journal, 16, 1075–1084.
Tanno, M., Furukawa, K. I., Ueyama, K., Harata, S., & Motomura, S. (2003). Uniaxial cyclic stretch induces osteogenic differentiation and synthesis of bone morphogenetic proteins of spinal ligament cells derived from patients with ossification of the posterior longitudinal ligaments. Bone, 33, 475–484.
Lui, P. P. Y., Chan, L. S., Cheuk, Y. C., Lee, Y. W., & Chan, K. M. (2009). Expression of bone morphogenetic protein-2 in the chondrogenic and ossifying sites of calcific tendinopathy and traumatic tendon injury rat models. Journal of Orthopaedic Surgery and Research, 21(4), 27.
Yee Lui, P. P., Wong, Y. M., Rui, Y. F., Lee, Y. W., Chan, L. S., & Chan, K. M. (2011). Expression of osteogenic BMPs in calcified failed tendon healing model of tendinopathy. Journal of Orthopaedic Research, 29(6): 816–821.
Rui, Y. F., Lui, P. P. Y., Rolf, C. G., Wong, Y. M., Lee, Y. W., & Chan, K M. (2011). Expression of chondro-osteogenic BMPs in clinical samples of calcifying and uncalcifying patellar tendinopathy—A histopathological study. In Proceedings of International Symposium of Ligament and Tendon (ISL&T) XI, 12th January, 2011, Long Beach, California, USA.
Csiszar, A., Smith, K. E., Koller, A., Kaley, G., Edwards, J. G., & Ungvari, Z. (2005). Regulation of bone morphogenetic protein-2 expression in endothelial cells: role of nuclear factor-kappaB activation by tumor necrosis factor-alpha, H2O2, and high intra-vascular pressure. Circulation, 111, 2364–2372.
Csiszar, A., Labinskyy, N., Smith, K. E., et al. (2007). Downregulation of bone morphogenetic protein 4 expression in coronary arterial endothelial cells-Role of shear stress and the cAMP/protein kinase A pathway. Arteriosclerosis, Thrombosis, and Vascular Biology, 27, 776–782.
Rath, B., Nam, J., Knobloch, T. J., Lannutti, J. J., & Agarwal, S. (2008). Compressive forces induce osteogenic gene expression in calvarial osteoblasts. Journal of Biomechanics, 41(5), 1095–1103.
Lui, P. P. Y., Chan, L. S., Lee, Y. W., Fu, S. C., & Chan, K. M. (2010). Sustained expression of proteoglycans and collagen type III / Type I ratio in a calcified tendinopathy model. Rheumatology, 49(2), 231–239.
Seib, F. P., Franke, M., Jing, D., Werner, C., & Bornhauser, M. (2009). Endogenous bone morphogenetic proteins in human bone marrow-derived multipotent mesenchymal stromal cells. European Journal of Cell Biology, 88, 257–271.
Lui, P. P. Y., Chan, L. S., Fu, S. C., & Chan, K. M. (2010). Expression of sensory neuropeptides in tendon is associated with failed healing and activity-related tendon pain in collagenase-induced tendon injury. American Journal of Sports Medicine, 38(4), 757–764.
Shih, C., & Bernard, G. W. (1997). Neurogenic substance P stimulates osteogenesis in vitro. Peptides, 18, 323–326.
Xu, Y., & Murrell, G. A. (2008). The basic science of tendinopathy. Clinical Orthopaedics and Related Research, 466, 1528–1538.
Tsuzaki, M., Bynum, D., Almekinders, L., Yang, X., Faber, J., & Banes, A. J. (2003). ATP modulates load-inducible IL-1beta, COX2, and MMP-3 gene expression in human tendon cells. Journal of Cellular Biochemistry, 89(3), 556–562.
Wang, J. H., Jia, F., Yang, G., et al. (2003). Cyclic mechanical stretching of human tendon fibroblasts increases the production of prostaglandin E2 and levels of cyclooxygenase expression: a novel in vitro model study. Connective Tissue Research, 44(3–4), 128–133.
Riley, G. (2004). The pathogenesis of tendinopathy. A molecular perspective. Rheumatology (Oxford), 43(2), 131–142.
Gotoh, M., Hamada, K., Yamakawa, H., Inoue, A., & Fukuda, H. (1998). Increased substance P in subacromial bursa and shoulder pain in rotator cuff diseases. Journal of Orthopaedic Research, 16(8), 618–621.
Lian, O., Dahl, J., Ackermann, P. W., Frihagen, F., Engebretsen, I., & Bahr, R. (2006). Pronociceptive and antinociceptive neuromediators in patellar tendinopathy. American Journal of Sports Medicine, 34, 1801–1808.
Schubert, T. E., Weidler, C., Lerch, K., Hofstadter, F., & Straub, R. H. (2005). Achilles tendinosis is associated with sprouting of substance P positive nerve fibers. Annals of the Rheumatic Diseases, 64, 1083–1086.
Lerner, U. H. (2002). Neuropeptidergic regulation of bone resorption and bone formation. Journal of Musculoskeletal & Neuronal Interactions, 2, 440–447.
Maffulli, N., Wong, J., & Almekinders, L. C. (2003). Types and epidemiology of tendinopathy. Clinics in Sports Medicine, 22(4), 675–692.
Butler, D. L., Juncosa, N., & Dressler, M. R. (2004). Functional efficacy of tendon repair processes. Annual Review of Biomedical Engineering, 6, 303–329.
Proctor, C. S., Jackson, D. W., & Simon, T. M. (1997). Characterization of the repair tissue after removal of the central one-third of the patellar ligament: an experimental study in a goat model. Journal of Bone and Joint Surgery American Volume, 79(7), 997–1006.
Hampson, K., Forsyth, N. R., El Haj, A., & Maffulli, N. (2008). Tendon tissue engineering. In N. Ashammakhi, R. Reis & F. Chiellini (Ed.), E-book: Topics in tissue engineering Vol 4 (chapter 3). http://www.oulu.fi/spareparts/ebook_topics_in_t_e_vol4/index.html
Awad, H. A., Butler, D. L., Boivin, G. P., et al. (1999). Autologous mesenchymal stem cell-mediated repair of tendon. Tissue Engineering, 5(3), 267–277.
Awad, H. A., Butler, D. L., Dressler, M. R., Smith, F., Boivin, G. P., & Young, R. G. (2003). Repair of patellar tendon injuries using mesenchymal stem cells and collagen scaffolds. Journal of Orthopaedic Research, 21(3), 420–431.
Young, R. G., Butler, D. L., Weber, W., Caplan, A. I., Gordon, S. L., & Fink, D. J. (1998). Use of mesenchymal stem cells in a collagen matrix for Achilles tendon repair. Journal of Orthopaedic Research, 16(4), 406–413.
Ouyang, H. W., Goh, J. C., Thambyah, A., Teoh, S. H., & Le, E. H. (2003). Knitted poly-lactide-co-glycolide scaffold loaded with bone marrow stromal cells in repair and regeneration of rabbit Achilles tendon. Tissue Engineering, 9(3), 431–439.
Kryger, G. S., Chong, A. K., Costa, M., Pham, H., Bates, S. J., & Chang, J. A. (2007). Comparison of tenocytes and mesenchymal stem cells for use in flexor tendon tissue engineering. Journal of Hand Surgery American Volume, 32(5), 597–605.
Juncosa-Melvin, N., Boivin, G. P., Galloway, M. T., Gooch, C., West, J. R., Sklenka, A. M., et al. (2005). Effects of cell-to-collagen ratio in mesenchymal stem cell-seeded implants on tendon repair biomechanics and histology. Tissue Engineering, 11(3–4), 448–457.
Juncosa-Melvin, N., Boivin, G. P., Gooch, C., et al. (2006). The effect of autologous mesenchymal stem cells on the biomechanics and histology of gel-collagen sponge constructs used for rabbit patellar tendon repair. Tissue Engineering, 12(2), 369–379.
Chong, A. K., Ang, A. D., Goh, J. C., Hui, J. H., Lim, A. Y., Lee, E. H., et al. (2007). Bone marrow-derived mesenchymal stem cells influence early tendon-healing in a rabbit Achilles tendon model. Journal of Bone and Joint Surgery American Volume, 89(1), 74–81.
Hankemeier, S., van Griensven, M., Ezechieli, M., et al. (2007). Tissue engineering of tendons and ligaments by human bone marrow stromal cells in a liquid fibrin matrix in immunodeficient rats: results of a histologic study. Archives of Orthopaedic and Trauma Surgery, 127(9), 815–821.
Chen, X., Song, X., Yin, Z., et al. (2009). Stepwise differentiation of human embryonic stem cells promotes tendon regeneration by secreting fetal tendon matrix and differentiation factors. Stem Cells, 27(6), 1276–1287.
Harris, M. T., Butler, D. L., Boivin, G. P., Florer, J. B., Schantz, E. J., & Wenstrup, R. J. (2004). Mesenchymal stem cells used for rabbit tendon repair can form ectopic bone and express alkaline phosphatase activity in constructs. Journal of Orthopaedic Research, 22(5), 998–1003.
Tasso, R., Augell, A., Carida, M., et al. (2009). Development of sacromas in mice implanted with mesenchymal stem cells seeded onto bioscaffolds. Carcinogenesis, 30(1), 150–157.
Sakaguchi, Y., Sekiya, I., Yagishita, K., & Muneta, T. (2005). Comparison of human stem cells derived from various mesenchymal tissues: superiority of synovium as a cell source. Arthritis & Rheumatism, 52(8), 2521–2529.
Deans, T. L., & Elisseeff, J. H. (2009). Stem cells in musculoskeletal engineered tissue. Current Opinion in Biotechnology, 20, 537–544.
Baksh, D., Yao, R., & Tuan, R. S. (2007). Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells, 25, 1384–1392.
Hayashi, O., Katsube, Y., Hirose, M., Ohgushi, H., & Ito, H. (2008). Comparison of osteogenic ability of rat mesenchymal stem cells from bone marrow, periosteum, and adipose tissue. Calcified Tissue International, 82, 238–247.
Lin, L., Shen, Q., Wei, X., et al. (2009). Comparison of osteogenic potentials of BMP4 transduced stem cells from autologous bone marrow and fat tissue in a rabbit model of calvarial defects. Calcified Tissue International, 85, 55–65.
Singh, S., Dhaliwal, N., Crawford, R., & Xiao, Y. (2009). Cellular senescence and longevity of osteophyte-derived mesenchymal stem cells compared to patient-matched bone marrow stromal cells. Journal of Cellular Biochemistry, 108, 839–850.
Wang, L., Tran, I., Seshareddy, K., Weiss, M. L., & Detamore, M. S. (2009). A comparison of human bone marrow-derived mesenchymal stem cells and human umbilical cord-derived mesenchymal stromal cells for cartilage tissue engineering. Tissue Engineering Part A, 15(8), 2259–2266.
Kolambkar, Y. M., Peister, A., Soker, S., Atala, A., & Guldberg, R. E. (2007). Chondrogenic differentiation of amniotic fluid-derived stem cells. Journal of Molecular Histology, 38, 405–413.
Kern, S., Eichler, H., Stoeve, J., Kluter, H., & Bieback, K. (2006). Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells, 24, 1294–1301.
Cheng, M. T., Liu, C. L., Chen, T. H., & Lee, O. K. (2010). Comparison of potentials between stem cells isolated from human anterior cruciate ligament and bone marrow for ligament tissue engineering. Tissue Engineering Part A, 16(7), 2237–2235.
Yoshimura, H., Muneta, T., Nimura, A., Yokoyama, A., & Koga, H. (2007). Comparison of rat mesenchymal stem cells derived from bone marrow, synovium periosteum, adipose tissue, and muscle. Cell and Tissue Research, 327, 449–462.
Matsubara, T., Tsutsumi, S., Pan, H., et al. (2004). A new technique to expand human mesenchymal stem cells using basement membrane extracellular matrix. Biochemical and Biophysical Research Communications, 313, 503–508.
Lui, P. P. Y., Rui, Y. F., Ni, M., et al. (2011). Comparison of potentials of stem cells isolated from tendon and bone marrow for musculoskeletal tissue engineering. In Proceedings of 57th Annual Meeting of the Orthopaedic Research Society, 13th–16th January, 2011, Long Beach, California, USA.
Ni, M., Lui, P. P. Y., Rui, Y. F., et al. (2011). Allogenic Tendon-derived stem cells (TDSCs) promoted tendon regeneration. In Proceedings of 57th Annual Meeting of the Orthopaedic Research Society, 13th–16th January, 2011, Long Beach, California, USA.
Ni, M., Lui, P. P. Y., Rui, Y. F., et al. (2011). Tendon-derived stem cell (TDSC): A better stem cell source compared to bone marrow-derived mesenchymal stem cell (BMSC) for tendon regeneration? In Proceedings of International Symposium of Ligament and Tendon (ISL&T) XI, 12th January, 2011, Long Beach, California, USA.
Acknowledgement
This work was supported by resources donated by the Hong Kong Jockey Club Charities Trust.
Disclosures
The authors indicate no potential conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lui, P.P.Y., Chan, K.M. Tendon-Derived Stem Cells (TDSCs): From Basic Science to Potential Roles in Tendon Pathology and Tissue Engineering Applications. Stem Cell Rev and Rep 7, 883–897 (2011). https://doi.org/10.1007/s12015-011-9276-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12015-011-9276-0