
Abstract
Cell cultures of Cayratia trifolia (Vitaceae), a tropical lianas, were maintained in Murashige and Skoog’s medium containing 0.25 mg l−1 NAA, 0.2 mg l−1 kinetin and casein hydrolysate 250 mg l−1. Cell suspension cultures of C. trifolia accumulate stilbenes (piceid, resveratrol, viniferin, ampelopsin), which on elicitation by any of 500 μM salicylic acid, 100 μM methyl jasmonate, 500 μM ethrel and 500 mg l−1 yeast extract, added on the 7th day, were enhanced by 3- to 6-fold (5–11 mg l−1) by the 15th day.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Polyphenolics such as stilbenes are associated with many health benefits that include antioxidant, anticancer, and anti-atherosclerosis properties (Waffo et al. 2008; Baur and Sinclair 2006; Delmas et al. 2006). Epidemiological studies have shown a link between stilbenes and reduced risk of cardiovascular disease because resveratrol inhibits the oxidation of low-density lipoprotein (LDL) and blocks platelet aggregation and eicosanoid synthesis (Renaud et al.1998; Waffo et al. 2008).
Plant secondary products are of immense use as potential drugs, nutraceuticals, and food additives. Due to their limited availability and complexity for chemical synthesis, an alternative route such as plant cell culture is gaining importance for large-scale production of desired compounds (Savitha et al. 2006). Enhancement of secondary metabolites by elicitation is one of the few strategies recently finding commercial application (Savitha et al. 2006). Phytoalexins from the Vitaceae constitute a rather restricted group of polyphenolic secondary metabolites belonging to the stilbenes family (piceid, resveratrol, viniferin, ampelopsin); their production is induced in plant cells in response to biotic stress. trans-resveratrol, as primary phytoalexin, can be transformed into several more potent antifungal stilbenes, such as ε-viniferin, a resveratrol dimmer (Soulard et al. 2006). Silbenes production is mainly studied in cell cultures of Vitis vinifera (Waffo et al. 2008). We have previously reported the occurrence of stilbenes in callus cultures of Cayratia trifolia (L.) Domin (Vitaceae), a wild relative of Vitis and tropical lianas occurring naturally in subtropical forests (Roat et al. 2008). In the present communication, we report the influence of elicitors on stilbenes accumulation in cell cultures of C. trifolia.
Materials and methods
Cell cultures
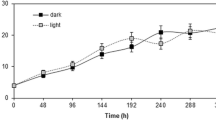
Cell suspension cultures were initiated from 4-week-old callus of Cayratia trifolia by transferring 2 g friable callus into a 250-ml Erlenmeyer flask containing 50 ml of liquid MS medium (Murashige and Skoog 1962) supplemented with 2,4-dichlorophenoxyacetic acid (2,4-D, 0.25 mg l−1) α-naphthaleneacetic acid (NAA, 0.2 mg l−1), kinetin (0.2 mg l−1), casein hydrolysate (250 mg l−1), and 3% w/v sucrose; pH was set to 5.8. The cultures were grown in the dark at 25°C on an orbital shaker at 100 rpm. Subsequently, 2,4-dichlorophenoxyacetic acid was withdrawn from the medium and cultures were maintained in MS medium supplemented with NAA (0.25 mg l−1) and kinetin (0.2 mg l−1). Subcultures were performed every 15 days by transferring 15 ml [125 mg dry mass (DM)] of the culture into 85 ml of fresh medium.
Cell cultures were treated with different concentrations of aqueous solution of yeast extract (YE, 100, 250, and 500 mg l−1), salicylic acid (SA, 100, 250, and 500 μM), ethrel (2-chloroethyl phosphonic acid, 100, 200, and 500 μM) and alcoholic methyl jasmonate (MeJa, 50, 100, and 250 μM) as shown in the "Results". The elicitors were prepared as stock solutions and added to the cultures after filter sterilization achieving the appropriate concentration on the 7th day of growth to induce polyphenol accumulation as described by Krisa et al. (1999).
Sample preparation
The cell cultures were harvested after 7 days of treatment (Krisa et al. 1999), washed with distilled water and filtered under mild vacuum. The cells were weighed to obtain the fresh weight per 100 ml medium (FW), and dry mass (DM) was then determined by drying the cells at 60°C in an oven to a constant weight. Dried homogenized cells (50 mg) were extracted in acetone-water 3:2 (v/v) for 12 h (room temperature) on a test tube rotator, centrifuged at 2,000 g for 15 min and then the supernatant was concentrated under vacuum at 40°C until the complete removal of acetone, the aqueous extract was partitioned twice with equal amount of ethyl acetate; finally, the ethyl acetate phase was concentrated under vacuum until dryness. This extract was then used for the HPLC analysis. All the extracts were redissolved in HPLC grade methanol and analyzed by using HPLC (pump L2130, auto sampler L-2200, FL detector L-2485; Merck-Hitachi). In brief, separation was accomplished on a (LichroCART)® 250 × 4 mm LiChrospher® (5 μm) RP-18 column protected by a guard column of the same material. The solvent system used was: solvent A-0.0025% trifluoroacetic acid in water; solvent B-80% acetonitrile (E. Merck, India) in solvent A. The mobile phase consisted of solvent (A) and (B). The step gradient programme of solvent B was as follows: 0–3 min: 14–18%; 3–12 min: 18–18%; 12–25 min: 18–22%; 25–30 min: 22–22%; 30–38 min: 22–40%; 38–43 min: 40–40%; 43–46 min: 40–60%; 46–48 min: 60–70%; 48–50 min: 70–70%; 50–52 min: 70–80%; 52–54 min: 80–80%; 54–56 min: 80–85%; 56–58 min: 85–100%; 58–60 min: 100–100%; 60–62 min: 100–14%; 62–65 min: 14–14%. Separation was performed at a flow rate of 1.0 ml min−1 and chromatographic peaks were monitored at λexc 300 nm and λem 390 nm (Roat et al. 2008). The spent medium was extracted with 100 ml ethyl acetate and analyzed by HPLC for stilbenes released in the medium.
Standard compound trans-resveratrol (3,5,4′-trihydroxystilbene) was purchased from Sigma Chemical (St. Louis, MO, U.S.A.) and trans-piceid (3,3′,4,5′-tetrahydroxystilbene), trans-ε-viniferin, and ampelopsin A were kindly provided by Prof J. M. Merillon, France. These were dissolved in methanol to yield a final concentration of 1.0 mg ml−1 and standard curve was prepared using resveratrol with concentrations ranging from 50–500 ng ml−1. The amount of the other compounds was calculated on the basis of trans-resveratrol. All results are averaged over two separate analyses for stilbenes estimation and two consecutive experiments with six replicate flasks in each treatment for growth value determination. The results are expressed as μg g−1 cell dry mass.
Results and discussion
Effect of SA
Maximum growth (8.9 DM g l−1) and stilbenes [resveratrol (68 μg g−1), viniferin (223 μg g−1), and ampelopsin (958 μg g−1)] accumulated in the cells treated with 500 μM of SA, while maximum piceid (174 μg g−1) accumulated in the cells treated with 250 μM SA. The maximum yield (11.4 mg l−1) of stilbenes was 6.2-fold higher in the cells treated with 500 μM of SA than that recorded in untreated cells and the yield of total stilbenes increased parallel to the concentration of SA (Table 1).
Effect of MeJa
Maximum growth (6.7 DM g l−1) and the yield of stilbenes [piceid (25 μg g−1) and ampelopsin (1340 μg g−1)] accumulated in the cells treated with 100 μM of MeJa, while maximum accumulation of resveratrol (186 μg g−1) was recorded in the cells treated with 250 μΜ. Maximum yield (11.1 mg l−1 DM) recorded was 6-fold higher than that in the control cultures and further increase in the concentration of MeJa resulted in decrease in stilbenes (Table 2). Viniferin accumulation remained unchanged in the cells treated with MeJa.
Effect of ethrel
Maximum growth (10 DM g l−1) was recorded in the cells treated with 100 μM of ethrel while maximum yield of stilbenes (7.3 mg l−1 DM) was recorded in the cells treated with 500 μM of ethrel [piceid (27 μg g−1), resveratrol (52 μg g−1), viniferin (309 μg g−1) and ampelopsin (418 μg g−1)]. Α parallel increase in ethrel concentration and the growth associated yield of stilbenes was observed in the cultures which resulted in 4-fold increase in stilbenes accumulation over the control (Table 3).
Effect of YE
Maximum growth (6.8 DM g l−1) and yield (5.0 mg l−1 DM) of stilbenes [resveratrol (81 μg g−1), viniferin (465 μg g−1) and ampelopsin (176 μg g−1)] accumulated in the cells treated with 500 mg l−1 of YE. This increase was 3-fold higher than that recorded in the control and lowest amongst the all elicitors used during the current work on C. trifolia (Table 4).
Like the intact plant, plant cell cultures are highly susceptible to external signals (Larronde et al. 2003). Fungal elicitors are known to induce defence related signals resulting in production of secondary metabolites, particularly production of polyphenols, i.e., phytoalexins. These signals are mediated through SA, ethylene and MeJa (Sudha and Ravishankar 2002). It was observed that a very low level of MeJa can increase stilbenes content in grapevine plants (Larronde et al. 2003), and addition of MeJa and fungal elicitor in the cell cultures of Vitis vinifera induced a 3 fold increase in stilbenes production (Soulard et al. 2006). In C. trifolia cell cultures, abiotic elicitors like SA and MeJa were more effective in enhancing stilbenes accumulation as compared to biotic elicitors like YE. YE also acts as a source of nitrogen, and this nutritional effect might have also played a role in growth and elicitation. Increased growth and phenylethanoid glycosides were also recorded in cell cultures of Cistanche deserticola (Cheng et al. 2005). We have also recorded a marked increase in isoflavonoids, another class of polyphenolics, in cells of Pueraria tuberosa treated with SA, MeJa, and ethrel (Goyal and Ramawat 2008a). Among these, ethrel was the best for isoflavonoid accumulation in short durations (Goyal and Ramawat 2008b). Thus, the elicitors stimulate the phenylalanine lyase, cinnamate 4-hydroxylase, coumarate CoA ligase pool of enzymes, and stilbene synthase, leading to higher production of stilbenes (Soulard et al. 2006). Ethrel is known to cause morphological changes and inhibit the development. Its action is based on ethylene production by its degradation on entering into plant cell. The resulting increased ethylene concentration inhibits cell extension or biomass production (Haque et al. 2007). In the present study, the cell culture treated with the aboitic elicitor ethrel showed increased growth and stilbenes production. Perhaps, due to higher growth, yield of stilbenes was lower than that recorded in the cells treated with SA and MeJa. The effect of various elicitors in decreasing order was SA→MeJa→ethrel→YE. These results clearly indicated a 6-fold increased stilbene production in 1-week treatment with abiotic elicitors in cell cultures of C. trifolia and are in accordance with earlier results with V. vinifera (Larronde et al. 2003). Effect of combination of elicitors in production system is being carried out.
References
Baur J, Sinclair DA (2006) Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Disc 5:493–506
Cheng XY, Guo B, Zhou HY, Ni W, Liu CZ (2005) Repeated elicitation enhances phenylethanpoid glycosides accumulation in cell suspension cultures of Cistanche deserticola. Biochem Eng J 24:203–207
Delmas D, Lancon A, Colin D, Jannin B, Latrufe N (2006) Resveratrol as a chemopreventive agent: a promising molecule for fighting cancer. Curr Drug Targets 7:423–442
Goyal S, Ramawat KG (2008a) Increased isoflavonoids accumulation in cell suspension culture of Pueraria tuberosa, by elicitors. Indian J Biotechnol 7:378–382
Goyal S, Ramawat KG (2008b) Ethrel treatment enhanced isoflavonoids accumulation in cell suspension cultures of Pueraria tuberosa, a woody legume. Acta Physiol Planta 30:849–853
Haque S, Farroqui AHA, Gupta MM, Sangwan RS, Khan A (2007) Effect of ethrel, chlormequat chloride and paclobutrazol on growth and pyrethrins accumulation in Chrysanthemum cinerariaefolium Vis. Plant Growth Regul 51:263–269
Krisa S, Larronde F, Budzinski H, Decendit A, Deffieux G, Merillon JM (1999) Stilbenes production by Vitis vinifera cell suspension cultures: methyl jasmonate induction and 13C biolabeling. J Nat Prod 62:1688–1690
Larronde F, Gaudillere JP, Krisa S, Decendit A, Deffieux G, Merillon JM (2003) Airborne methyl jasmonate induces stilbene accumulation in leaves and berries of grapevine plants. Am J Enol Vitic 54:63–66
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15:473–497
Renaud S, Guégen R, Schenker J, D’Houtaud A (1998) Alcohol and mortality in middle-aged men from eastern France. Epidemiology 9:184–188
Roat C, Kumar N, Ramawat KG (2008) Occurrence of stilbenes in unorganized cultures of Cayratia trifolia (L.) Domin. J Herb Med Toxicol 2:11–15
Savitha BC, Timmaraju R, Bhagyalaksami N, Ravishankar GA (2006) Different biotic and abiotic elicitors influence betalain production in hairy root cultures of Beta vulgaris in shake flask and bioreactor. Process Biochem 41:50–60
Soulard CS, Richard T, Merillon JM, Monti JP (2006) 13CNMR analysis of polyphenol biosynthesis in grape cells: impact of various inducing factors. Analytica Chimica Acta 563:137–144
Sudha G, Ravishankar GA (2002) Involvement and interaction of various signaling compounds on the plant metabolic events during defense response, resistance to stress factors, formation of secondary metabolites and their molecular aspects. Plant Cell Tiss Org Cult 71:181–212
Waffo P, Krisa S, Richard T, Merillon JM (2008) Grapevine stilbenes and their biological effects. In: Ramawat KG, Merillon JM (eds) Bioactive molecules and medicinal plants. Springer, Heidelberg, Germany, pp 24–55
Acknowledgments
This work was supported by financial assistance from UGC-DRS under special assistance programme for medicinal plant research and DST-FIST programme for infrastructure development to KGR. The authors wish to thank Prof J.M. Merillon, Faculty Pharmacy, University of Bordeaux-2, France, for the gift of authentic compounds and his help in the work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Roat, C., Ramawat, K.G. Elicitor-induced accumulation of stilbenes in cell suspension cultures of Cayratia trifolia (L.) Domin. Plant Biotechnol Rep 3, 135–138 (2009). https://doi.org/10.1007/s11816-009-0082-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11816-009-0082-y