
Abstract
Sugar cane bagasse was used as substrate for cellulase production using Trichoderma reesei RUT C30, and the culture parameters were optimized for enhancing cellulase yield. The culture parameters, such as incubation temperature, duration of incubation, and inducer concentration, were optimized for enhancing cellulase yield using a Box–Behnken experimental design. The optimal level of each parameter for maximum cellulase production by the fungus was determined. Predicted results showed that cellulase production was highest (25.6 FPAase units per gram dry substrate) when the inducer concentration was 0.331 ml/gds, and the incubation temperature and time were 33 °C and 67 h, respectively. Crude inducer generated by cellulase action was found to be very effective in inducing cellulases. Validation of predicted results was done, and the experimental values correlated well with that of the predicted.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
There is an increased demand for alternative liquid fuels world wide due to the rapidly depleting petroleum resources. Liquid fuels for transportation can be derived from lignocellulosic biomass by enzymatic hydrolysis and subsequent fermentation to produce bioethanol [1]. Cellulases are the enzymes primarily implicated in the saccharification process, and the production economics of bioethanol is largely dependent on the cost of cellulases [2]. Substrate costs account for a major fraction of the costs of cellulase production, and the use of cheap biomass resources as substrates can help to reduce cellulase prices [3]. Also the use of cheaper technologies like solid-state fermentation (SSF) can further improve the production economics. SSF technology results in an enzyme preparation, which is more concentrated and hence best suited for biomass conversion applications [4].
Sugar cane bagasse is a low-cost and abundant biomass material [5] containing about 27–54% cellulose [6], which can serve as a potent substrate for cellulase production. There have been several studies on the use of sugar cane bagasse as a substrate in cellulase production both under submerged and SSF [7–9] but a systematic study on the effect of process parameters considering their interaction effects under SSF is rare. We had previously produced cellulases on wheat-bran as substrate under SSF using Trichoderma reesei RUT C30 and could improve the yield by optimizing fermentation conditions [10]. Because sugar cane bagasse could be another important biomass resource suitable for cellulase production, we have tried to optimize the process parameters for improving cellulase production by T reesei under SSF on sugar cane bagasse a response surface Box–Behnken design.
Materials and Methods
Microorganism and Inoculum Preparation
The cellulase hyper-producing fungus T. reesei RUT-C30, was a kind gift from Prof. George Scakacs, Technical University of Budapest, Hungary. Culture was maintained on potato dextrose agar slants and was incubated at 30 °C. The fully sporulated slants obtained after 5 days were either used immediately or stored at 4°C in refrigerator. A conidial suspension of T reesei was prepared by adding 5 ml distilled water supplemented with 0.1% (v/v) Tween 80 into agar slants of the fungal culture and dislodging the spores into it by gentle pipetting. The spore count was adjusted to 107 spores/ml, and 1 ml of this suspension was used to inoculate 100 ml of mineral salts medium containing glucose. The inoculated medium was incubated at 30 °C for 4 days. Protein content of this seed culture was estimated by the method of Herbert et al. [11]. Dilutions of the seed culture was done with sterile saline so as to obtain a total protein content of 150 mg/ml, and 1 ml of this suspension was used to inoculate each flask.
Enzyme Production and Assay
Substrate
Sugarcane bagasse was air dried, milled, and size fractionated. Bagasse with a particle size range 500–1,000 μm was used as substrate for SSF without any pretreatment.
Preparation of Crude Inducer
Crude inducer for cellulase was prepared from cellulose by the action of whole Trichoderma cellulase. A 10% cellulose (SolkaFloc , Sigma) solution in 0.05M citrate buffer was prepared and was incubated with cellulase (Zytex, India) at a concentration of 15 FPU/ml of reaction mixture at 50 °C in a water-bath for 12 h after which the reaction was terminated by incubating for 5 min in boiling water bath. The reaction mixture was centrifuged to recover the supernatant. The supernatant was sterilized by filtration through a 0.22-μ membrane filter (Millipore) and was kept frozen until used. Required amount of the reaction mixture was added to the substrate when it was used as inducer.
Enzyme Production
Three grams of the substrate was weighed into 250-ml Erlenmeyer flasks and was moistened with mineral salt medium to attain 70% initial moisture content. The basal mineral salts solution used for the experiment had the following composition (g/l): KH2PO4, 5; NH4NO3, 5; MgSO4.7H2O, 21; urea, 2; CaCl2, 1; Peptone, 5; NaCl, 5; Tween-80, 0.5; and trace elements: FeSO4.7H2O, 0.005; MnSO4.7H2O, 0.001; ZnSO4.7H2O, 0.001; and CoCl2, 0.0002. The initial pH of the salt solution was adjusted to 5. The flasks were sterilized by autoclaving at 121 °C for 15 min at 15 lbs pressure, and after cooling, filter sterilized crude inducer preparation was added. The medium was inoculated with 1 ml of the seed inoculum (150 mg/ml total protein). The contents were mixed thoroughly and were incubated under controlled conditions of temperature and humidity. Incubation was continued for the duration indicated in the experimental designs, and at the end of incubation period, the enzyme was recovered by extraction with 0.05 N citrate buffer (pH 4.8). The extract was centrifuged to remove debris at 6,000 rpm for 10 min at 4 °C, and the supernatant was used as the crude enzyme sample.
Enzyme Assay
Filter paper assay [12] was used to estimate total cellulase activity in the crude enzyme preparation as given below. A rolled Whatman no. 1 filter paper strip of dimension 1.0 × 6 cm (50 mg) was placed into each assay tube. The filter paper strip was saturated with 0.5 ml of Na–citrate buffer (0.05 M, pH 4.8) and was equilibrated for 10 min at 50 °C in a water bath. Half milliliter of an appropriately diluted (in Na–citrate buffer, 0.05M; pH 4.8) enzyme was added to the tube and incubated at 50 °C for 60 min. Appropriate controls were also run along with the test. At the end of the incubation period, tubes were removed from the water bath, and the reaction was stopped by addition of 3 ml of 3,5-dinitrosalicylic acid reagent per tube. The tubes were incubated for 5 min in a boiling water bath for color development and were cooled rapidly. The reaction mixture was diluted appropriately and was measured against a reagent blank at 540 nm in a UV-VIS spectrophotometer. The concentration of glucose released by enzyme was determined by comparing against a standard curve constructed similarly with known concentrations of glucose. One unit of cellulase activity was defined as the amount of enzyme required for liberating 1 μM of glucose per milliliter per minute and was expressed as U/gds (units per gram dry substrate).
Optimization of Cellulase Production
The SSF production of cellulase on sugar cane bagasse is influenced by various process variables including media components and environmental parameters. Incubation temperature, time, and the concentration of inducer were identified as significant parameters affecting cellulase production by an initial screening of parameters. The levels of these variables were optimized for enhancing the cellulase yield using a response surface Box–Behnken experiment design [13]. The design matrix with 17 experimental runs in two blocks with five replicates of the midpoint is shown in Table 1. The variables selected for optimization, i.e., incubation temperature, incubation time, and the concentration of inducer, were coded as X 1, X 2, and X 3, respectively.
The model constructed as a response function of the variables on cellulase production was a second-order polynomial as follows (Eq. 1):
Where, Y is the measured response (cellulase yield); β, β i , and β ii are the regression coefficients, and X 1–X 3 are the factors under study.
For three variable systems, the model equation is given below (Eq. 2).
Regression analysis and estimation of the coefficients were performed using Design Expert ® software (Statease Corp, USA). The contributions of individual parameters and their quadratic and interaction effects on cellulase production were determined.
Results and Discussion
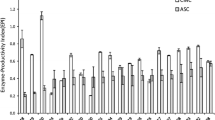
Based on results of the preliminary experiment, the levels of media components were set at the levels that gave maximal yield of cellulase to design the medium composition used in the present study. The levels of incubation time, temperature, and inducer concentration were varied as indicated in the experimental design. Statistical optimization by experimental design offers the opportunity to find out the optimal levels of process variables under any given condition by establishing the relationship between factors and the predicted responses. For response surface methodology based on the Box–Behnken design, which was used for optimization of cellulase production, 17 experimental runs with different combinations of three factors were carried out. For each run, the experimental responses along with the predicted response calculated from the regression equation (Eq. 3) are shown in Table 1. The maximum response (25.1 U/gds) was obtained in run number 17, and in general, all the runs with middle levels of parameters gave higher yields compared to other combinations. The data was analyzed by multiple regression analysis, and the regression coefficients were determined (Table 2).
A second-order polynomial equation (Eq. 3) was derived to represent the cellulase production as a function of the independent variables tested.
Where, Y is the predicted response, and X 1, X 2, and X 3 are coded values of incubation temperature, incubation time, and inducer concentration, respectively. The regression equation was used to calculate the predicted responses given in Table 1. A comparison of the predicted values with the experimentally obtained values indicated that these data are in reasonable agreement (Fig. 1).

Comparison of experimental and predicted results of cellulase production in 17 experimental runs
Adequacy of the model was tested by the Fisher’s statistical test for the analysis of variance (ANOVA) using Design Expert software, and the results are shown in Table 3. ANOVA of the quadratic regression model suggests that the model is significant with a computed F value of 4.69 and a P > F value lower than 0.05. A lower value for the coefficient of variation suggests higher reliability of the experiment, and in this case, the obtained CV value of 7.48% demonstrates a greater reliability of the trials. The R 2 value obtained was 0.8579, indicating that 85.79% of the sample variation is attributed to the factors and only 14.21% can occur due to chance. Table 3 also gives the P values of each of the parameters and their quadratic and interaction terms. The significance of individual variables can be evaluated from their P values, the more significant terms having a lower P value. The values of P > F less than 0.05 indicates that the model terms are significant and this case X 1, X 3, \(X_1^2 \), and \(X_2^2 \) were found to be significant model terms. There were no significant interactions between the parameters.
Response surface curves were plotted to understand the interaction effects of variables and for identifying the optimal levels of each parameter for attaining maximal cellulase yield. Figure 2a–c represents the response surfaces obtained for the interaction effects of tested variables. The data presented in the response plots indicated that the cellulase production increased with an increase in the concentration of crude inducer, and the maximum production was obtained at the maximal concentration of inducer (0.333 ml/gds), indicating that the inducer had the highest effect on cellulase production.

a Three dimensional response surface plot for interaction of incubation temperature and incubation time. b Three dimensional response surface plot for interaction of incubation temperature and inducer. c Three dimensional response surface plot for interaction of inducer and incubation time
The shapes of contour plots indicate the nature and extent of the interactions. It is clearly observed from the response surface and contour plot that there are no significant interactions between the tested variables.
Regardless of the incubation temperature, the maximal cellulase yield was obtained at or near the middle level of incubation time (64–67 h), and variations in incubation time did not affect the temperature optima between 32 and 33 °C, confirming the lack of interaction between these parameters (Fig. 2a). The optimal temperature and initial moisture content were within these ranges where the maximal activity of 23.5 U/gds was predicted by the model.
The effects of incubation temperature and concentration of inducer is shown in Fig. 2b. It may be observed from the response surface plot that with the increase in concentration of inducer, there is a minor shift in optimal temperature toward the lower values, though this is not a very significant interaction effect. The optimal combinations of incubation temperature and inducer concentration can be deduced to be 32–33 °C and 0.3–0.33 ml/gds, respectively, when the predicted yield was 24.1 U/gds.
Similarly, the interaction effects plotted for inducer concentration and incubation time showed that there are no significant interactions between these variables that affect cellulase production (Fig. 2c). However, this confirmed that optimal time range lied between 64 and 67 hours, and the optimal inducer concentration was between 0.3 and 0.333 ml/gds.
Discussion
The cost of cellulase enzyme is a major factor in the process economics of biomass to ethanol technology [14]. The production cost of cellulases can be brought down if cheaper substrates are used, as the cost of substrate account for a major fraction of the production cost [3]. Considering these, the present study was undertaken to explore the potential of using sugar cane bagasses—a cheap substrate—for cellulase production by the filamentous fungus T. reesei RUT C30. As “solid-state fermentation” can be effectively utilized for the production of cellulases from lignocellulosic biomass, this technology was employed in the study. SSF technology needs less space per unit biomass used as substrate and is less demanding on infrastructure and running costs [15, 16]. Environmental factors such as temperature, pH, water activity, oxygen levels, and concentrations of nutrients and products in the medium can significantly affect microbial growth and product formation under SSF [16], and a judicious selection of the levels of these parameters can dramatically improve the yield of enzymes under SSF. T. reesei RUT C30 produced 5.11 U/gds of cellulase under SSF on sugar cane bagasse in the basal medium. We expected to increase the cellulase yield from T reesei using sugar cane bagasse substrate by the optimization of process parameters, and the study was undertaken to evaluate the effects of process variables and optimize their levels for improving production of the enzyme.
Based on the results obtained from the SSF experiments on sugarcane bagasse, most significant influence was exhibited by parameters such as incubation temperature and inducer concentration, and no parameter interactions were found to have any significant effects on cellulase production. In this study, temperature had the second largest influence on production of cellulase. Though T reesei have been cultivated largely at 30 °C for production of the enzyme [17, 18], in the current study, a temperature range of 32–33 °C was found to be optimal for cellulase production. Though this is different from the reported optimal values for incubation temperature including our own previous report on SSF production using wheat bran as substrate [10], it has been reported that T reesei RUT C30 is capable of growth at a wide range of temperatures (17–37 °C) and is able to secrete enzymes [19]. Increase in protein secretion was observed for the T reesei strains QM6a and RL-P37 at 37 °C [20, 21]. Though these results were pertaining to xylanase production by the fungus, it has also been observed by the authors that endoglucanase production by the fungus increased to almost threefold upon cultivation at 37 °C. There could be differences in the synthesis and/or secretion of proteins in response to incubation temperature and/or carbon source, and the optimal incubation temperature may be different with different substrates. The relevance of these findings in the current context has to be evaluated for a conclusive statement on the positive effect of increased incubation temperature. Nevertheless, repeated trials on cellulase production at the optimal temperature range demonstrated the reliability of the data.
The parameter that showed the highest influence on cellulase production by T. reesei in the optimization studies was the concentration of inducer in the medium. It is well known that the cellulase system of T reesei is inducible, and cellobiose is a potent inducer at lower concentrations [22–24]. Generation of crude inducers making use of the tran-glucosidase activity of beta glucosidases is not a new concept. Allen and Mortensen [25] had used the purified beta-glucosidase from Aspergillus pheonicis for generating crude cellulase inducing preparation from glucose syrup. Cellobiose is also a known inducer of cellulase below a certain threshold level and can be generated by the action of T reesei cellulases on cellulose. We used this technology for generating a crude mixture of inducer, which was added in the fermentation medium to improve cellulase induction. The results demonstrated that the addition of crude inducer was highly effective in improving cellulase production. Previous studies in our lab had demonstrated that cellobiose with other uncharacterized compounds are generated by such a treatment and might be aiding to induce the cellulase genes of T reesei (unpublished results).
Under the optimal conditions of temperature and inducer concentration, the ideal incubation time was found to be 66 h, which is advantageous because most of the cellulase fermentations needed longer duration to obtain maximal activities.
Parameter interactions did not play any major role in cellulase production as evidenced from the results. The optimization studies performed on process variables affecting cellulase production under SSF by T reesei RUT C 30 on sugarcane bagasse was effective in enhancing the production of enzyme from 5.11 to 24.15 FPAse U/gds a 4.7-fold increase in production. Several authors have reported cellulase production using T reesei with a wide variety of carbon sources both under SmF and SSF (see Sukumaran et al. [26] for a review), and the yield of total cellulase activity ranges from 0.3 to ~7–8 U/ml in the case of SmF or from 10–15 to ~430 U/g cellulose. However, in majority of the cases, the activity units are expressed differently. In some reports, the units are expressed as units per gram dry substrate (U/gds) where, as in others, it is units per gram of cellulose. The optimized yield obtained in this study is a reasonably good production on an untreated natural substrate like sugar cane bagasses, and further improvements in production are envisaged with a pretreated substrate and with the fine tuning of other parameters.
Conclusions
Optimization of environmental parameters and media for fermentation is a significant concern in developing a suitable bioprocess for cellulase production. The present study addresses this issue and tries to provide a robust solution in developing a suitable SSF process with the cheap biomass resource “sugarcane bagasse” as substrate. Response surface Box–Behnken optimization of the significant process variables resulted in an almost fivefold increase in cellulase production. Enhancing cellulase production in SSF on cheap substrates can go a long way in bringing down the cost of cellulases, which will eventually help to develop an economical process for bio-fuel production.
References
Lynd, L. R., Wyman, C. E., & Gerngross, T. U. (1999). Biocommodity engineering. Biotechnology Progress, 15, 777–793.
Reith, J. H., den Uil, H., van Veen, H., de Laat WTAM, Niessen, J. J., de Jong, E., et al. (2002). Co-production of bioethanol, electricity and heat from biomass residues. 12th European Conference and Technology Exhibition on Biomass from Energy, Industry and Climate Protection, Amsterdam, The Netherlands, 17–21 June.
Wen, Z., Liao, W., & Chen, S. (2005). Production of cellulase/b-glucosidase by the mixed fungi culture Trichoderma reesei and Aspergillus phoenicis on dairy manure. Process Biochemistry, 40, 3087–3094.
Chahal, D. S. (1985). Solid-state fermentation with Trichoderma reesei for cellulase production. Applied Environmental Microbiology, 56, 554–557.
Pandey, A., Soccol, C. R., Nigam, P., & Soccol, V. T. (2000). Biotechnological potential of agro-industrial residues I: sugarcane bagasse. Bioresource Technology, 74, 69–80.
Du Toit, P. J., Olivier, S. P., & van Bijon, P. L. (1984). Sugarcane bagasse as a possible source of fermentable carbohydrates. I. Characterization of bagasse with regard to monosaccharide, hemicellulose, and animoacid composition. Biotechnology and Bioengineering, 26, 1071–1078.
Aiello, C., Ferrer, A., & Ledesma, A. (1996). Effect of alkaline treatments at various temperatures on cellulase and biomass production using submerged sugarcane bagasse fermentation with Trichoderma reesei QM9414. Bioresource Technology, 57, 13–18.
Gutierrez-Correa, M., & Tengerdy, R. P. (1997). Production of cellulase on sugar cane bagasse by fungal mixed culture solid substrate fermentation. Biotechnology Letters, 19(7), 665–667.
De-Paula, E. H., Ramos, L. P., & Azevedo, M. O. (1999). The potential of Humicola grisea var thermoida for bioconversion of sugar cane bagasse. Bioresource Technology, 68, 35–41.
Singhania, R. R., Sukumaran, R. K., & Pandey, A. (2007). Improved cellulase production by T reesei RUT C30 under SSF through process optimization. Applied Biochemistry and Biotechnology, 142(1), 60–70.
Herbert, D., Phipps, P. J., & Strange, P. E. (1971). Chemical analysis of microbial cells. Methods in Microbiology, 5B, 249–344.
Ghose, T. K. (1987). Measurement of cellulase activities. Pure & Applied Chemistry, 59, 257–268.
Box, G. E. P., & Behnken, D. W. (1960). Some new three level designs for the study of quantitative variables. Technometrics, 2, 455–475.
Von Sivers, M., & Zacci, G. (1995). A techno-economical comparison of three processes for the production of ethanol. Bioresource Technology, 51, 43–52.
Raimbault, M. (1998). General and microbiological aspects of solid substrate fermentation. Electronic Journal of Biotechnology [online], 1(3). Retrieved August 28, 2001 from http://www.ejbiotechnology.info/content/vol1/ issue3/full/9/9.PDF. ISSN 0717-3458.
Pandey, A., Selvakumar, P., Soccol, C. R., & Nigam, P. (1999). Solid-state fermentation for the production of industrial enzymes. Current Science, 77, 149–152.
Reczey, K., Szengyel, Z. S., Eklund, R., & Zacchi, G. (1996). Cellulase production by T. reesei. Bioresource Technology, 57, 25–30.
Gutierrez-Correa, M., Portala, L., Moreno, P., & Tengerdy, R. P. (1999). Mixed culture solid substrate fermentation of Trichoderma reesei with Aspergillus niger on sugar cane bagasse. Bioresource Technology, 68, 173–178.
Haltrich, D., Nidetzky, B., Kulbe, K. D., Steiner, W., & Zupancic, S. (1996). Production of fungal xylanases. Bioresource Technology, 58, 137–161.
Suh, D. H., Becker, T. C., Sands, J. A., & Montenecourt, B. S. (1988). Effects of temperature on xylanase secretion by Trichoderma reesei. Biotechnology Bioengineering, 32, 821–825.
Merivuori, H., Tornkvist, M., & Sands, J. (1990). Different temperature profiles of enzyme secretion by two common strains of Trichoderma reesei. Biotechnology Letters, 12, 117–120.
Mandels, M., & Reese, E. T. (1960). Induction of cellulase in fungi by cellobiose. Journal Bacteriology, 79(6), 816–826.
Kubicek, C. P., & Penttila, M. E. (1998). Regulation of production of plant, polysaccharide degrading enzymes by Trichoderma. In E. Harman, C. P. (Eds.), Trichoderma and Gliocladium, vol 2G (pp 49–72). London: Taylor & Francis.
Lynd, L. R., Weimer, P. J., van Zyl, W. H., & Pretorious, I. S. (2002). Microbial cellulase utilization: Fundamentals and biotechnology. Microbiology Molecular Biology Review, 66, 506–577.
Allen, U. A. L., & Mortensen, R. E. (1981). Production of cellulase from Trichoderma reesei in fed-batch fermentation from soluble carbon sources. Biotechnology Bioengineering, 23, 2641–2645.
Sukumaran, R. K., Singhania, R. R., & Pandey, A. (2005). Microbial cellulases—Production, applications and challenges. Journal of Scientific and Industrial Research, 64, 832–844.
Acknowledgments
Authors are thankful to the Council of Scientific and Industrial Research, Govt. of India for the research grant on project CMM013 which funded this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mekala, N.K., Singhania, R.R., Sukumaran, R.K. et al. Cellulase Production Under Solid-State Fermentation by Trichoderma reesei RUT C30: Statistical Optimization of Process Parameters. Appl Biochem Biotechnol 151, 122–131 (2008). https://doi.org/10.1007/s12010-008-8156-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-008-8156-9