
Abstract
The phytotoxic effects of lead (Pb) on seed germinability, seedling growth, photosynthetic performance, and nutrient accumulation (K+ and Cu2+) in two maize genotypes (EV-1098 and EV-77) treated with varying levels of PbSO4 (0.01, 0.1, and 1.0 mg L−1) were appraised in this study. In the seed germination experiment, lead stress significantly reduced seed germination percentage and index, plumule and radicle lengths as well as fresh and dry weights in both genotypes. In the second experiment, lengths and fresh and dry weights of shoots and roots decreased due to Pb in both genotypes with increase in plant age. Higher Pb levels also decreased photosynthetic rate (A), water use efficiency (A/E), and intrinsic water use efficiency (A/g s), but increased transpiration rate (E) and C i/C a ratio as a result of increase in stomatal conductance (g s). The concentrations of K+ and Cu2+ decreased in root, stem, and leaves of both genotypes, which could be a direct consequence of multifold increase in Pb concentration in these tissues. Overall, cv. EV-1098 had better Pb tolerance potential than EV-77 because the former genotype showed less reduction in seed germinability parameters, photosynthetic performance, and K+ and Cu2+ accumulation in shoot and root under lead stress.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil contamination by heavy metals is a major ecological concern due to its widespread release from agriculture, industry, and human activities [1]. Soils of many areas have been contaminated with heavy metal toxicity that poisons the plant–soil system, degrade the soil, and reduce the quality and yield of most crops [2–5]. Moreover, it also poses a serious threat to the health of animals and humans upon entering the food chain.
Among different potentially toxic metals, lead (Pb) is one of the most common pollutants in the environment that readily accumulates in soils and sediments [6]. Main sources of Pb release are mining and smelting of Pb ores, chimneys of factories using Pb, metal plating and finishing operations, effluents from the storage battery, industry, pesticides, fertilizers, and additives in pigments and gasoline [7]. Soil contamination with Pb has gained considerable attention in the recent era and it seems to be not mitigated in the near future [8].
Some metals, such as Mn, Cu, and Zn, are essential micronutrients for microorganisms, plants, and animals in low concentrations, but at high concentrations, they have strong toxic effects particularly on plants and animals [9–11]. However, as compared to these metals, Pb is typically nonessential metal because its biological function has not been reported in the literature [12, 13]. In plants, its uptake, transport, and accumulation have been reported to be mainly dependent on soil type and nature of a plant species. This metal is absorbed and accumulated in different plant tissues [14], generally with the highest amount in the root tissues [15–17].
Plants grown in Pb-contaminated soils show visible nonspecific symptoms including stunted growth, chlorosis, and inhibition of root growth [18–21]. When Pb enters the cells, it produces a wide range of toxic effects on physiological and biochemical processes of plants. It causes inhibition of enzyme activities, perturbation in mineral nutrition and water balance, change in hormonal status [22], and alterations in lipid composition and protein content in tissues [23, 24]. These disorders upset normal physiological activities causing reduction in growth and metabolism that leads to membrane disorganization and reduced photosynthesis [22, 25].
Heavy metals have been reported to have inhibitory effects on seed germination and seedling growth in a number of earlier studies [26–29]. As germination and early seedling growth are critical stages in the development of plants [30], so they play an important role in getting food crop for subsequent growth. Therefore, keeping in view of all these deleterious aspects of Pb toxicity, the present study was conducted to examine the effects of Pb stress on seed germinability, seedling growth, photosynthesis as well as nutrient concentration (K+ and Cu2+) in two most widely cultivated maize genotypes (EV-1098 and EV-77).
Materials and Methods
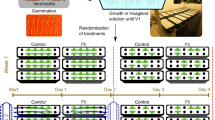
The seeds of two most widely cultivated maize genotypes (EV-77 and EV-1098) were obtained from the Maize and Millet Research Institute, Sahiwal, Pakistan and evaluated for their Pb tolerance potential in two different experiments. In seed germination experiment, ten seeds were sown in Petri plates lined with double filter paper and surface sterilized with 0.1% HgCl2 for 2 min. Varying levels of lead (control, 0.01, 0.1, and 1.0 mg L−1 of PbSO4) were prepared in Hoagland’s nutrient solution [31], and 10 ml of each solution was applied to each Petri plate. The Petri plates were placed under continuous white fluorescent light (PAR 350 μmol m−2 s−1) in a growth room at 26 ± 3°C. Seeds were allowed to germinate for 7 days, and seeds germinated were counted daily to calculate seed germination percentage and index. A seed was counted germinated when both plumule and radicle had emerged ≥0.5 cm. After 7 days of sowing, seedlings were harvested and rinsed with distilled water. Their plumules and radicles were separated and fresh weights determined. They were then wrapped in paper bags and dried in an oven at 65°C up to constant dry weights and their dry weights recorded.
In the second experiment, seeds of both genotypes were sterilized with 0.1% HgCl2 for 2 min and sown in plastic pots containing 10 kg well-washed river sand. Varying levels of lead (control, 0.01, 0.1, and 1.0 mg L−1 of PbSO4) were prepared in Hoagland’s nutrient solution as mentioned above, and 2.0 L of each solution was applied to each pot. The solutions were applied weekly to maintain constant levels of Pb by completely replacing old solution with appropriate fresh one. The pots were placed under natural conditions with PPFD 1,450 μmol m−2 s−1 (14.2 h day−1), average day and night temperatures 21 ± 4°C and 16 ± 2°C, and relative humidity 40 ± 3.2% and 60 ± 4.5%, respectively. The experiment was laid down in a completely randomized design with factorial arrangement in three replicates. The data for growth parameters were recorded at 21 days after sowing. Plants were uprooted and their shoot and root lengths measured. Detached shoots and roots after properly washing them were placed in an oven and dried at 65°C up to constant dry weights and their dry weights recorded. Before harvesting the plants, the data for photosynthetic and gas exchange parameters including CO2 assimilation (A) and transpiration (E) rates, stomatal conductance (g s) were recorded using an open system LCA-4 ADC portable infrared gas analyzer (Analytical Development Company, Hoddesdon, England). The condition adjustments of the equipment used were: leaf surface areas 6.25 cm2, ambient CO2 concentration (Cref) 290.1 μmol mole−1, temperature of leaf chamber (Tch) varied from 31°C to 33.8°C, leaf chamber gas flow rate (V) 394 mL min−1, leaf chamber gas flow rate (U) 256.66 mol m−2 s−1, ambient pressure (P) 98.9 kPa, water vapor pressure (eref) into the leaf chamber ranged from 4.4 to 6.6 mbar, and molar flow of air per unit leaf area (US) 410.6 mol m−2 s−1. The values of A and E were used to calculate water use efficiency (A/E), whereas the values of A and g s were used to work out intrinsic water use efficiency (A/g s). The values of intrinsic (C i) and ambient (C a) CO2 concentrations were used to calculate C i/C a ratio. For the determination of inorganic elements, the oven-dried well-ground material (0.1 g) of leaves and roots was digested in concentrated HNO3. The amount of dissolved cations like K+ was determined with a flame photometer (Jenway, PFP-70), whereas Pb2+ and Cu2+ were determined with an atomic absorption spectrophotometer (AAnalyst-300, Perkin Elmer, Germany). The data so collected were subjected to a two-way ANOVA, and mean values were compared with the least significance difference test at P ≤ 0.05 with a COSTAT computer package (CoHort Software, 2003, Monterey, CA) to determine statistical difference among them.
Results
During the germination experiment (after 7 days of metal treatment), lead (Pb) had a significantly adverse effect on seed germination and seedling growth of both maize genotypes (EV-1098 and EV-77). Both treatment (T) and genotype (G) terms differed significantly for percent seed germination, germination index, plumule and radicle length, and fresh and dry weights. Whereas the T × G interaction term was significant only for radicle length and plumule and radicle fresh and dry weights. Overall, seed germination percentage and index and radicle fresh and dry weights of both genotypes decreased consistently with increase in Pb levels. Similarly, plumule and radicle lengths also decreased with increasing Pb level of the growth medium, but the effect on radicle length in genotype EV-1098 was more inhibitory as compared to that on EV-77. The highest level of Pb (1.0 mg L−1) had the most adverse effects on these parameters as compared to the low Pb levels (Fig. 1).

Germination and seedling growth parameters of the two maize genotypes after 7 days of lead application [Bars represent SE values; *, **, *** = significant at 0.05, 0.01 and 0.001 levels, respectively; ns=non-significant. Means sharing the same lowercase letters are non-significant at 5% level, n = 10; T=treatments and G=genotype]
Similar to the inhibitory effects of Pb on seed germinability parameters, severe reduction in vegetative growth, photosynthesis, and K+ and Cu2+concentration was also observed in Pb-stressed maize plants after 21 days of growth in the second experiment. Statistical analysis of the data revealed that T, G, and T × G interaction terms differed significantly for shoot and root lengths and shoot and root fresh and dry weights recorded after 21 days of Pb application. Shoot length of both genotypes decreased consistently with increasing concentration of exogenously applied Pb, while root length and fresh and dry weights of shoots and roots of genotype EV-77 were not much affected at lower level of Pb stress (0.01 mg L−1). All these parameters decreased gradually with a gradual increase in Pb application. Genotype EV-1098 showed less reduction in growth attributes as compared to that in EV-77 (Fig. 2).

Germination and vegetative growth parameters of the two maize genotypes after 21 days of lead application [Bars represent SE values; *, **, *** = significant at 0.05, 0.01 and 0.001 levels, respectively; ns=non-significant. Means sharing the same lowercase letters are non-significant at 5% level, n = 3; T=treatments and G=genotype]
All gas exchange parameters showed a significant reduction after 21 days of Pb application. Photosynthetic rate (A), transpiration rate (E), stomatal conductance (g S), water use efficiency (A/E), and intrinsic water use efficiency (A/g s ) as well as leaf substomatal to ambient CO2 ratio (C i /C a) were significantly affected due to Pb stress. Overall, photosynthetic rate (A) of both genotypes decreased consistently with increase in Pb level. Lead stress caused a marked reduction in A/E and A/g s ratios of both genotypes with increasing concentration of Pb, while a significant increase in transpiration rate (E), stomatal conductance (g s), and C i /C a ratio was observed at the highest level of Pb (1.0 mg L−1; Fig. 3).

Photosynthetic and gas exchange parameters of the two maize genotypes after 21 days of lead application. [Bars represent SE values; *, **, *** = significant at 0.05, 0.01 and 0.001 levels, respectively; ns=non-significant. Means sharing the same lowercase letters are nonsignificant at 5% level, n = 3; T=treatments and G=genotype]
Potassium (K+) and copper (Cu2+) concentrations in root and leaves of both maize genotypes were also significantly affected by the addition of Pb to the rooting medium. Leaf Cu2+ content in leaves and roots of both genotypes decreased consistently with the increasing levels of Pb application. Although leaf K+ content also decreased significantly in both genotypes, the reduction was more in EV-1098 at higher levels of Pb stress (0.1 and 1.0 mg L−1) than that in EV-77. In contrast, root K+ content decreased markedly in EV-1098, but it was not much affected in EV-77 by Pb stress. The decrease in leaf and root K+ and Cu2+ was accompanied by a multifold increase in Pb2+ contents in the respective organs of both genotypes. Genotype EV-77 showed more increase in root Pb2+ contents as well as those of leaves. The reduction in all growth and photosynthetic attributes as well as that of K+ and Cu2+ accumulation parameters was negatively correlated with high accumulation of Pb in the roots and leaves of both maize genotypes (Fig. 4).

Potassium, copper, and lead concentrations in roots and leaves of the two maize genotypes after 21 days of lead application. [Bars represent SE values; *, **, *** = significant at 0.05, 0.01 and 0.001 levels, respectively; ns=non-significant. Means sharing the same lowercase letters are nonsignificant at 5% level, n = 3; T=treatments and G=genotype]
Discussion
The data obtained from this showed indicated that cv. EV-23 1098 exhibited better Pb tolerance potential than EV-77 because the former genotype showed less reduction in all parameters studied. Pb is considered to be typically nonessential nutrient for plants; however, plants easily take up Pb from the soil, accumulate in roots, and a small fraction is translocated upward to the shoots [13]. In this study, lead stress had a significant adverse effect on seed germination, seedling growth, and photosynthetic performance as well as on nutrient accumulation in two maize genotypes examined in this study. This is parallel to what has earlier been observed in a number of studies on different crops, e.g., wheat [32], rice [17], maize, and barley [33]. In the first experiment on germination, reduction in seed germination and early seedling attributes was suggested to be a direct consequence of excess Pb concentration in the growth medium of this crop. It has been shown that decreased seed germination may be a consequence of reduction in breakdown and mobilization of stored food in germinating seed as a result of decreased activity of starch- and protein-degrading enzymes, such as α-amylase and protease, caused by high Pb concentration [34]. This can directly result in reduced activity of meristematic cells and enzymes contained in the cotyledons and endosperm, thereby reducing seedling length and fresh and dry weights under Pb stress [35].
In the second experiment, all vegetative growth parameters decreased drastically under Pb stress applied for 21 days. However, the reduction in root length and fresh and dry weights was more prominent in roots as compared to shoot length under varying concentrations of Pb. Similar results have already been reported in wheat [35, 36], Albizia lebbeck [29], and Brassica juncea [37]. Roots are directly exposed to high concentration of Pb in growth medium, and Pb, being relatively less mobile, is sequestered in roots. Thus, roots have to face direct consequences of high Pb concentration. For example, it has been reported that high concentration of Pb can directly result in reduced mitosis in meristematic zone of Allium cepa roots, thereby arresting mitotic cell division [38] which results in reduced root proliferation [39, 40]. This can ultimately lead to reduced nutrient and water supply to aerial plant parts that reduces photosynthetic rates, alters water balance, and membrane structure and perturbs permeability functions [22], thereby causing reduced shoot growth under Pb stress.
Analogous to the inhibitory effects of Pb on vegetative growth paramaters, photosynthesis and concentrations of different elements were also affected in maize plants subjected to Pb stress for 21 days. The photosynthetic rate (A) was adversely affected by Pb toxicity which could be due to metal-induced reduction in the concentration of photosynthetic pigments [22, 41, 42], changes in fine structure of chloroplasts [43], inhibition in electron transport system [44], changes in lipid and protein composition of thylakoid membrane [24, 45], and inhibition of the activities of Calvin cycle enzymes [46]. Rate of photosynthesis is also determined by substomatal CO2 concentration that is controlled by stomatal conductance (g s) [47]. In the present study, Pb stress caused a significant increase in transpiration rate (E) and stomatal conductance (g s), but in contrast, water use efficiency (A/E) and intrinsic water use efficiency (A/g s) decreased markedly with increasing concentration of external Pb. Thus, it can be concluded that reduction in photosynthetic rate (A) may have been due to factors other than stomatal limitation (g s).
Lead (Pb) stress significantly reduced the concentrations of K+ and Cu2+ in the roots as well as in the leaves of both maize genotypes in the present study. High levels of Pb have been reported to cause severe imbalance in concentration of mineral nutrients in different plant tissues [48, 49] as it competes with the entry of different cations (K+, Cu2+, Ca2+, Mg2+) in the root system [22]. As discussed earlier, Pb stress can result in altered lipid and protein composition of biological membranes that can increase leakage of cations such as K+ and Cu2+ from the root cells [50]. In view of some earlier studies, it is evident that only a little proportion of Pb absorbed by the roots is translocated via xylem into the aboveground plant organs [51, 52]. However, leaves can accumulate a significant amount of Pb in a concentration-dependent manner over time as observed in the present study [48, 53]. Therefore, reduction in accumulation of these nutrients in leaves under Pb stress might be a direct consequence of buildup of high concentration of Pb in the leaves that competitively reduced the concentration of K+ and Cu2+.
Conclusion
Overall, application of Pb stress decreased seed germinability and plant growth of both maize genotypes. Moreover, disturbance in uptake and transport of mineral nutrition and decrease in photosynthesis were observed with increase in concentration of Pb in the growth medium. Cv. EV-1098 was more tolerant to Pb toxicity as compared to cv. EV-77 because it showed relatively less decrease in growth, photosynthetic performance, and K+ and Cu2+ contents and less accumulation of Pb in the roots and leaves.
References
Valko M, Morris H, Cronin MT (2005) Metals toxicity and oxidative stress. Curr Med Chem 12:1161–1208
Blaylock MJ (1999) Enhanced accumulation of lead in Indian mustard by soil applied chelating agents. Environ Sci Technol 31:860–865
Athar R, Ahmad M (2002) Heavy metal toxicity: effect on plant growth and metal uptake by wheat, and on free living Azotobacter. Water Air Soil Pollut 138:165–180
Belyaeva ON, Haynes RJ, Birukova OA (2005) Barley yield and soil microbial and enzyme activities as affected by contamination of two soils with lead, zinc or copper. Biol Fertil Soils 41:85–94
Opeolu BO, Adenuga OO, Ndakidemi PA, Olujimi OO (2010) Assessment of phyto-toxicity potential of lead on tomato (Lycopersicon esculentum L) planted on contaminated soils. Int J Physical Sci 5:068–073
Watanabe MA (1997) Phytoremediation on the brink of commercialization. Environ Sci Technol 31:182–186
Eick MJ, Peak JD, Brady PV, Pesek JD (1999) Kinetics of lead adsorption and desorption on goethite: residence time effect. Soil Sci 164:28–39
Yang YY, Jung JY, Song WY, Suh HS, Lee Y (2000) Identification of rice varieties with high tolerance or sensitivity to lead and characterization of the mechanism of tolerance. Plant Physiol 124:1019–1026
Nedelkoska TV, Doran PM (2000) Characteristics of metal uptake by plants species with potential for phytoremediation and phytomining. Min Eng 13:549–561
Hall JL (2002) Cellular mechanism for heavy metal detoxitification and tolerance. J Exp Bot 53:1–11
He ZL, Yang XE, Stoffella PJ (2005) Trace elements in agroecosystems and impacts on the environment. J Trace Elemen Med Biol 19:125–140
Walker CH, Hopkin SP, Sibly RM, Peakall DB (1996) Principles of ecotoxicology. Taylor and Francis, Bristol
Patra M, Bhowmik N, Bandopadhyay B, Sharma A (2004) Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ Exp Bot 52:199–223
Kabata-Pendias A, Pendias H (1999) Biogeochemistry of trace elements. Panstwowe Wydawnictvo Naukowe Publishers, Warszawa
Kumar PBAN, Dushenkov V, Motto H, Raskin I (1995) Phytoextraction: the use of plants to remove heavy metals from soils. Environ Sci Technol 29:1232–1238
Piechalak A, Tomaszewska B, Baralkiewicz D, Malecka A (2002) Accumulation and detoxification of lead ions in legumes. Phytochemistry 60:153–162
Verma S, Dubey RS (2003) Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci 164:645–655
Burton KW, Morgan E, Roig A (1984) The influence of heavy metals on growth of Sitka-spruce in South Wales forests. Π. Greenhouse experiments. Plant Soil 78:271–282
Arduini I, Godbold DL, Onnis A (1994) Cadmium and copper change root growth and morphology of Pinus pinea and Pinus pinaster seedlings. Physiol Plant 92:675–680
Wozny A, Schneider J, Gwozdz EA (1995) The effects of lead and kinetin on greening of barley leaves. Biol Plant 37:541–552
Uveges JL, Corbett AL, Mal TK (2002) Effects of Pb contamination on the growth of Lythrum salicaria. Environ Pollut 120:319–323
Sharma P, Dubey RS (2005) Lead toxicity in plants. Brazil J Plant Physiol 17:35–52
Przymusinski R, Spychala M, Gwozdz EA (1991) Inorganic lead changes growth polypeptide pattern of lupin roots. Biochem Physiol Pflan 187:51–57
Stefanov K, Seizova K, Popova I, Petkov VL, Kimenov G, Popov S (1995) Effects of lead ions on the phospholipids composition of in leaves of Zea mays and Phaseolus vulgaris. J Plant Physiol 147:243–246
Ahmad MSA, Hussain M, Ijaz S, Alvi AK (2008) Photosynthetic performance of two mung bean (Vigna radiata) cultivar under lead and copper stress. Int J Agric Biol 10:167–172
Wierzbicka M, Obidzinska J (1998) The effect of lead on seed imbibition and germination in different plant species. Plant Sci 137:155–171
Shaukat SS, Mushtaq M, Siddiqui ZS (1999) Effects of cadmium, chromium and lead on seed germination, early seedling growth and phenolic contents of Parkinsonia aculeate L. and Pennisatum americanum (L.) Schumann. Pak J Biol Sci 2:1307–1313
Iqbal MZ, Shazia Y (2004) Reduction of germination and seedling growth of Leucaena leucocephala caused by lead and cadmium individually and combination. Ekologia Braslava 23:162–168
Farooqi ZR, Iqbal MZ, Kabir M, Shafiq M (2009) Toxic effects of lead and cadmium on germination and seedling growth of Albizia lebbeck (L.) Benth. Pak J Bot 41:27–33
Vange V, Hevchand I, Vandvik V (2004) Does seed mass and family affect germination and juvenile performance in Knautia arvensis? A study using failure time methods. Acta Oecologia 25:169–178
Hoagland DR, Arnon DI (1950) The water culture method for growing plant without soil. California Agri. Exp. Sta. Cir. No. 347. University of California Berkley Press, CA., p 347
Munzuroglu O, Geckil H (2002) Effects of metals on seed germination, root elongation, and coleoptile and hypocotyl growth in Triticum aestivum and Cucumis sativus. Arch Environ Contam Toxicol 43:203–213
Tomulescu IM, Rodoviciu EM, Merca VV, Tuduce AD (2004) Effect of copper, zinc lead and their combinations on germination capacity of two cereals. J Agric Sci Debrecen 15:39–42
Kabir M, Iqbal MZ, Shafiq M, Farooqi ZR (2008) Reduction in germination and seedling growth of Thespesia populnea L., caused by lead and cadmium treatments. Pak J Bot 40:2419–2426
Datta JK, Ghanty S, Banerjee A, Moudal NK (2009) Impact of lead on germination physiology of certain wheat cultivar (Triticum aestivum L.). J Ecophysiol Occup Hlth 9:145–151
Mesmar MN, Jaber K (1991) The toxic effect of lead on seed germination and growth of wheat and lens. Acta Biol Hung 42:331–344
Donghua L, Jiang W, Liu C, Xin C, Hou W (2000) Uptake and accumulation of lead by roots, hypocotyls and roots of Indian mustard. J Bioresource Technol 71:273–277
Lerda D (1992) The effect of lead on Allium cepa L. Mut Res 281:89–92
Goldbold DL, Kettner C (1991) Lead influences on root growth and mineral nutrition of Picea abies seedling. Plant Physiol 139:95–99
Sharifah BA, Hishashi O (1992) Effect of lead, cadmium and zinc on the cell elongation of Impatiens balsmina. Environ Exp Bot 32:439–448
Drazkiewicz M (1994) Chlorophyll—occurrence, functions, mechanism of action, effects of internal and external factors. Photosynthetica 30:321–331
Hussain M, Ahmad MSA, Kausar A (2006) Effect of lead and chromium on growth, photosynthetic pigments and yield components in mash bean [Vigna mungo (L.) Hepper]. Pak J Bot 38:1389–1396
Fargasova A (2001) Phytotoxic effects of Cd, Zn, Pb, Cu and Fe on Sinapsis alba L. seedling and their accumulation in roots and shoots. Biol Plant 44:471–473
Rashid A, Camm EL, Ekramoddoullah KM (1994) Molecular mechanism of action of Pb and Zn2+ on water oxidizing complex of photosystem Π. FEBS Lett 350:296–298
Skorzynska-Polit E, Baszynski T (1997) Differences in sensitivity of the photosynthetic apparatus in Cd-stressed runner bean plants in relation to their age. Plant Sci 128:11–21
Parys E, Romanowaska E, Siedlecka M, Poskuta J (1998) The effect of lead on photosynthesis and respiration in detached leaves and in mesophyll protoplasts of Pisum sativum. Acta Physiol Plant 20:313–322
Kosobrukhov A, Knyazeva I, Mudrik V (2004) Plantago major plants responses to increase content of lead in soil: growth and photosynthesis. Plant Grow Regul 42:145–151
Kabata-Pendias A, Pendias H (1992) Trace elements in soil and plants. CRC Press, Boca Raton
Eun SO, Youn HS, Lee Y (2000) Lead disturbs microtubule organization in the root meristem of Zea mays. Physiol Plant 110:357–365
Malkowski E, Kita A, Galas W, Karcz W, Kuperberg M (2002) Lead distribution in corn seedlings (Zea mays L.) and its effect on growth and the concentrations of potassium and calcium. Plant Grow Regul 37:69–76
Jones LHP, Clement CR, Hopper MJ (1973) Lead uptake from solution by perennial ryegrass and its transport from roots to shoots. Plant Soil 38:403–414
Fargasova A (2004) Toxicity comparison of some possible toxic metals (Cd, Cu, Pb, Se, Zn) on young seedlings of Sinapsis alba L. Plant Soil Environ 50:33–38
Nwosu JU, Harding AK, Linder G (1995) Cadmium and lead uptake by edible crops grown in a silt loam soil. Bull Environ Contam Toxicol 54:570–578
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ahmad, M.S.A., Ashraf, M., Tabassam, Q. et al. Lead (Pb)-Induced Regulation of Growth, Photosynthesis, and Mineral Nutrition in Maize (Zea mays L.) Plants at Early Growth Stages. Biol Trace Elem Res 144, 1229–1239 (2011). https://doi.org/10.1007/s12011-011-9099-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-011-9099-5