
Abstract
Wireworms, the larval stage of click beetles (Coleoptera: Elateridae), continue to be one of the major concerns of cereal producers, primarily due to the lack of effective pesticides and species-specific management options. To have a better understanding of species-specific interactions of one of the most damaging wireworms in the Pacific Northwest and intermountain regions of the USA, a greenhouse study was set to evaluate the damage from the sugar beet wireworm Limonius californicus to wheat and barley planted at different depths and in soil media with varying levels of organic content and texture. Overall, the evaluated wheat appeared to be more susceptible than the barley, showing greater reductions in emergence success and foliar biomass. The greatest loss of foliar biomass was observed in peatmoss-dominated medium, as indicated by a significant host plant-by-soil media interaction. Percentage of plants fed upon by L. californicus was significantly higher in the sand-dominated medium than peatmoss-dominated and 1:1 mix media. Moreover, manipulation of soil media by the addition of diatomaceous earth showed no consistent effect in protecting the planted wheat. Our findings indicated that in addition to quantifying wireworm species-specific interactions, host plant interactions with the environment in the presence of wireworm infestation should also be further studied. These relationships could influence the outcome of integrated management approaches and future risk assessment models and recovery plans.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wireworms, the larval stage of click beetles (Coleoptera: Elateridae), are soil-dwelling organisms that have reemerged as a significant threat to a wide range of agricultural crops in recent years. Wireworms represent one of the major current concerns for cereal production in the Pacific Northwest (PNW) and intermountain regions of the USA. In cereals, while wireworm feeding on sprouting seed can result in failed emergence, young seedlings may survive and root and stem damage be observed. The typical damage in cereals is characterized as dead central leaves, also referred to as ‘dead heart,’ which would eventually result in seedling death. Wireworm-affected cereal fields may be spotted by the presence of large patches of missing and weakened plants.
It is widely thought that as the residual effects of the environmentally persistent conventional insecticides, which are now banned due to environmental and human health concerns, started to fade away, wireworm pressure has resurged (Vernon et al. 2008). The only substitute class of pesticides registered for application in cereals (i.e., neonicotinoids) does not induce considerable mortality, providing only limited protection against wireworms (Vernon et al. 2008, 2009). In a recent study, however, application of entomopathogenic fungi provided effective control against wireworms (Reddy et al. 2014). As management options in cereals have remained limited for more than a decade, a detailed understanding of wireworm ecology has become crucial in developing integrated management approaches to minimize their negative impact on cereal cropping systems.
Despite their limited mobility, wireworms are able to use environmental cues to orient themselves within their subterranean habitat. The vertical movement of wireworms in the soil profile is primarily in response to changes in ambient temperature and soil moisture (Lange et al. 1949; Jones 1951; Lafrance 1968; Lefko et al. 1998; Barsics et al. 2013). Variation in soil properties can change both water holding capacity and wetting patterns, and thus, soil characteristics may considerably influence wireworm ecological responses within its dynamic environment. For instance, soil porosity and compaction may not only influence fluctuations in moisture and temperature, but can also influence the ability of wireworms to relocate and/or survive (Jones and Shirck 1942; Parker and Seeney 1997; van Herk and Vernon 2006; Hermann et al. 2013). In relation to this, species-specific associations with variation in soil texture have been previously reported (Hawkins 1936). In addition to the abiotic factors, the presence of a host plant can also trigger directional movement of wireworms as they respond to carbon dioxide (CO2), and other volatile compounds (e.g. fatty acids), released from germinating seeds, roots, and stems (Gfeller et al. 2013; Barsics et al. 2013), which could perhaps drive species-specific preferences (Cherry and Stansly 2008).
As wireworms are continuously in contact with their subterranean habitat, it is also conceivable to assume that manipulations of soil, by adding particles damaging to their armored waxy cuticle could negatively impact their fitness. Diatomaceous earth (DE) is a natural silicon dioxide dust particle, obtained from fossilized unicellular diatom algae, with insecticidal potential (Korunic 1998). While insect mortality may result from desiccation, as sharp silica particles damage the waxy cuticle (Ebeling 1971), sorptive dusts are also known to act as repellents since the insect would avoid coming into contact with the substance (Korunic 1998; Mohan and Fields 2002). Due to the non-toxic nature of the dust, and lack of harmful environmental residue, DE has been employed to manage a range of home and storage pests such as mites (Collins 2006), weevils (Arthur and Throne 2003), cockroaches (Faulde et al. 2006), and termites (Ebeling and Wagner 1959). The effectiveness of this product as a repellent and/or an insecticide against wireworms has not been evaluated.
Despite decades of research on wireworm ecology and management, wireworms continue to be a formidable challenge to cereal production, in part, due to the lack of species-specific knowledge of ecology (Traugott et al. 2015a, b). Focusing on one of the most damaging species in the Pacific Northwest (Milosavljevic et al. 2015; Esser et al. 2015) and the Intermountain regions in the USA (Morales-Rodriguez et al. 2014; Rashed et al. 2015), the sugar beet wireworm Limonius californicus, the aim of the present study was twofold: (1) to compare the extent of wireworm damage when wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.) hosts were planted in different soil media and at varied seeding depths and (2) to evaluate the effectiveness of DE in protecting two widely planted wheat cultivars in Idaho.
Materials and methods
Study site and baiting wireworms
The study was conducted at the University of Idaho Aberdeen Research and Extension Center greenhouses in Aberdeen, ID, between May and August of 2015. Daily average temperature (standard deviation) was recorded at 26.8 °C (2.47). Wireworm L. californicus was collected from traps placed in a dryland wheat field in eastern Idaho, located east of Ririe, ID (43°34′56.5″N 111°32′21.5″W). All wireworms were baited between May and July, by a mixture of germinating wheat and barley placed at an approximately 15 cm (6 in.) depth (i.e., solar bait trap). Collected insects were maintained in 5 × 5 × 10-cm (W × L × H) plexiglass containers filled with field soil and a 1:1 mix of eight to ten wheat and barley seeds. Wireworms were used in the experiment within 10–14 days of collection.
Soil media preparation and characterizations
The three soil media were prepared by manipulating sand-to-peatmoss (Sun Gro Horticulture Canada Ltd., Seba Beach, AB, Canada) ratios in the experimental pots. In addition to the peatmoss and sand component, the soil media mix contained fixed amounts of fertilizer (Osmocote; Scott-Sierra Horticultural Products Co., Marysville, OH, USA) and vermiculite (Therm-o-Rock West INC., Chandler, AZ, USA). The sand-dominated medium consisted of 75% sand and 25% peatmoss, the peatmoss-dominated medium, rich in organic matter, contained 75% peatmoss and 25% sand. The 1:1 mix medium contained equal amounts of both peatmoss and sand contents. Soil media was homogenized prior to study initiation. Sand, peatmoss, and the 1:1 mix were analyzed for chemical properties where subsamples from individual pots (5/soil medium) were collected prior to the initiation of the study, dried at 40 °C prior to chemical analyses, and ground to pass through a 2-mm sieve. Soil media electrical conductivity and pH were determined potentiometrically using a 1:2 (w:v) soil-to-water ratio. Total carbon (TC) and total nitrogen (TN) were determined via high-temperature combustion on a VarioMax CN analyzer (Elementar Americas Inc., Mt. Laurel, NJ). The soil media C:N ratio was calculated based on the measured TC and TN concentrations.
Wireworm damage to wheat and barley in different soil media
The hard red spring wheat variety Klasic released by Northrup-King Company (Minneapolis, MN; now Syngenta, Golden Valley, MN) and the spring barley variety Idagold, released by Coors Brewing Company (Golden, CO, USA) were used throughout the experiment. Every square-shaped pot (35.5 × 35.5 × 24 cm; W × L × H) was planted with two wheat and two barley seeds, at 2.54 and 7.62 cm (1 and 3 in.) of seeding depth. Seeds were soaked for 6 h prior to planting and positioned at each corner of the pot, approximately three inches from the two closest edges. Immediately following seeding, a single wireworm of a late instar (1.9–2.5 cm in length) was placed at the 15.2 cm (6 in.) depth, equally distanced from each of the planted wheat and barley seeds, in the center of each pot.
The experiment was repeated three times, hereafter referred to as ‘time-blocks.’ Within every time-block, there were 9 experimental replicates (plus one control pot) per soil treatment (total 30 pots/time-block) arranged in a completely randomized design.
Parameters evaluated in this experiment included emergence success, percent of damaged plants (characterized by the presence of the discolored point of feeding; Fig. 1), and/or seedling death. In the case of dead seedlings, wireworm damage was confirmed by observing the point of feeding at the base of the stem just below soil surface. Root damage was not quantified but visual inspection indicated minimal detectable damage throughout our greenhouse experiment. Thus, for the sake of consistency, feeding damage is referred to the observed point of feeding on the stem tissue of an affected (wilting or exhibiting the ‘dead heart’ symptom) and/or dead seedling. Aboveground tissue was removed 5 weeks after emergence and dried at 40 °C until a constant mass was achieved for foliar biomass determination. Loss of biomass was calculated by subtracting biomass of the non-infested control plants from those infested with L. californicus.

Sugar beet wireworm Limonius californicus feeding on a wheat plant in the sand-dominated soil medium
Diatomaceous earth (DE) and managing wireworm damage
A second experiment was set up to evaluate the effectiveness of DE (Perma-Guard, Bountiful, UT, USA) in protecting planted wheat against wireworms. Using 3.8-l cylindrical pots, DE was applied at the rate 1.56 g/pot (168 kg/ha) and incorporated into the top three inches of soil, to mimic a surface application followed by shallow tillage in the field.
Each pot was planted with a single seed of the spring cultivars UI Stone (Chen et al. 2013) and UI Platinum (Chen et al. 2016). Seeds were soaked in water for 6 h prior to planting and were planted four inches apart at a 2.54 cm (1 in.) depth (shallow seeding). Immediately after planting, a single L. californicus was placed in 15 cm depth in the middle of every pot. There were two time-blocks. The first time-block consisted of 10 pots treated with DE and 10 non-treated control pots. There were two non-infested controls, one for each of the two treatments. The second time-block of the study included 8 plants for each of the two treatments (DE-treated and non-treated) and two non-infested control pots. Plants were removed 3 weeks after emergence. Plant emergence success, percentage of damaged plants, and foliar biomass were measured.
Statistical analysis
All statistical analyses were conducted in IBM-SPSS (ver. 22). Electrical conductivity, soil media pH, total carbon and nitrogen, and C:N ratios were compared using analysis of variance (ANOVA). Analysis of variance with binomial error was used to compare wheat and barley emergence success and percent damaged seedlings in the presence of wireworms. Controls were not included in emergence success analysis. Thus, descriptive comparisons between the emergence success of controls and experimental pots were also presented. Time-block, pot, host plant, soil medium, seeding depth, soil medium-by-host plant, soil medium-by-seeding depth, and seeding depth-by-host plant interactions were factors included in the initial model. Time-block, pot, and interaction terms were removed in a stepwise approach, if their effect was non-significant (P > 0.05). ANOVA was also used to compare foliar biomass loss in response to wireworm presence among different soil media treatments. Analysis of variance with binomial errors was also used to compare emergence success and the percentage of damaged seedlings between DE-treated and non-treated pots, and between the two wheat varieties. Similar to the previous experiment, the loss of biomass was compared using ANOVA. Time-block, pot, treatment (DE-treated vs non-treated), variety, and treatment-by-variety interaction were included in the starting model. A stepwise approach was used to remove time-block, pot, and the interaction term from the final model, where no effect was detected (P > 0.05).
Results
Soil media characterization
The evaluated parameters varied significantly among the three experimental soil media (Pillai’s Trace = 1.59, F 10,14 = 5.46, P = 0.002), with levels of electrical conductivity (F 2,10 = 15.27, P = 0.001; Fig. 2a), pH (F 2,10 = 22.42, P < 0.001; Fig. 2b), TN (F 2,10 = 47.15, P < 0.001; Fig. 2c), TC (F 2,10 = 87.80, P < 0.001 Fig. 2d), and C:N ratio (F 2,10 = 6.26, P = 0.017; Fig. 2e) all showing detectable variability. Electrical conductivity, TC, and TN levels were significantly higher in peatmoss-dominated medium than the sand-dominated and 1:1 mix media; the reverse pattern was observed for pH (Fig. 2b). The C:N ratio was significantly greater in peatmoss-dominated medium than the sand-dominated medium.

Soil characterization of the three experimental media; a electrical conductivity, b hydrogen concentration (pH), c total carbon, d total nitrogen, and e C:N ratio. Error bars represent standard errors (+/−1). Different letters indicate statistical differences at α = 0.05 (Tukey HSD)
Wireworm damage to wheat and barley in different soil media
Overall, the emergence success was significantly lower in wheat than barley in the presence of wireworm (Wald \(X_{1}^{2} = 26.55\), P < 0.001; Fig. 3). In non-infested controls, the majority of treatment combinations exhibited 100% successful emergence; the only exceptions were wheat planted at a 7.62 cm (3 in.) depth, in 1:1 media mix, and barley planted at a 7.62 cm depth, in the sand-dominated media, which had 60 and 80% emergence, respectively. Overall, emergence rate of pots infested with L. californicus ranged between 50 and 70% for wheat and between 85 and 95% for barley. Emergence success was not influenced by soil media (Wald \(X_{2}^{2} = 9.19\), P = 0.632; Fig. 3) or seeding depth (Wald \(X_{1}^{2} = 1.76\), P = 0.185; Fig. 2). A significant effect of time-block was detected (Wald \(X_{2}^{2} = 8.82\), P = 0.012). All interaction terms were removed from the final model due to their non-significant effects (P > 0.250).

Emergence success of wheat and barley hosts in pots infested with a single L. californicus. A significant difference in emergence success was detected between wheat and barley (P < 0.05). Seeding depth or soil media did not influence probability of successful emergence
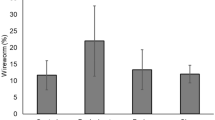
The point of feeding was mostly observed at the base of the stem just beneath the soil surface. The percentage of seedlings exhibiting feeding damage on the stem tissue was not statistically different between wheat and barley (Wald \(X_{1}^{2} = 1.31\), P = 0.252; Fig. 4) and was also not influenced by seeding depth (Wald \(X_{1}^{2} = 0.162\), P = 0.678; Fig. 4). The percentage of damaged seedlings was significantly affected by soil media, with sand-dominated medium being associated with the highest percentage of damaged plants (Wald \(X_{2}^{2} = 13.02\), P = 0.001; Fig. 4). No significant effects of pot or interaction terms were detected (all Ps > 0.05); therefore, all these factors were excluded from the final model.

Percentage of wheat and barley plants fed upon by Limonius californicus. There was a significantly higher rate of feeding damage in sand-dominated medium than the peatmoss-dominated or the 1:1 mix. Different letters indicate statistical differences at α = 0.05 (Tukey HSD)
Analysis of variance revealed that the loss in foliar biomass was predicted by host plant (F 1,205 = 9.25, P = 0.003), as it was significantly higher in the wireworm-affected wheat than barley seedlings. The extent of biomass loss to wireworm damage was also impacted by soil media (F 2,205 = 3.58, P = 0.030). However, variations in foliar biomass loss in response to soil media were primarily influenced by variations in wheat and not barley, as reflected in a significant host plant-by-soil media interaction (F 2,205 = 4.44, P = 0.013; Fig. 5).

Loss of foliar biomass in wheat and barley plants in pots infested with wireworms. While the loss of foliar biomass was overall greater in wheat then barley (marked with the asterisk), this effect was primarily driven by the loss of biomass in wheat planted in the peatmoss-dominated medium. Error bars represent confidence intervals (95% CI)
Diatomaceous earth and managing wireworm damage
Emergence success of both UI Platinum and UI Stone wheat varieties did not significantly differ (Wald \(X_{1}^{2} = 0.00\), P = 1.00), and was not influenced by diatomaceous earth application (Wald \(X_{1}^{2} = 0.703\), P = 0.455). There was 100% emergence success in both DE-treated and non-treated controls not infested with wireworms. There were no effects of time-block, pot, and variety-by-treatment detected; thus, these variables were not included in the final model.
The percentage of damaged seedlings was also not affected by wheat variety (Wald \(X_{1}^{2} = 3.05\), P = 0.08) or DE application (Wald \(X_{1}^{2} = 1.25\), P = 0.262) (Fig. 6). Likewise, the overall loss of foliar biomass was not impacted by variety (F 1,47 = 0.913, P = 0.344) or DE treatment (F 1,47 = 0.955, P = 0.333), and differences detected between DE-treated and non-treated substrates appeared to be rather random. Significant effects of time-block (F 2,47 = 8.13, P = 0.006) and treatment-by-variety interaction (F 1,47 = 11.75, P = 0.001) were detected, and results for each of the two time-blocks were presented, separately (Fig. 7).

Percentage of plants affected by wireworms (characterized by the presence of the point of feeding) in the wheat varieties UI Stone and UI Platinum. No significant effect of variety or diatomaceous earth treatment was detected

Loss of foliar biomass in UI Stone and UI Platinum across the two time-blocks. Asterisks indicate significant differences between DE-treated and non-treated pots. Error bars represent confidence intervals (95% CI)
Discussion
Due to the wireworm subterranean habitat, they are constantly in contact with soil substrate; thus, it is conceivable that their ability to cause damage may be influenced by variations in physical properties of their immediate surroundings. Focusing on L. californicus, a key damaging species in the PNW and the intermountain regions, we quantified wireworm impact on different host plant genotypes in soil media of varying ratios of organic matter, as well as in the presence of potentially efficacious/repellent DE dust incorporated into soil media.
Wireworms affected both wheat and barley hosts. Overall, the wheat appeared to be relatively more susceptible to wireworms when compared to the barley. This higher susceptibility level was reflected in relatively lower rate of emergence and significantly greater loss of aboveground biomass in wheat than barley. It is also important to note, however, that since there was only one variety of wheat and barley included in our study, generalizing the observed relative tolerance of barley compared to wheat to other varieties would not be plausible, even though field observations also support this contention (A. Esser and D. Crowder, personal communication).
Despite decreased seedling emergence in wheat, the percentage of seedlings exhibiting feeding damage at the base of the stem (visible point of feeding) was not statistically different between the wheat and barley hosts (Fig. 4). However, this percentage was greatly influenced by soil media, being primarily associated with sand-dominated medium. This observation supported previous landscape-scale analysis by Hermann et al. (2013) who detected an association between damage to potato tubers and sandy soil. There, the increase in wireworm feeding and damage was attributed to rapid changes in moisture in porous sand, which would cause wireworms to obtain moisture by burrowing into plant tissue more frequently (Hermann et al. 2013). Alternatively, or in addition, while it has been suggested that soils rich in organic matter may help protect plants from wireworm feeding damage, more recent work by Traugott et al. (2007, 2008) did not find substantial evidence of Agriotes spp. feeding on significant amounts of soil organic matter in native soils (Traugott et al. 2007, 2008, 2015a, b, review). In the present study, plants and wireworms were grown in a wide-ranging soil environmental gradient from very low organic matter (i.e., sand) to very high organic matter (i.e., peatmoss), as confirmed by TC levels (Fig. 2d). Thus, it is plausible under the current study conditions that the feeding ecology of the wireworm on organic matter varied, at least to some degree, due to gradation in organic matter levels in their growing environment, and that our results provided evidence of the preceding notion that organic matter could potentially act as a buffering mechanism to decrease wireworm feeding damage. However, further research to quantify the specific feeding ecology of the wireworm under organic matter gradients is needed to support this contention.
Although aboveground biomass loss was significantly higher in wheat than barley, and despite the greater feeding damage observed in sand-dominated medium (Fig. 4), the difference in plant biomass was primarily driven by the greater biomass loss in wheat seedlings planted in peatmoss-dominated medium, a trend which was not observed for barley. The peatmoss-dominated medium contains significantly higher levels of TC, TN, EC, and lower pH levels when compared to sand-dominated and 1:1 mix media. This indicates that host plant interactions with soil substrate can have significant implications when developing management strategies, as the host plant ability to withstand wireworm damage may vary considerably across environments. The underlying mechanism for the increase in biomass loss in wheat in a medium rich in organic matter should be the subject of future studies.
In cereals, shallow seeding can promote quick emergence and vigorous stand growth where adequate soil moisture is present (Robertson et al. 2004; Cook and Veseth 1991; Kephart et al. 1989). Thus, it has been suggested that shallow seeding could potentially reduce losses to wireworms (Rashed et al. 2015). Here, however, no effect of seeding depth was detected in either soil media assay. Our failure to detect the effect of seeding depth on the extent of wireworm damage could be explained by the optimal greenhouse conditions, which supported healthy and vigorous plant growth, regardless of seeding depth. An increased seeding depth (>7.62 cm) may help to detect significant differences in wireworm damage.
Results from the first set of experiments revealed that the percentage of plants affected by wireworm damage was influenced by variations in soil media composition/texture, and also indicated that our greenhouse assays were sensitive enough to detect this variability. Subsequently, we examined the possibility of limiting wireworm damage/movement by manipulating substrate texture through incorporating non-chemical additives into the sand-dominated medium, which we showed to be associated with the highest percentage of damaged plants under our experimental conditions (Fig. 4).
Diatomaceous earth is a commercially available insecticide/repellent dust and has been gaining momentum as an efficacious replacement for environmentally toxic pesticides against storage and urban pests (e.g. Korunic 1998; Mewis and Ulrichs 2001; Arthur and Throne 2003). However, in our greenhouse experiments DE failed to protect seedlings of either wheat varieties against wireworms. As the mode of action of this product is primarily through damaging the protective cuticle and promoting insect desiccation, this lack of effectiveness could be attributed to relatively high moisture levels under greenhouse conditions, where pots were watered on regular basis. It has been documented that there is a reduction in the effectiveness of diatomaceous earth in the presence of higher moisture levels (Ebeling and Wagner, 1959; Ebeling 1971; see Korunic 1998, for a review). In addition, L. californicus wireworms used in the present study were all at their later stages of larval development possessing thick integuments. There is a possibility that DE could be more effective against larvae of early instars, a topic that needs further evaluations. The effectiveness of DE against wireworms under dryland field conditions and in response to various application rates is the subject of ongoing studies.
Interestingly, however, there was a consistent, although non-significant, trend where relatively fewer UI Stone seedlings exhibited feeding damage when compared to UI Platinum, varieties that represent different market classes of wheat. Although the difference was not statistically significant, it is worth evaluating further, especially since among-variety variations in susceptibility to wireworm damage have recently been reported (Higginbotham et al. 2014). Variations in volatiles and carbon dioxide levels produced from roots (Gfeller et al. 2013) and/or differences in plant development and underground physical attributes (Higginbotham et al. 2014) may play roles as predictors of the likelihood of wireworm feeding damage. Although differences in foliar biomass loss was detected between DE-treated and non-treated treatments were detected they did not appear to be consistent with our hypothesis that DE would provide some degree of protection for the planted seeds against wireworm damage (Fig. 7).
In summary, wheat seedlings were relatively more susceptible to L. californicus damage than barley. Although wireworms fed upon either host, barley was able to withstand the damage at a level, which was statistically detectable in our controlled experimental setup. Moreover, soil medium proved to be an important predictor of the percentage of damaged seedlings. More importantly, results revealed that host interaction with soil medium was an important variable influencing the outcome of wireworm–plant interaction, and should be taken into account when developing risk assessment models and management plans.
References
Arthur FH, Throne JE (2003) Efficacy of diatomaceous earth to control internal infestations of rice weevil and maize weevil (Coleoptera: Curculionidae). J Econ Entomol 96:510–518
Barsics F, Haubruge E, Verheggen FJ (2013) Wireworms’ management: an overview of the existing methods, with particular regards to Agriotes spp. (Coleoptera: Elateridae). Insects 4:117–152
Chen J, Wheeler J, Clayton J, Zhao W, O’Brien K, Zhang J, Jackson C, Marshall JM, Brown BD, Campbell K, Chen XM, Zemetra R, Souza EJ (2013) Registration of ‘UI Stone’ soft white spring wheat. J Plant Regist 7:321–326
Chen J, Wheeler J, O’Brien K, Zhao W, Klassen N, Zhang J, Bowman B, Wang Y, Jackson C, Marshall JM, Chen XM (2016) Release of ‘UI Platinum’ hard white spring wheat. J Plant Regist 10:36–40
Cherry R, Stansly P (2008) Abundance and spatial distribution of wireworms (Coleoptera: Elateridae) in Florida sugarcane fields on muck versus sandy soils. Fla Entomol 91:383–387
Collins DA (2006) A review of alternatives to organophosphorus compounds for the control of storage mites. J Stored Prod Res 42:395–426
Cook RJ, Veseth RJ (1991) Wheat health management. American Phytopathological Society Plant Health Management Series. APS Press, St. Paul
Ebeling W (1971) Sorptive dusts for pest control. Annu Rev Entomol 16:123–158
Ebeling W, Wagner RE (1959) Rapid desiccation of drywood termites with insert sorptive dusts and other substances. J Econ Entomol 52:190–207
Esser AD, Milosavljevic I, Crowder DW (2015) Effects of neonicotinoids and crop rotation for managing wireworms in wheat crops. J Econ Entomol 108:1786–1794
Faulde MK, Tisch M, Scharninghausen JJ (2006) Efficacy of modified diatomaceous earth on different cockroach species (Othoptera, Blattellidae) and silverfish (Thysanura, Lepismatidae). J Pest Sci 79:155–161
Gfeller A, Laloux M, Barsics F, Kati DE, Haubruge E, du Jardin P, Verheggen FJ, Lognay G, Wathelet JP, Fauconnier ML (2013) Characterization of volatile organic compounds emitted by barley (Hordeum vulgare L.) roots and their attractiveness to wireworms. J Chem Ecol 39:1129–1139
Hawkins JH (1936) Relation of soil utilization to wireworm injury. J Econ Entomol 29:728–731
Hermann A, Brunner N, Hann P, Wrbka T, Kromp B (2013) Correlations between wireworm damages in potato fields and landscape structure at different scales. J Pest Sci 86:41–51
Higginbotham RW, Froese PS, Carter AH (2014) Tolerance of wheat (Poales: Poaceae) seedlings to wireworm (Coleoptera: Elateridae). J Econ Entomol 107:833–837
Jones EW (1951) Laboratory studies on the moisture relations of Limonius (Coleoptera: Elateridae). Ecology 32:284–293
Jones EW, Shirck FH (1942) The seasonal vertical distribution of wireworms in the soil in relation to their control in the Pacific Northwest. J Agric Res 56:125–142
Kephart KD, Stark JC, and Robertson L (1989) Irrigated spring wheat production guide for Southern Idaho. University of Idaho College of Agricultural and Life Sciences Extension Bulletin 697
Korunic Z (1998) Diatomaceous earth, a group of natural insecticides. J Stored Prod Res 34:87–97
Lafrance J (1968) The seasonal movements of wireworms (Coleoptera: Elateridae) in relation to soil moisture and temperature in organic soils of Southwestern Quebec. Can Entomol 100:801–807
Lange WH, Carlson EC, Leach LD (1949) Seed treatments for wireworm control with particular reference to the use of lindane. J Econ Entomol 42:942–955
Lefko SA, Pedigo LP, Batchelor WD, Rice ME (1998) Spatial modeling of preferred wireworm (Coleoptera: Elateridae) habitat. Environ Entomol 27:184–190
Mewis I, Ulrichs Ch (2001) Action of amorphous diatomaceous earth against different stages of the stored product pests Tribolium confusum, Tenebrio molitor, Sitophilus granarius and Plodia interpunctella. J Stored Prod Res 37:153–164
Milosavljevic I, Esser AD, Crowder DW (2015) Identifying wireworms in cereal crops. Washington State University Extension FS175E
Mohan S, Fields PG (2002) A simple technique to assess compounds that are repellent or attractive to stored-product insects. J Stored Prod Res 38:23–31
Morales-Rodriguez A, O’Neill RP, Wanner KW (2014) A survey of wireworm (Coleoptera: Elateridae) species infesting cereal crops in Montana. Pan Pac Entomol 90:116–125
Parker WE, Seeney FM (1997) An investigation into the use of multiple site characteristics to predict the presence and infestation level of wireworms (Agriotes spp., Coleoptera: Elateridae) in individual grass fields. Ann Appl Biol 130:409–425
Rashed A, Etzler F, Rogers CW, Marshall JM (2015) Wireworms in Idaho cereals: Monitoring and identification. University of Idaho Extension Bulletin 898
Reddy GV, Tangtrakulwanich K, Wu S, Miller JH, Ophus VL, Prewett J, Jaronski ST (2014) Evaluation of the effectiveness of the entomopathogens for the management of wireworms (Coleoptera: Elateridae) on spring wheat. J Invertebr Pathol 120:43–49
Robertson LD, Guy SO, Brown BD (2004) Southern Idaho dryland winter wheat production guide. University of Idaho. College of Agricultural and Life Sciences Extension Bulletin 827
Traugott M, Pazmandi C, Kaufmann R, Juen A (2007) Evaluating 15 N/14 N and 13C/12C isotope ratio analysis to investigate trophic relationships of elaterid larvae (Coleoptera: Elateridae). Soil Biol Biochem 39:1023–1030
Traugott M, Schallhart N, Kaufmann R, Juen A (2008) The feeding ecology of elaterid larvae in central European arable land: new perspectives based on naturally occurring stable isotopes. Soil Biol Biochem 40:342–349
Traugott M, Benefer CM, Blackshaw RP, van Herk WG, Vernon RS (2015a) Biology, ecology, and control of elaterid beetles in agricultural land. Annu Rev Entomol 60:313–334
Traugott M, Schallhart N, Staudacher K, Wallinger C (2015b) Understanding the ecology of wireworms and improving their control: a special issue. J Pest Sci 86:1–2
Van Herk WG, Vernon RS (2006) Effect of temperature and soil on the control of a wireworm, Agriotes obscurus L. (Coleoptera: Elateridae) by flooding. Crop Prot 25:1057–1061
Vernon RS, van Herk WG, Tolman J, Ortiz Saavedra H, Clodius M, Gage B (2008) Transitional sublethal and lethal effects of insecticides after dermal exposures to five economic species of wireworms (Coleoptera: Elateridae). J Econ Entomol 101:365–374
Vernon RS, van Herk WG, Clodius M, Harding C (2009) Wireworm management I: stand protection versus wireworm mortality with wheat seed treatments. J Econ Entomol 102:2126–2136
Acknowledgments
We would like to thank our field technician Fabiola Aguilar, our laboratory technician Scott Pristupa, and our laboratory assistant Cindrie Lowder for their help with different aspects of this experiment. Funding for this project was provided by the Idaho Wheat Commission, Idaho Barley Commission, USDA-ARS-REACCH-2011-68002-30191 and USDA-NIFA-Hatch-IDA01506.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Joseph Dickens.
Rights and permissions
About this article

Cite this article
Rashed, A., Rogers, C.W., Rashidi, M. et al. Sugar beet wireworm Limonius californicus damage to wheat and barley: evaluations of plant damage with respect to soil media, seeding depth, and diatomaceous earth application. Arthropod-Plant Interactions 11, 147–154 (2017). https://doi.org/10.1007/s11829-016-9474-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-016-9474-4