
Abstract
Two microalgae species (Scenedesmus obliquus and Neochloris oleoabundans) were cultivated in closed sleeve photobioreactors in order to select the best oil producer for further large-scale open raceway pond cultivations, aiming at biofuel production. Scenedesmus obliquus reached a higher maximum biomass concentration (1.41 g l−1) with a lower lipid content (12.8% w/w), as compared to N. oleoabundans [maximum biomass concentration of 0.92 g l−1 with 16.5% (w/w) lipid content]. Both microalgae showed adequate fatty acid composition and iodine values as substitutes for diesel fuel. Based on these results, N. oleoabundans was selected for further open raceway pond cultivations. Under these conditions, N. oleoabundans reached a maximum biomass concentration of 2.8 g l−1 with 11% (w/w) of lipid content. A high correlation between the Nile Red fluorescence intensity measured by flow cytometry and total lipid content assayed by the traditional gravimetric lipid analysis was found for both microalgae, making this method a suitable and quick technique for the screening of microalgae strains for lipid production and optimization of biofuel production bioprocesses. Medium growth optimization for enhancement of microalgal oil production is now in progress.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Due to the diminishing petroleum reserves and environmental consequences of exhaust gases from fossil diesel, production of alternative fuel materials has attracted wide attention [1]. The current reliance on petro-based fuels and chemicals is no longer sustainable, as the energy demand is projected to grow more than 50% by 2025 [2]. Renewable, carbon neutral, transport fuels are necessary for environmental and economic sustainability. The ecological footprint of energy generation will reside in a multifaceted approach that includes nuclear, solar, hydrogen, wind, fossil fuels (from which carbon is sequestered), and biofuels. Biodiesel derived from oil crops is a potentially renewable and carbon-neutral alternative to petroleum fuels. Unfortunately, biodiesel derived from oil crops, waste cooking oil, and animal fat can realistically satisfy only a small fraction of the existing demand for transport fuels [3].
Microalgae, a renewable energy source that has not been fully exploited, have been suggested as very good candidates for fuel production, as they appear to be the only renewable biofuel that is capable of meeting the global demand for transport fuels [3]. Autotrophic microalgae are sunlight-driven cell factories that convert carbon dioxide into potential biofuels, foods, feeds, and high-value products. They use a photosynthetic process similar to that of higher developed plants and can complete an entire growing cycle every few days, resulting in a much higher oil productivity as compared to oil crops. In addition, they are efficient CO2 fixers. The ability of microalgae to fix CO2 has been proposed as a method of removing CO2 from flue gases from power plants and, thus, can be used to reduce emissions of greenhouse gases [1]. In addition, microalgae production is nonseasonal, with the possibility of harvesting daily; may take place on nonarable land (e.g., desert and seashore lands, saltpans, rocky and sandy areas); does not compete with food crops; and can use nonpotable water. Raceways may be used in such conditions. Because of their simplicity and the low investments costs involved, raceway ponds are often used for biodiesel production [3].
Despite all of these benefits, microalgal lipid production is strongly dependent on environmental factors. Therefore, it is essential to monitor cell lipid content during the biotechnological process. Accurate estimates of cell lipid content are essential so that informed decisions on process control can be made. Traditional analysis of lipid content in microalgae are usually performed by solvent extraction and gravimetric determination [4, 5], and further analysis and characterization is carried out by gas chromatography (GC) or high-performance liquid chromatography [6]. These steps are time consuming and generate high amounts of waste (organic solvent), which are harmful to the environment if not recycled by distillation. In addition, enough biomass must be obtained for subsequent lipid extraction and derivatization. Moreover, lipid content data are usually only available a considerable time after the sample is taken, too late for alterations to be made to process control.
Multiparameter flow cytometry can monitor total cell lipid content, in situ, in real time, and with a high degree of statistical resolution, during the microalgal growth. If the cell lipid content information is available during the time course of the bioprocess, decisions on process control strategy can be made (for example, adjusting the carbon/nitrogen ratio of the medium, based on the output from the flow cytometry) so that lipid productivity can be increased.
Lipid measurements have been proposed previously using the Nile Red fluorescent stain for spectrofluorometric quantification of lipids. Its fluorescence is produced in highly hydrophobic environments and quenched in hydrophilic ones [7–10]. In this work, two microalgal species (Neochloris oleoabundans and Scenedesmus obliquus), known as promising oil producers due to their high cellular lipid content [11, 12], were grown in closed plastic sleeve photobioreactors in order to evaluate their lipid production capacity for biofuel production. The microalgal species that showed the highest lipid content was further grown in a scale-up outdoor raceway pond. During all the microalgal growths, biomass and lipid content were monitored. A correlation between the microalgal lipid content assayed by the traditional gravimetric method [4] and the Nile Red fluorescence intensity measured by flow cytometry [7] was established. Fatty acid profile and iodine value were determined as important parameters to evaluate the final biodiesel quality.
Materials and Methods
Organisms
Freshwater microalgae N. oleoabundans and S. obliquus were purchased from the UTEX collection (Texas University of Austin, USA) and the Coimbra University algotec collection, respectively. Both genera belong to Chlorophyceae class and Chlorococcaceae family [13].
Growth Conditions
Closed Sleeve Photobioreactors
The two microalgae species were grown previously in 50-L plastic sleeve photobioreactors in order to select the best microalga for further experiments in outdoor raceway ponds. The cultures were maintained in a semicontinuous mode, being stirred continuously by bubbling filtered air, in Bristol medium [14] at 25°C. The cultures were kept illuminated with six fluorescent lamps (Philips TL-DM 36W/54-765) at an irradiance level of 150 μE m−2 s−1 with photoperiod 24:0 h (light/dark) during at least 103 days.
Outdoor Raceway Pond
The microalga species that depicted the highest lipid content was further grown in a 2.5-m2 polyethylene outdoor raceway pond. The depth of the culture was 15 cm. Mechanical agitation was obtained by a paddle-wheel that maintained a water velocity of 25 cm/s. Harvesting and dilution were routinely performed to maintain the adequate optical density values. The experiment was carried out for 59 days from July to August 2007 and the culture was maintained in a semicontinuous mode.
Growth Evaluation
Growth was periodically (at least every 2 days) evaluated by optical density at 540 nm in duplicate, which was converted into dry cell weight per liter of culture by a regression equation. Closed sleeve bioreactor experiments lasted at least 103 days, while open raceway experiment lasted 59 days.
Specific Growth Rate and Volumetric Biomass Productivity Calculations
Specific growth rate μ (day−1) was calculated as follows:
where X (g l−1) was the biomass produced during the time interval t 2 − t 1. Volumetric biomass productivity r X (g l−1 day−1) was calculated as follows:
where X 1 and X 2 were the biomass concentration (g l−1) on days t 1 and t 2, respectively.
Lipid Content
Total lipid content was evaluated by traditional gravimetric analysis [4] and by multiparameter flow cytometry using the fluorescent stain Nile Red [7]. Microalgal cells were stained with 5 mM Nile Red 50 μl of a working solution of NR and acetone (0.1 mg ml−1) was added to 1 ml of a cell suspension (105–106 cells ml−1). This mixture was gently vortexed and incubated for 10 min at 37°C in darkness. Nile Red florescence was determined using a FACScan flow cytometer (Becton-Dickinson Instruments) equipped with a 488-nm argon laser. Upon excitation by a 488-nm argon laser, Nile Red exhibits yellow-gold and red fluorescences when dissolved in neutral and polar lipids, which were detected in the FL2 and FL3 channels, respectively. Nonstained cells were used as an autofluorescence control (AF). The total fluorescence corresponding to total cellular lipids was determined as the sum of the ratios FL2/AF and FL3/AF. A correlation between total microalgal lipid content assayed by the Bligh and Dyer method [4] and Nile Red fluorescence intensity was established by analyzing at least four samples of each microalga with different lipid contents.
Volumetric Lipid Productivity Calculation
Volumetric lipid productivity was calculated as:
where L 1 and L 2 were the lipid concentration (g l−1) on days t 1 and t 2, respectively.
Lipid Characterization
Fatty Acid Composition
To determine the fatty acid composition of each raw material, oil samples were chemically methylated using the borum trifluoride method described in EN ISO 5509 [15]. The organic phase obtained was analyzed by gaseous chromatography using a CP-3800 GC (Varian, USA) equipped with 30 m DB-WAX (J&W, Agilent) capillary column (0.25 mm of internal diameter and 0.25 μm of film thickness). Injector (split 1:100) and detector (flame ionization) temperatures were kept constant at 250°C. The oven temperature program started at 180°C for 5 min, increased at 4°C min−1 until 220°C, and was kept constant at this temperature for 25 min. The velocity of the carrier gas, He, was kept constant at 1 ml min−1. Fatty acid composition was calculated as percentage of the total fatty acid present in the sample determined from the peak areas.
Iodine Value
Oils from microalgae were characterized in terms of iodine value from its fatty acid composition, according to EN 14214: Annex B [12] [16].
Results and Discussion
Light Scatter Measurements
It has been reported that the marine microalga Crypthecodinim cohnii can be detected from the background on the basis of their intrinsic light scattering properties in forward angle light scatter (FALS) and right angle light scatter (RALS) [17, 18].
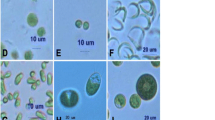
Figure 1 shows FALS vs RALS measurements of S. obliquus and N. oleoabundans, respectively, grown in plastic sleeve photobioreactors. Both microalgae show elliptical and broaden distributions with respect to FALS and RALS flow cytometric analysis, suggesting cell size and shape heterogeneity, which can be due to the different G1–S–G2–M cell phases of these two eukaryotic microalgae.

Forward (FALS) vs right (RALS) angle slight scatter for a S. obliquus and b N. oleoabundans. Scenedesmus obliquus plot (A) shows two populations A and B with respect to RALS
However, while S. obliquus showed two subpopulations with respect to RALS, N. oleoabundans showed only one elliptical distribution with respect to RALS. Such differences may be explained on their reproduction cycle. Scenedesmus obliquus reproduction is by autocolony formation, which parental cell forms a miniature colony that is liberated through a tear in the parental wall, and the wall of the parent colony remains [19]. Those colony parent walls may explain subpopulation A (Fig. 1a), as they should have a lower RALS signal, which is dependent on the cytoplasmic complexity. Neochloris oleoabundans reproduction is by zoospores without membranes and autospores [13]. Therefore, flow cytometric analysis based on changes in light scatter allows the differentiation between these two microalgae species.
Sleeve Photobioreactor Experiments
Figures 2 and 3 show the growth profiles of biomass and daily productivity for S. obliquus and N. oleoabundans cultivated in sleeve photobioreactors. It can be seen that, while S. obliquus attained a maximum biomass concentration of 1.96 g l−1 at t = 91 days and productivity of 0.26 g l−1 day−1, at t = 64 days, N. oleoabundans attained biomass and productivity maximums of 1.87 g l−1 at t = 87 days and 0.19 g l−1 day−1 at t = 54 days, respectively. However, the specific growth rate (μ) related to the first 20 days of growth were similar for both microalgae (0.08 and 0.07 day−1 for S. obliquus and N. oleoabundans, respectively).

Dry cell weight and biomass productivity profiles during S. obliquus semicontinuous cultivation in a sleeve photobioreactor. Dry cell weights are affected by a relative error not exceeding 10% (n = 2). ------ Dilution of the culture. –··–·· Centrifugation of the culture followed by dilution with fresh medium

Dry cell weight and biomass productivity profiles during N. oleoabundans semicontinuous cultivation in a sleeve photobioreactor. ------ Dilution of the culture. –··–·· Centrifugation of the culture followed by dilution with fresh medium. Dry cell weights are affected by a relative error not exceeding 10% (n = 2)
Regarding the lipid content, N. oleoabundans showed the highest cellular lipid content, attaining 16.5% (w/w) after 74 days, while S. obliquus only reached 12.8% (w/w) at t = 71 days (Tables 1 and 2). The former microalga showed a lower lipid content as compared to that reported by Tornabene [11] (35–54% of cell dry weight). Indeed, microalgal lipid content is strongly dependent on environmental factors, including growth medium composition, which can explain these differences in cellular lipid content. However, S. obliquus depicted a similar lipid content to that reported by [20] (11.7%).
It has been reported that light favored growth and lipid synthesis in microalgae [21]. However, as S. obliquus culture attained higher biomass concentrations, the greater number of cells might have increased the self-shading effect, reducing the available light intensity per cell; thus, reducing the cellular lipid content. Despite the higher lipid concentration and productivity depicted by S. obliquus (Table 1), the higher lipid content shown by N. oleoabundans made this microorganism eligible for further large-scale cultivations using open raceway pond, as in a large-scale biofuel production process, a higher microalgae cellular lipid content will reduce the costs of the downstream processing steps, with particular emphasis on the oil recovery step.
Nile Red Fluorescence Correlation with Cellular Lipid Content
In order to obtain a correlation between Nile Red fluorescence and cell lipid content, samples from S. obliquus and N. oleoabundans cultures were stained with Nile Red and analyzed by flow cytometry. A high correlation between total NR fluorescence (FL2/AF+FL3/AF) and lipid content was obtained for both microalgae (Fig. 4a and b, S. obliquus R 2 = 0.80; N. oleoabundans R 2 = 0.84). In this way, microalgal cellular oil content can be monitored through total Nile Red fluorescence flow cytometric analysis during microalgal growth, an easier and quicker method for lipid analysis as compared to traditional gravimetric lipid analyses usually used to assay microalgal lipids for biodiesel production [5, 19, 22].

Linear correlations between total Nile Red fluorescence and total microalgal lipid content assayed by Bligh and Dyer method (1959) for a S. obliquus and b N. oleoabundans
Characteristics of Microalgal Oil
For user acceptance, microalgal biodiesel will need to comply with existing standards. To assess the potential of the microalgae oils as raw material for a substitute of diesel fuel, the properties of the microalgal lipids such as fatty acid composition and iodine value were evaluated. In the European Union, separate standards exist for biodiesel intended for vehicle use (European Standard EN 14214) [16].
Table 3 shows the fatty acid composition of S. obliquus and N. oleoabundans grown in sleeve bioreactors. Both microalgal lipids are mainly composed of unsaturated fatty acids (54.6% and 64.7% w/w for S. obliquus and N. oleoabundans, respectively), and a significant percentage of palmitic acid (16:0) was also present (∼20%). Among the unsaturated fatty acids, special attention should be taken in relation to the linolenic methyl ester (18:3) and polyunsaturated methyl esters (≥4 double bonds) content due to the EN 14214 that specifies limits of 12% and 1%, respectively. The microalga N. oleoabundans present a value of 17.5% and 9.3% w/w for linolenic and polyunsaturated fatty acids, respectively, which are not impeditive for a good quality biodiesel if associated with other oils. In addition, the extent of unsaturation of microalgal oil and its content of fatty acids with more than four double bonds can be reduced easily by partial catalytic hydrogenation of the oil [3].
Total unsaturation of an oil is indicated by its iodine value. Standard EN 14214 requires the iodine value of biodiesel not exceeding 120 g iodine/100 g biodiesel, respectively. The iodine values for S. obliquus and N. oleoabundans were 68.4 and 87.6, respectively, which meet both standard specifications.
Outdoor Raceway Pond Experiment
Figure 5 depicts the growth of N. oleoabundans when cultivated in an outdoor raceway pond. Higher biomass concentrations were attained in this system (maximum of 2.8 g l−1 at t = 21 days) as compared to the sleeve cultivation (maximum of 1.8 g l−1 at t = 82 days) (Fig. 3). This is not surprising since the mass transfer in the plastic sleeves should be lower than that in the raceway due to stagnant regions present in the former, which were mainly located in the plastic folds. The specific growth rate related to the first 20 days of growth (0.18 day−1) was more than double that of the calculated for N. oleoabundans when growing in sleeve photobioreactors (0.07 day−1).

Dry cell weight and biomass productivity of N. oleoabundans growing in an open raceway, in a semicontinuous mode. The experiment was carried out for 59 days, from July to August 2007. –··–·· Centrifugation of the culture followed by dilution with fresh medium. Dry cell weights are affected by a relative error not exceeding 10% (n = 2)
The main environmental variables affecting microalgal lipid production are related to radiant energy (light and temperature), both related to solar radiation [23]. Therefore, outdoor raceways are more dependent on environmental and seasonal factors than sleeve photobioreactors wherein these factors may be controlled [10]. This may explain the higher cellular lipid content of the microalgal cells observed in the N. oleoabundans sleeve cultivation (maximum of 16.5% w/w at t = 74 days, Table 1), as compared to the N. oleoabundans lipid content obtained in open raceway cultivations (11% w/w at t = 9 days). However, the higher biomass production observed in the raceway pond resulted in a higher lipid concentration and productivity [maximums of 26.7 g l−1 at t = 21 days and 0.03 g l−1 day−1 (4.8 g m−2 day−1) between t = 17 days and t = 21 days, respectively] as compared to the N. oleoabundans sleeve cultivation (Table 4).
The maximum lipid content of N. oleoabundans in the raceway open pond (11% w/w) was lower than that reported by Chisti [24] (20% w/w). Nevertheless, its oil yield (21,523 l ha−1) is still higher than that depicted by various crops (corn, soybean, canola, jatropha, and coconut) [3] due to the higher biomass productivities of the microalgae. However, the low lipid content obtained in this work can be improved by changing the microalgae growth conditions, namely C/N ratio. Such studies are now in progress.
Conclusions
Although the oils from both microalgae showed adequate fatty acid composition and iodine values as substitutes for diesel fuel, N. oleoabundans was selected for large-scale open raceway cultivations due to its higher cellular lipid content. A high correlation between the Nile Red fluorescence intensity measured by flow cytometry and total lipid content assayed by the gravimetric lipid analysis was found for both microlagae, making this method a suitable technique for quick screening of microalgae strains for lipid production and optimization of biofuel production bioprocesses. Growth conditions optimization for microalgal oil production enhancement is now in progress.
References
Ma, F. R., & Hanna, M. A. (1999). Bioresource Technology, 70, 1–15. doi:10.1016/S0960-8524(99)00025-5.
Patil, V. (2008). Current Science, 92, 707.
Chisti, Y. (2007). Biotechnology Advances, 25, 294–306. doi:10.1016/j.biotechadv.2007.02.001.
Bligh, E. G., & Dyer, W. J. (1959). Journal of Biochemistry and Physiology, 37, 911–917.
Miao, X., & Wu, Q. (2006). Bioresource Technology, 97, 841–846. doi:10.1016/j.biortech.2005.04.008.
Elsey, D., Jameson, D., Raleigh, B., & Cooney, M. J. (2007). Fluorescent measurement of microalgal neutral lipids. Journal of Microbiological Methods, 68, 639–642. doi:10.1016/j.mimet.2006.11.008.
de la Jara, A., Medonza, H., Martel, A., Molina, C., Nordströn, L., de la Rosa, V., & Díaz, R. (2003). Journal of Applied Phycology, 15, 433–438. doi:10.1023/A:1026007902078.
Fowler, S. D., Brown, W. J., Warfel, J., & Greenspan, P. (1979). Journal of Lipid Research, 28, 1225–1232.
Greenspan, P., Mayer, E. P., & Fowler, S. D. (1985). The Journal of Cell Biology, 100, 965–973. doi:10.1083/jcb.100.3.965.
Pulz, O. (2001). Applied Microbiology and Biotechnology, 57, 287–293. doi:10.1007/s002530100702.
Tornabene, T. G. (1983). Enzyme and Microbial Technology, 5, 435–440. doi:10.1016/0141-0229(83)90026-1.
Sheehan, J., Dunahay, T., Benemann, J., & Roessler, P. (1998). A look back at the US Department of Energy’s Aquatic Species Program—Biodiesel from Algae. National Renewable Energy Laboratory, Golden, Colorado, USA.
Bourrelly, P. (1972). Les algues vertes. Tome I, Editions N. Boubée & Cie, Paris.
Bold, H. C. (1949). Bulletin of the Torrey Botanical Club, 76, 101–108. doi:10.2307/2482218.
European Standard EN ISO 5509 (2000).
European Standard EN 14214 (2003).
da Silva, T. L., & Reis, A. (2008). Journal of Industrial Microbiology & Biotechnology, 35, 875–887.
Yeung, P. K. K., & Wong, J. T. Y. (2003). Protoplasma, 220, 173–178. doi:10.1007/s00709-002-0039-2.
Richmond (1986). Handbook of microalgal mass culture. Boca Raton: CRC.
Choi, K. J., Kakhost, Z., Barzana, E., & Karel, M. (1987). Food Technology, 11, 117–128.
Zhukova, N. V., & Titlyanov, E. A. (2006). Botanica Marina, 49, 339–346. doi:10.1515/BOT.2006.041.
Xu, H., Miao, X., & Wu, Q. (2006). Journal of Biotechnology, 126, 499–507. doi:10.1016/j.jbiotec.2006.05.002.
Nalewajko, C., & Voltolina, D. (1985). Canadian Journal of Fisheries and Aquatic Sciences, 43, 1163–1170.
Chisti, Y. (2008). Trends in Biotechnology, 26, 351–352. doi:10.1016/j.tibtech.2008.04.002.
Acknowledgements
The authors wish to acknowledge Doutora Narcisa Bandarra from IPIMAR for the fatty acid analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
da Silva, T.L., Reis, A., Medeiros, R. et al. Oil Production Towards Biofuel from Autotrophic Microalgae Semicontinuous Cultivations Monitorized by Flow Cytometry. Appl Biochem Biotechnol 159, 568–578 (2009). https://doi.org/10.1007/s12010-008-8443-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-008-8443-5