
Abstract
In this paper, we would like to show unexpected morphogenic potential of cell suspensions derived from seedling explants of Gentiana kurroo (Royle). Suspension cultures were established with the use of embryogenic callus derived from seedling explants (root, hypocotyl and cotyledons). Proembryogenic mass proliferated in liquid MS medium supplemented with 0.5 mg l−1 2,4-D and 1.0 mg l−1 Kin. The highest growth coefficient was achieved for root derived cell suspensions. The microscopic analysis showed differences in aggregate structure depending on their size. To assess the embryogenic capability of the particular culture, 100 mg of cell aggregates was implanted on MS agar medium supplemented with Kin (0.0–2.0 mg l−1), GA3 (0.0–2.0 mg l−1) and AS (80.0 mg l−1). The highest number of somatic embryos was obtained for cotyledon-derived cell suspension on GA3-free medium, but the best morphological quality of embryos was observed in the presence of 0.5–1.0 mg l−1 Kin, 0.5 mg l−1 GA3 and 80.0 mg l−1 AS. The morphogenic competence of cultures also depended on the size of the aggregate fraction and was lower when size of aggregates decreased. Flow cytometry analysis reveled luck of uniformity of regenerants derived from hypocotyl suspension and 100% of uniformity for cotyledon suspension.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Gentiana kurroo Royle (Gentianaceae) is an important, native Indian species used for medicinal purposes. Because of possible extinction, it is legally protected by law and cannot be exported (Ministry of Commerce, Government of India; Raina et al. 2003).
Sharma et al. (1993) attempted to micropropagate this species from nodal segments. They obtained an average 3.5 shoots per explant. Liquid culture constitutes a more efficient method of plant propagation, but the fundamental disadvantage is the risk of hyperhydration (Preil 2005). Up to the present time, only Hosokawa et al. (1998) has described the propagation of G. scabra cv. WSP-3 in liquid culture with a very high plantlet yield via organogenesis. The process is most often induced for in vitro multiplication of the genus (Morgan et al. 1997; Yamada et al. 1991; Momčilović et al. 1997).
Suspension cultures derived from callus tissue, which had been initiated from seedling explants, were previously obtained only for G. pannonica (Mikuła et al. 2002), G. tibetica and G. cruciata (Mikuła et al. 1996). This type of culture appears very efficient and a long-term source of somatic embryos, and could be used to study embryo development (Mikuła et al. 2001). It is relatively easy to control the growth of liquid cultures to determine those predisposed for cell suspension for transformation experiments (Hosokawa et al. 2000) and protoplast isolation. Gentian suspension cultures with a lack of embryogenic capability can be used for secondary metabolites production (Kamińska et al. 1998).
The occurrence of unexpectedly high morphogenic potential of G. kurroo seedling explants expressed by abundant somatic embryo production is the reason for the enlargement of our in vitro experiments, including the establishment of cell suspension cultures. The aim of the paper is to describe the prolific embryogenic character of suspensions derived from callus of various seedling explants in the presence of selected plant growth regulators.
Materials and methods
Culture material
Three cell suspension cultures derived from cotyledon (K/C), hypocotyl (K/H) and root (K/R) were used. They were established from callus tissue initiated on G. kurroo seedling explants cultured on initiation medium (IM) consisting of MS (Murashige and Skoog 1962) agar medium supplemented with 0.5 mg l−1 2,4-D and 1.0 mg l−1 Kin. After 2 months in culture, 2.0 g of callus tissue of each of explants were transferred separately into 100-ml conical flasks with 10 ml of liquid IM medium. During the first 4 weeks, 10.0 ml of fresh medium were regularly added every 7 days. After that, cultures were transferred into 250-ml conical flasks with 80 ml of medium and subcultured at 1-week intervals. Cultures were maintained in diffuse light, in the culture chamber at 22 ± 1°C in photoperiod 16:8 day:night and a rotary shaker at 130 g.
Microscopic analysis
Established cell suspensions were sampled with the application of Sigma stainless steel sieves with different mesh size for aggregate selection of >300, 300–150 and <150 μm. To describe the morphogenic events leading to embryogenesis in cell suspensions, the samples of plant material were selected and observed microscopically. In vivo observations were carried out with help of light microscopy Vanox (Olympus) without staining.
Transmission electron microscope studies of suspension were performed on the different aggregates. Samples were fixed with 2.0% glutardehyde in 0.1 mol l−1 cacodylate buffer at pH 7.0 for 2 h and 2.0% OsO4 for 2 h at 4°C. After that, they were dehydrated using a graded series of ethanol and embedded in Spurr’s resin. Semi-thin (1–2 μm) and ultra-thin sections were cut with LKB (Sweden) ultramicrotome. Semi-thin sections were stained with 0.1% toluidine blue in 1.0% borax and observed under a light microscope. Ultra-thin sections were contrasted with uranyl acetate and lead citrate and observed in Tesla transmission electron microscope.
For scanning electron microscope investigation samples were also fixed with 2.0% glutardehyde in 0.1 mol l−1 cacodylate buffer at pH 7.0 for 2 h followed by 2.0% OsO4 for 24 h. They were dehydrated using a graded series of ethanol and ethanol with acetone and dried with liquid CO2 in a critical point dryer Type E 3100 JAMBO SERIES II (Polarno Equipment). After that samples were coated with gold using a Coater “Joel”. Finally, samples were scanned using a scanning electron microscope “Joel” model JSM-S1.
Curve of cell suspension growth
For growth curve analyses 1.0 g of 2-month-old suspension cultures K/C, K/H and K/R were resuspended in 30 ml of IM medium in 100-ml conical flask. Tissue growth was measured every 3 days from the day of subculture to day 24. To determine fresh mass, cells from one flask were collected on a filter paper, dryed under vacuum and weighed. Dry mass was measured after drying the sample at 70°C for 48 h. Each experiment was repeated three times with three samples per data point. The coefficient of the growth was defined as a ratio of fresh and dry weights of final to initial tissue weight.
Morphogenic capacities of cell suspensions
Three cell suspension cultures K/C, K/H and K/R were divided onto four fractions: >500, 500–300, 300–150 and <150 μm. From each fraction, 100 mg of tissue was plated into 9-cm-diameter Petri dishes containing of 50 ml of MS medium supplemented with Kin (0.0; 0.1; 0.5; 1.0 or 2.0 mg l−1), GA3 (0.0; 0.1; 0.5; 1.0 or 2.0 mg l−1) and AS (80.0 mg l−1). Every combination was repeated ten times. After 8 weeks in culture, somatic embryos in cotyledonary stage were counted.
Embryo conversion into plantlets
Embryos derived from three studied suspensions were implanted into 0.5 MS medium. To each jars containing 25 ml of medium, five well-developed embryos in cotyledonary stage were implanted. For each cell suspension ten replicates were done. Culture were carried out during 8 weeks and after that were scored.
Flow cytometry analysis of regenerants
DNA content was determined in 25 and 43 regenerants derived from cotyledon and hypocotyl suspension, respectively. Young leaves of seedling and suspension cultures which serve as a source of somatic embryos were used as a control. The youngest leaves were chopped fine with a razor blade in a Petri dish containing 1 ml of nucleus isolation buffer (0.1 M Tris, 2.5 mM MgCl2, 85 mM NaCl, 0.1% Triton X-100; pH 7.0) with propidium iodide (50 mg ml−1) and RNase (50 μg ml−1). Suspension of nuclei was passed through a 50-μm mesh nylon filter and measured in flow cytometer (Partec, Münster, Germany). Percentage of nuclei containing 2C and 4C and was calculated with the using DPAC V.2.2 software (Thiem and Śliwińska 2003).
Statistical analysis
Data were evaluated by analysis of variance using Type III sum of Squares (STATGRAPHICS Plus 2.1 for Windows). The least significant differences (LSD) between mean values derived from data were verified with Fisher test. Differences significant at the 95% probability level were counted for each pair of means.
Results
Characteristic of the cell suspension cultures
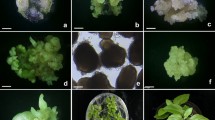
The cotyledon and hypocotyl derived suspensions of G. kurroo were established during 6 weeks (Fig. 1a). It was impossible to obtain a fully stabilized root derived suspension. The suspension mostly consisted of elongate cells with large, single vacuole (Fig. 1b). Prolific established cotyledon- and hypocotyl derived suspensions formed different size of aggregates (Fig. 1c). The diversification of the aggregate structure depended on the fraction. Cells of size <150 μm were round shaped, divided longitudinally and transversely creating small aggregates (Fig. 2a). They possessed dense cytoplasm, thick external wall, centrally located big nuclei and small vacuoles (Fig. 2b). In the cytoplasm, numerous amyloplasts, Golgi apparatus, rough endoplasmic reticulum, mitochondria and single lipid bodies were visible (Fig. 2c). In aggregates 150–300 μm in size, cells showed cytological differences depending on their location in the structure (Fig. 2d). In externally located cells, large amyloplasts with numerous starch grains were observed (Fig. 2e). Internal cells were smaller and compact, and contained fewer plastids with small starch grains and many lipid bodies (Fig. 2f). Their ultrastructure was similar to the fraction of <150 μm. Aggregates of >300 μm in size possessed three different layers of cells. The first, most internal, led strongly vacuolated cells. The second ones had the storage layer, the third ones were externally layer of merystematic character. The centrally located layer consisted of aged cells with little cytoplasm and large vacuoles, which succumbed and led to aggregate breakdown. In the storage layer, cells were smaller, and rich in amyloplasts with slight starch grains, small vacuoles and large nuclei. The most external layer was strongly separated from the culture medium by means of thick wall which was additionally covered by a protein–sucrose matrix. Walls between cells inside this layer were thinner with numerous plasmodesmata. Nucleus with homogeneous nucleoli took central place of cells. As the result of the cell division from external layers and single embryogenic cells, somatic embryos from globular (Fig. 2g) to cotyledonary stage were formed.

Characteristic of suspension cultures: a 3-month-old suspension in a conical flask (MS + 0.5 mg l−1 2,4-D + 1.0 Kin); b heterogeneous root-derived suspension culture with long shaped and round cells; c homogeneous cotyledon-derived embriogenic suspension; d somatic embryos after 3 weeks of culture onto conversion medium; e typical somatic embryo with small rootlet (R), short hypocotyl (H) and long, fused cotyledons (C); f embryo conversion into plantlets

Ultrastructure of suspension culture: a round shaped, divided longitudinal and transverse cells of size <150 μm; b visible dense cytoplasm, thick external wall (Cw), central located big nuclei (N) and small vacuoles (V) in single cells after division (×2,000); c numerous amyloplasts (Am), Golgi apparatus (GA), rough endoplasmic reticulum (rER), mitochondria (M) and single lipid bodies (Lb) in cells of fraction <150 μm (×6,000); d cell diversity in structure of aggregates 150–300 μm in size. External located cells with large amyloplasts (arrows) and internal smaller and compact cells; e external cell with amyloplast (Am) contained several grains of starch (S) (×8,000); f ultrastructure of internal cells, fraction of 150–300 μm: thin cell walls (Cw), amyloplasts (Am), small vacuoles (V), nucleus (N), mitochondria (M) and numerous lipid bodies (Lb); g somatic embryos in globular stage (arrows) formed from external situated cells of aggregates (×8,000)
Curves of the cell suspension growth
Two months after culture establishment, biomass growth was assessed. The growth curves of three studied suspensions were similar but differences in maximal value of biomass were observed (Fig. 3). During the first 9 days of the subculture the growth rate was low. From about day 9 suspension cultures started to grow very fast up to day 18 for K/R and day 21 for K/H and K/C. Later, the growth decreased. The highest amount of tissue (3.1 g) was obtained in the root-derived culture and the lowest (2.3 g) for hypocotyl-derived suspension. Dry weight for all studied G. kurroo suspensions was about tenfold lower than fresh weight. The growth coefficient was the highest for root-derived culture and make 6.7 and 6.1 for fresh and dry weight, respectively. Ratio of growth coefficient was the highest for cotyledon-derived cell suspension and reached 1.06.

Growth of fresh and dry masses depending on the origin of suspension culture (K/C, K/H, K/R cotyledon-, hypocotyle-, root-derived cell suspension, respectively)
Morphogenic capacities of cell suspensions
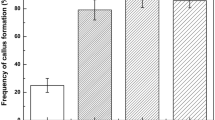
The highest amount of somatic embryos was achieved for K/C and the lowest for K/R culture. For all investigated suspensions, the decreasing of aggregate size also caused the decrease of the number of somatic embryos (Table 1). Statistical analysis proved the effect of GA3 and Kin concentrations on morphogenic potential of cell suspensions. From 100 mg of K/C aggregates >500 μm in size, 813 embryos were obtained (Table 1). The decrease of their number was caused by decreasing of Kin concentration up to the level 1.0 mg l−1 with or without low concentration of GA3. Aggregates 300–500 μm in size required similar concentration of plant growth hormones, but 2.0 mg l−1 Kin was most beneficial. After 6 weeks of culture for 150–300 μm fraction, the highest number of somatic embryos were obtained when the medium was only supplemented with 0.1–0.5 mg l−1 Kin and 80.0 mg l−1 AS. The lowest number of somatic embryos was produced by aggregates <150 μm in size. For hypocotyl-derived suspension culture the best results for all fractions was achieved on MS medium supplemented with 0.5 mg l−1 GA3, 1.0 mg l−1 Kin and 80 mg l−1 AS. The highest number of somatic embryos reached 163.5 (fraction >500 μm) and was fivefold less than for cotyledon origin suspension. The lowest morphogenic capacities were presented by root-derived suspension culture, about 29-fold less than for cotyledon ones.
Embryo conversion into plantlets
During the 8-week-long culture, plantlets reached the few leaf stage with very well-developed root systems. The yielding of conversion was origin dependent and the following data were achieved: 87, 85 and 68% for K/C, K/H and K/R, respectively.
Flow cytometry analysis
Among 68 regenerants studied the majority of them presented 2C DNA level. In case of K/H regenerants, 86% possessed 2C DNA and 4C DNA was presented by 14% of regenerants. K/C regenerants appeared more stable and 100% of them showed only the same contents of 2C DNA as control plants.
Discussion
Morphogenic competence of investigated suspension cultures depended on: (1) initial explant origin, (2) size of aggregates and ultastructure of their cells, and (3) concentration of the plant growth hormones.
The data presented in this paper gave evidence of very prolific somatic embryo production by two out of three studied cell suspensions of G. kurroo. Liquid culture is a unique system for cell proliferation because of its easy access to medium ingredients by the cells, better conditions for aeration, and easier possibilities to control the pH level. The rotation of the medium helps to mix the medium, metabolites and disperse oxygen. Induction medium which was used for suspension culture multiplication led to their maintaining embryogenic potential. At previously reported, auxin-cytokinin medium favors somatic embryo formation in gentian cultures (Mikuła et al. 2005; Bach and Pawłowska 2003). The embryogenic character of the cell found its expression on the ultrastructural level. The majority of published papers underline the presence of amyloplasts and starch grains as the ultrastructural marker of embryogenecity. Our studies revealed that the lipid bodies could be included in the matter.
Lipid bodies were also observed in the rice epithelial cells which are presumed to be embryogenically determined cells (Yeung 1995) and were found in developing somatic embryos (Hakman and von Arnold 1985; Joy et al. 1991; Avjioglu and Knox 1989). Results confirmed previously described ultrustructural features typical for other gentian species and are complementary for them (Mikuła et al. 2001).
To get more precise results, considering yield and quality of somatic embryos, it was necessary to use fractioned suspensions for implantation on agar media in our experiments. It was proved that for selected species of Gentianaceae the different size of aggregates was connected with the somatic embryo differentiation. Exacum affine formed the highest number of somatic embryos from fraction 100 μm (Ørnstrup et al. 1993), but in the case of Lisianthus russellianus the best results were obtained from aggregates >500 μm (Ruffoni and Massabò 1996). The fractionation use is one of methods to obtain synchronization of somatic embryos development (Nomura and Komamine 1995; Kim and Soh 1996; Choi et al. 1997). The asynchrony was present even when fractionation was employed in G. kurroo culture.
In our studies, the high level of Kin caused continuous embryogenic proliferation, and very often a second generation of embryos appeared, which resulted in obtaining the highest number of embryos regenerated from cotyledon-derived suspension culture. GA3 helps to produce properly formed embryos. The stimulus role of GA3 in plant regeneration systems is well known. Gibberellins stimulate further development of somatic embryos or make possible their conversion. The lack of GA3 in the suspension culture caused the stopping of the somatic embryo development at the cotyledon stage (Choi et al. 1999; Shimizu et al. 1997). It is worth mentioning that morphological disturbances of G. kurroo embryos were observed, namely twin embryos, some possessing numerous cotyledons and also somewhat hyperhydrated. The highest level of morphogenic disturbances was observed for >500 μm fraction. However, the embryos which presented morphological disturbances correctly passed conversion to reach properly developing plantlets.
In conclusion, our experiments revealed that long-term culture of studied cell suspensions had a direct relationship with the level of morphogenic competence typical for explants of their origin. The developed system of plant regeneration could be easily employed for plant genetic manipulation and biotechnological purposes.
Abbreviations
- 2,4-D:
-
2,4-Dichlorophenoxyacetic acid
- Kin:
-
6-Furfurylamonopurine
- GA3 :
-
Gibberellic acid
- AS:
-
6-Aminopurine hemisulfate
- MS:
-
Murashige and Skoog medium (1962)
- IM:
-
Initiation medium
- K/C:
-
Cotyledon derived suspension culture
- K/H:
-
Hypocotyl derived suspension culture
- K/R:
-
Root derived suspension culture
References
Avjioglu A, Knox RB (1989) Storage lipid accumulation by zygotic and somatic embryos in culture. Ann Bot 63:409–420
Bach A, Pawłowska B (2003) Somatic embryogenesis in Gentiana pneumonanthe L. Acta Biol Crac 45(2):79–86
Choi YE, Kim JW, Soh WY (1997) Somatic embryogenesis and plant regeneration from suspension cultures of Acanthopanax koreanum Nakai. Plant Cell Rep 17:84–88
Choi YE, Kim JW, Yoon ES (1999) High frequency of plant regeneration via somatic embryogenesis from callus or cell suspension cultures in Eleutherococcus senticosus. Ann Bot 83:308–314
Hakman I, von Arnold S (1985) Plantlet regeneration through somatic embryogenesis in Picea abies (Norway spruce). J Plant Physiol 121:149–158
Hosokawa K, Matsuki R, Oikawa Y, Yamamura S (2000) Production of transgenic gentian plants by particle bombardment of suspension-culture cells. Plant Cell Rep 19:454–458
Hosokawa K, Oikawa Y, Yamamura S (1998) Mass propagation of ornamental gentian in liquid medium. Plant Cell Rep 17:747–751
Joy IVRW, Yeung EC, Kong L, Thorpe TA (1991) Development of white spruce somatic embryos: I. Storage product deposition. In Vitro Cell Dev Biol 27P:32–41
Kamińska D, Wesołowska M, Króliczak P, Skrzypczak L, Grajek W (1998) Growth kinetics of cell cultures of Gentiana cruciata L., Gentiana tibetica King and gentiopicroside production. Biotechnology 3:107–113
Kim JW, Soh WY (1996) Plant regeneration through somatic embryogenesis from suspension cultures of Allium fistulosum L. Plant Sci 114:215–220
Mikuła A, Rybczyński J, Tykarska T, Kuraś M (2001) Somatic embryogenesis of Gentiana genus II. Scanning and ultrastructural analysis of early stages of somatic embryogenesis in liquid medium. Biol Bull Poznań 38:47–53
Mikuła A, Rybczyński JJ, Skierski J, Latkowska MJ, Fiuk A (2005) Somatic embryogenesis of Gentiana genus IV.: characterization of Gentiana cruciata and Gentiana tibetica embryogenic cell suspensions. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, Dordrecht, pp 345–356
Mikuła A, Skierski J, Rybczyński JJ (2002) Somatic embryogenesis of Gentiana genus III. Characterization of three-year old embryogenic suspensions of G. pannonica originated from various seedling explants. Acta Physiol Plant 24:311–322
Mikuła A, Tykarska T, Kuraś M, Iwanowska A, Rybczyński JJ (1996) Studies on the morphogenic potential of gentians in cell suspension. In: Proceedings of the international symposium. breeding research on medicinal and aromatic plants, Quedlinburg, Germany, pp 290–295
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Momčilovič I, Grubišić D, Nešković M (1997) Micropropagation of four Gentiana species (G. lutea, G. cruciata, G. purpurea, G. acaulis). Plant Cell Tiss Org Cult 49:141–144
Morgan ER, Butler RM, Bicknell RA (1997) In vitro propagation of Gentiana cerina and Gentiana corymbifera. NZ J Crop Hort 25:1–8
Nomura K, Komamine A (1995) Physiological and Biochemical aspects of somatic embryogenesis. In: Thorpe AT (ed) In vitro embryogenesis in plants. Kluwer, Dordrecht, pp 155–203
Ørnstrup H, Mølgaard JP, Farestveit B (1993) Somatic embryogenesis and plant regeneration from cell suspensions of Exacum affine. Plant Cell Tiss Org Cult 35:37–41
Preil W (2005) General introduction: a personal reflection on the use of liquid media for in vitro culture. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, Dordrecht, pp 1–18
Raina R, Behera MC, Chand R, Sharma Y (2003) Reproductive biology of Gentiana kurroo Royle. Curr Sci 5:667–670
Ruffoni B, Massabò F (1996) Plant production by somatic embryogenesis in cell suspension cultures of Lianthus russellianus Hook. Plant Tissue Cult Biotech 4:194–198
Sharma N, Chandel KPS, Paul A (1993) In vitro propagation of Gentiana kurroo—an indigenous threatened plant of medicinal importance. Plant Cell Tiss Org Cult 34:307–309
Shimizu K, Nagaike H, Yabuya T, Adachi T (1997) Plant regeneration from suspension culture of Iris germanica. Plant Cell Tiss Org Cult 50:27–31
Thiem B, Śliwińska E (2003) Flow cytometric analysis of nuclear DNA content in cloudberry (Rubus chamaemorus L.) in vitro cultures. Plant Sci 164:129–134
Yamada Y, Shoyama Y, Nishioka I, Kohda H, Namera A, Okamoto T (1991) Clonal micropropagation of Gentiana scabra BUNGE var. buergeri Maxim. and examination of the homogeneity concerning the gentiopicroside content. Chem Pharm Bull 39:204–206
Yeung EC (1995) Structural and developmental patterns in somatic embryogenesis. In: Thorpe AT (ed) In vitro embryogenesis in plants. Kluwer, Dordrecht, pp 205–247
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fiuk, A., Rybczyński, J.J. Factors influencing efficiency of somatic embryogenesis of Gentiana kurroo (Royle) cell suspension. Plant Biotechnol Rep 2, 33–39 (2008). https://doi.org/10.1007/s11816-008-0045-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11816-008-0045-8