
Abstract
We propose that oxidative stress resulting from an imbalance between generation and scavenging hydrogen peroxide contributes to tissue regeneration efficiency during somatic embryogenesis of hexaploid winter wheat (Triticum aestivum cv. Kamila) and organogenesis of faba bean (Vicia faba ssp. minor cv. Nadwislanski). Endogenous hydrogen peroxide content and antioxidant capacity of cells were determined in initial explants and callus cultures derived from these explants. Regeneration-competent explants (immature embryos) contained more endogenous H2O2 than explants initiated from regeneration-recalcitrant tissue (mature wheat embryos and faba bean epicotyls). Higher H2O2 levels were observed despite the higher activity of antioxidative enzymes (superoxide dismutase and catalase) and the induction of their gene expression. Calli originating from immature embryos retained the capacity of the initial explants: high H2O2 production was observed during the whole culture period. Low temperature treatment (4°C) was found to be an effective factor, which improved both regeneration ability and H2O2 production. Exogenous application to the medium of H2O2 and catalase blocker (3-aminotriazole), but not FeEDTA and superoxide dismutase blocker (diethyldithiocarbamate), also resulted in the enhancement of regeneration efficiency. These results clearly indicate that plant regeneration is specifically regulated by endogenous H2O2 and by factors, which improve its accumulation. Moreover, a study of the activity of various SOD isoforms suggests that not only the absolute concentration of H2O2, but also its localisation might be responsible for controlling regeneration processes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The development of biotechnology provides an additional incentive to continue research efforts aimed at optimising the conditions under which it would be possible to obtain many genetically identical regenerants. Somatic embryogenesis and organogenesis are the most commonly used pathways to induce efficient regeneration processes (Halperin 1995; Krikorian and Simola 1999; Jiménez and Bangerth 2001; Rose et al. 2010). However, some species are recalcitrant to regeneration, meaning that the efficiency of shoot induction is very low (Merkle et al. 1995; Rao 1996; Jiménez and Bangerth 2001). Due to the fact that regeneration is a multi-step process, at the present time, it is still not obvious, which changes are crucial to develop cell competence of forming a new plant (Toonen and de Vries 1996; Rose et al. 2010). It has been suggested that plant totipotency, which is necessary for shoot regeneration, is the result of gene expression, which is blocked in non-totipotent plants (Papadakis et al. 2001). However, other studies suggest that the number of genes specifically expressed during regeneration is rather limited (Dodeman and Ducreux 1996; Schrader et al. 1997; Dong and Dunstan 2000; Daimon et al. 2003); these genes include the SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE (SERK) (Schmidt et al. 1997; Somleva et al. 2000) and a family of KNOTTED 1-like homeobox genes (knox) (Hjortswang et al. 2002; Zhang et al. 2002; Hake et al. 2004).
The regeneration process is undoubtedly controlled from the gene level; however, proper physical and chemical conditions are necessary to initiate the regeneration programme encoded in the genome and/or to increase regeneration efficiency (Loschiavo et al. 1989; Imani et al. 2001; Padmanabhan et al. 2001; Papadakis et al. 2001; Chiappetta et al. 2006; Gupta 2011). The regeneration state has been induced by a variety of procedures, including exposure to plant growth regulators (Szechynska-Hebda et al. 2007), pH shock, temperature shock, osmotic and/or starvation stress (Merkle et al. 1995; Touraev et al. 1997; Zavattieri et al. 2010; Gupta 2011), and treatment with various chemical substances (Krikorian and Simola 1999; Gupta 2011).
On the other hand, these factors could also induce oxidative stress (Prasad et al. 1994; Rai et al. 2011). It has been generally accepted that reactive oxygen species (ROS) overproduction under stress is a detrimental factor, which causes lipid peroxidation, enzyme inactivation and oxidative damage to DNA, ending in cell dysfunction and death (Scandalios 1993; Pellinen et al. 2002; Vranová et al. 2002). However, during recent years evidence has accumulated regarding the participation of ROS in a signal transduction cascade (Prasad et al. 1994; Pellinen et al. 2002; Zavattieri et al. 2010); and their involvement in various metabolic and developmental processes (Inzé and Montagu 1995; Mehlhorn et al. 1996; Foyer et al. 1997; Ślesak et al. 2007; Rai et al. 2011).
H2O2 is the most stable compound among the ROS, and the most probable candidate for ROS-mediated signal transduction. It is able to penetrate the plasma membrane as an uncharged molecule, thus allowing it to be transported to the site of action (Foyer et al. 1997; Henzler and Steudle 2000; Vranová et al. 2002). A wide range of endogenous concentrations of H2O2 have been reported, from nmols to several hundred μmols of H2O2 per g fresh weight (Veljovic-Jovanovic et al. 2002; Szechyńska-Hebda et al. 2010). Additionally, the H2O2 content can change significantly within a very short period of time; for example, in a wheat leaf, chilling at 4°C caused the H2O2 level to increase threefold within 1 min (Okuda et al. 1991). In order to deal with these rapid dynamic changes in the endogenous hydrogen peroxide content, cells have developed antioxidative systems, which maintain the balance between hydrogen peroxide production and detoxification. The cellular enzymatic machinery for H2O2 control comprises several enzymes, which often display synergetic action. Superoxide dismutase (SOD) catalyses the disproportion of O ·−2 to H2O2 and dioxygen (Scandalios 1993). Different SOD isoforms are specifically localised in the cytoplasm, mitochondria and chloroplasts (Scandalios 1993; Jevremović et al. 2010). H2O2 is removed by catalase (CAT) in microbodies (peroxisomes, glyoxysomes) and mitochondria, as well as by different isoforms of peroxidases in chloroplasts, cytoplasm, peroxisomes, mitochondria and cell walls (Mittler and Zilinskas 1991; Asada 1992; Scandalios 1993; Willekens et al. 1997; Karpinski et al. 1999).
H2O2 could play an important role in plant regeneration processes (de Marco and Roubelakis-Angelakis 1996a; b; Cui et al. 1999; Zavattieri et al. 2010; Zhang et al. 2010), but the specifics of its role are still unclear; it has been proposed both to contribute to recalcitrance and to induce gene expression and synthesis of proteins which aid embryogenesis (Cutler et al. 1991; Roubelakis-Angelakis 1993; Papadakis and Roubelakis-Angelakis 1999; Papadakis et al. 2001). Changes in the activity of antioxidative enzymes have also been detected during shoot initiation and development in the regeneration process (Kairong et al. 1999; Qiusheng et al. 2005; Szechynska-Hebda et al. 2007).
The aim of our study was to determine: (1) whether oxidative stress induced by low and high temperatures can play a positive role in embryogenesis and organogenesis; (2) which oxidative/antioxidative abilities of initial cells are modified by temperature stress; (3) whether H2O2 could be a universal second messenger in the regeneration processes. To answer these questions, we assessed the hydrogen peroxide levels, changes in antioxidative enzyme activity and their gene expression in the regeneration-competent and non-regeneration-competent cells of initial explants and their corresponding calli before and after the application of stress conditions. We chose to compare the callus cultures of two different plant species, i.e. wheat (monocotyledon) and faba bean (dicotyledon), in order to investigate whether particular features are unique or are generally applicable to other species.
Materials and methods
Plant material, growth conditions, in vitro culture
Experiments were carried out on tissues of two crop species: hexaploid winter wheat (Triticum aestivum L.) cultivar Kamila and faba bean (Vicia faba ssp. minor) cultivar Nadwislanski.
Seedlings of wheat were vernalised for 9 weeks in moist perlite (5°C, 8-h photoperiod), and then planted in soil under greenhouse conditions (20/17°C, 16-h photoperiod). Some spikes were cut 14 days after pollination for the isolation of immature embryos, and the remaining spikes were kept until maturity. Both immature and mature kernels were surface-sterilised by brief immersion in 70% ethanol, followed by a 20-min wash with 10% solution of commercial bleach (Domestos), and then rinsed with sterile water. Immature and mature embryos were aseptically excised and placed on a culture medium with their axes adjacent to the medium surface. Callus was induced for 4 weeks (25°C, 16-h photoperiod 20 μM m−2 s−1) on MS medium (Murashige and Skoog 1962) supplemented with 2 mg l−1 2,4-dichlorophenoxyacetic acid (2,4-D), 30 g l−1 sucrose, and 6 g l−1 agar (pH 5.8). Callus initiated from the immature embryos, unlike that from the mature embryos, was capable of shoot regeneration in optimal conditions.
Plants of faba bean were grown on experimental fields with a sowing density of 30 seeds per square metre. Young pods, 4–5 cm long, were harvested 10 weeks after sowing. Pods were surface-sterilised by immersion in 20% solution of commercial bleach for 45 min, then rinsed with sterile water. Seeds were aseptically excised from pods, and immature embryos were isolated and cultured. For collection of epicotyls, dry mature seeds of faba bean were sterilised (20% solution of commercial bleach for 45 min) and germinated on 3% agar in darkness for 5–6 days. Both types of explants (immature embryos and epicotyls) were cultured on MS medium containing 1 mg l−1 casein hydrolysate, 750 mg l−1 myo-inositol, 1 mg l−1 6-benzyl-aminopurine (BAP), 0.5 mg l−1 naphthalene acetic acid (NAA), 0.25 mg l−1 gibberellic acid (GA3), 30 g l−1 sucrose, and 6 g l−1 agar (pH 5.8). The cultures were grown for 4 weeks at 25°C using a 16-h photoperiod (20 μM m−2 s−1). Callus initiated from immature embryos was potentially capable of shoot regeneration in optimal conditions, whereas callus initiated on epicotyls was not capable of regeneration.
Explants and calli induced from wheat and faba bean explants served as the experimental material.
Exposure to temperature stress
To study the effect of temperature stress, 4-week-old calli of both species were transferred onto a fresh callus-inducing medium (MS with growth regulators) and exposed to 4, 30 and 25°C (control) for 2 weeks. Next, all cultures were transferred onto a growth regulator-free MS medium (the regeneration medium). Material for analyses was collected 3 h and 1, 2, 4, 7, 14 and 28 days after transferring to the regeneration medium.
Regeneration efficiency
Callus cultures were kept for four weeks on MS regeneration medium (without growth regulators) in conditions optimal for regeneration (25°C, 16-h photoperiod, 20 μM m−2 s−1). To study the effect of stress level on regeneration efficiency, the medium was supplemented with hydrogen peroxide (200 mM) and ferric ethylenediaminetetraacetic acid (FeEDTA, 50 mM), as well as diethyldithiocarbamate (DDC, SOD inhibitor, 0.1 mM) and 3-aminotriazole (AT, CAT inhibitor, 0.02 mM). Callus cultures grew on these media for 4 weeks. Regeneration efficiency was expressed as the frequency of calli regenerating shoots (%).
Determination of H2O2 level
Total H2O2 content was measured in fresh material by a fluorometrical assay with homovanillic acid according to Ishikawa et al. (1993). Samples (0.4 g) were homogenised in 0.6 ml of ice-cold 5% TCA and centrifuged at 4,000g for 5 min. The reaction mixture contained the supernatant (0.3 ml), 1.25 mM homovanillic acid, l unit of horseradish peroxidase (Sigma-Aldrich Co., Poland), and 25 mM potassium phosphate buffer (pH 7.5). The fluorescence yield was measured at the excitation wavelength of 315 nm and emission of 425 nm.
Enzyme assays
Tissues were homogenised with 1% PVP-40 and 0.05 M phosphate buffer (pH 7.0) containing 0.1 mM (EDTA) at 4°C. The plant biomass:extraction buffer (w:v) ratio was 1:2. The homogenate was centrifuged at 15,000g for 10 min and the supernatant was dialysed overnight in 0.05 M phosphate buffer.
Total SOD [EC 1.15.1.1.] activity was assayed according to the modified method of McCord and Fridovich (1969). The reaction mixture consisted of 0.05 M phosphate buffer, 0.013 mM cytochrome c, 0.1 mM xanthine, 0.024 U per ml xanthine oxidase, and supernatant. Absorbance was measured at 550 nm. One unit was defined as the amount of enzyme necessary for 50% inhibition of cytochrome c in a coupled system with xanthine and xanthine oxidase.
Catalase [EC 1.11.1.6] activity was determined according to Aebi (1984). The reaction mixture consisted of 0.05 M phosphate buffer, 0.1 mM H2O2, and supernatant. The rate of H2O2 decomposition at 240 nm was measured. One unit of CAT activity was defined as the amount of enzyme that decomposed 1 μM of H2O2 per minute under standard conditions.
The reaction kinetics of enzymes were measured over 2 min at 25°C using a Perkin Elmer UV–vis spectrophotometer. Enzyme activities were calculated per mg protein.
For SOD isoenzyme analysis, tissue samples were homogenised in ice-cold protein extraction phosphate buffer (0.05 M, pH 6.8) containing 0.1 mM EDTA, 0.1% N,N,N′,N′-tetramethylenediamine (TEMED), 1 mM dithiothreitol (DTT), and 0.1% ascorbic acid. The homogenate was centrifuged for 10 min at 15,000g. The supernatants were decanted and the volumes were adjusted to obtain equal protein concentrations, then 50 μg of protein was loaded onto each well of the gel. SOD isozymes were separated by native PAGE using 11.5% polyacrylamide gels and 0.l M Tris–glycine as an electrode buffer. Electrophoresis was performed at 4°C at 60 mA (Bio-Rad Mini Protean power supply). Gels were stained by the method of Beauchamp and Fridovich (1971) in phosphate buffer (pH 7.8) containing 0.1 mM EDTA, 0.5 μM nitroblue tetrazolium (NBT), 26.5 mM riboflavin, and 26.5 μM TEMED in the dark for 20 min. Gels were exposed to the light, about 100 μM m−2 s−1 photon flux density (PPFD), for 30 min in 0.05 M phosphate buffer, and then transferred to 1% (v/v) acetic acid to stop the reaction. The isoenzyme profile was differentiated using inhibitors of Cu/ZnSOD and FeSOD (1 mM KCN and 5 mM H2O2, respectively) according to Beauchamp and Fridovich (1971).
Proteins were assayed by Bradford’s dye-binding technique (1976) with bovine serum albumin as a protein standard.
RNA extraction and northern hybridisation
Total RNA used for the Northern blot analysis was isolated according to the protocol of TRI-Reagent of MRC, Cincinnati (Chomczynski and Sacchi 1987). In order to remove any contamination with DNA, total RNA was treated with RNAse-free DNAse at a concentration of 1 U/ml (Promega). Total RNA was quantified with a spectrophotometer (GeneQuant, Pharmacia), and further confirmed by gel electrophoresis and ethidium bromide staining. Twenty micrograms of total RNA was separated on a 1.2% agarose gel containing 6% formaldehyde (ICN Biomedicals Inc.) and blotted onto a nylon membrane (Hybond-N+) by standard capillary blotting, according to Sambrook et al. (1989). RNA was fixed on the membranes using 30 s of UV exposure (GS Gene Linker, BioRad). MnSOD3.1 cDNA from T. aestivum (GenBank accession no. U72212) length 934 bp (Wu et al. 1999) was used as a probe for wheat tissue, and the molecular probe of MnSOD, obtained by RT-PCR amplification of the 318-bp fragment, was used for the faba bean experiment (Dąbrowska et al. 2007). The molecular probe for Cat2 expression analysis was prepared by RT-PCR amplification with C1 (5′-cgtctggaacaacaacaacg-3′) and C2 (5′-tccatccctgctgattaagg-3′) primers. RT-PCR wheat products of length 580 bp were cloned into pCRII-TOPO Vector (Invitrogen Life Technologies, California, USA). The cloned cDNA was sequenced using fluorescent primers and an automated DNA sequencer (DNA Sequencing and Oligonucleotide Synthesis Laboratory of the Institute of Biochemistry and Biophysics PAS, Warszawa). The cDNA of Cat1 of cassava (Manihot esculenta) length 1,300 bp (Reilly et al. 2001) was a molecular probe for hybridisation with RNA from tissues of the faba bean. The membranes hybridised with specific probes corresponding to SODs and CATs were labelled with [α-32P]–dCTP (20 μCi) using the T7 Transcription Kit (Fermentas GMBH, Germany). Hybridisation was carried out with cDNA probes at 65°C for 16 h in a hybridisation buffer containing 0.5 M sodium phosphate buffer (pH 7.2), 7% w/v sodium dodecyl sulphate (SDS), 1 mM EDTA, and 1% w/v bovine albumin (BSA). The membranes were washed in 2× SSC and 0.1% SDS at room temperature for 15 min and then twice in 0.2× SSC and 0.1% SDS for 20 min at 60°C. After washing, membranes were exposed to X-ray films (X-Omat AR, Kodak) at −80°C, which were analysed using Phosphoimager FLA3000 (Fuji).
Statistical analysis
All of the experiments were performed with 20 calli per Petri dish for wheat and 10 per dish for faba bean. One dish was regarded as a replicate for the regeneration experiment. Regeneration ability was calculated from five dishes. All physiological measurements were performed in at least ten replicates. The replicate was tissue from different Petri dishes in the required amounts. Northern blot analysis was repeated three times. Data were analysed by calculating means, standard deviation values and statistical significance were analysed by Duncan’s test (P < 0.05).
Results
Effect of stress conditions on regeneration ability
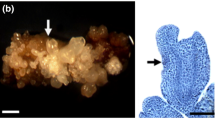
Young, small cells indicating intensive cell division were observed in 4-week-old calli induced from immature embryos (potentially competent to regenerate) in both wheat (Fig. 1a) and faba bean (Fig. 1g). After subculture onto the regeneration medium under control conditions (25°C), embryo-like structures arose from clusters of cells of wheat callus, while faba bean shoots were produced via organogenesis. In both species, shoot formation was first observed on the seventh day of subculture, and the maximum number of emerging shoots was reached in the second week. Regeneration efficiency was 33% for wheat and 30% for faba bean (Fig. 1b, h), respectively, in the second week after subculture, and was profoundly improved when the tissues were pre-treated with cold (applied before subculture onto the regeneration medium), increasing to 72% for wheat and 95% for faba bean (Fig. 1c, i). Conversely, pre-treatment of the calli at 30°C blocked their regeneration potential, and no shoot formation was obtained (Fig. 1e, k).

Microscopic images of 4-week-old calli before transfer onto the regeneration medium. Calli were induced from immature embryos (explants potentially capable of regeneration) of wheat (a) and faba bean (g) as well as from mature wheat embryos (d) and faba bean epicotyls (j) (explants non-capable of regeneration). The figure also presents the effects of culture conditions on callus regeneration efficiency of wheat (a–f) and faba bean (g–l) after 2 weeks of subculture on the regeneration medium. Regenerating callus cultures derived from immature embryos under control conditions at 25°C (b, h); regenerating callus cultures derived from immature embryos after 2 weeks of cold pre-treatment (at 4°C) followed by 2 weeks of subculture at 25°C (c, i); regenerating callus cultures derived from immature embryos after 2 weeks of heat pre-treatment (at 30°C) followed by 2 weeks of subculture at 25°C (e, k); non-regenerating culture derived from wheat mature embryos and faba bean epicotyls under control conditions at 25°C (f, l). The figure shows representative examples of regeneration from at least five replicates (Petri dishes). Values express the regeneration efficiency as the frequency of calli regenerating shoots (%)
Calli induced from mature wheat embryos and faba bean epicotyls were built of larger, highly vacuolated cells (Fig. 1d, j, respectively) and did not regenerate plantlets on the regeneration medium in control conditions (25°C) (Fig. 1f, l), even after low temperature pre-treatment (not shown in Fig. 1).
H2O2 level and antioxidant enzyme activity
To test whether cold and heat pre-treatment caused oxidative stress, and to examine whether regeneration ability was dependent on the abilities of the cells’ antioxidative machinery, cellular hydrogen peroxide content and the activities of the antioxidative enzymes were studied in wheat and faba bean tissues competent (R) or non-competent (NR) to regenerate. As H2O2 content and enzymatic response may vary with tissue origin, these parameters were compared in the initial explants (immature and mature wheat embryos, faba bean epicotyls), in calli before transfer onto the regeneration medium, and in tissue under conditions stimulating (control, 4°C pre-treatment) or inhibiting the regeneration process (30°C pre-treatment).
Figure 2 shows the quantification of H2O2 in initial explants and the corresponding calli before culture on the regeneration medium. In wheat and faba bean tissues, the total H2O2 content was related to future shoot production capacity. Accumulation of hydrogen peroxide was significantly higher in competent than in non-competent initial explants, and the enhanced H2O2 accumulation was maintained in R calli.

Endogenous hydrogen peroxide levels and total activity of antioxidant enzymes (SOD and CAT) in wheat and faba bean immature embryos (R initial explants potentially capable of regeneration), in wheat mature embryos and faba bean epicotyls (NR initial explants non-capable of regeneration), and in corresponding calli before temperature treatment and subculture on the regeneration media. All data are mean ± SE from 15 (n = 15) independent samples, shown with error bars corresponding to the standard deviation. Statistical significances are indicated with different letters according to Duncan’s test (P < 0.05)
A relationship was also observed between regeneration competence and antioxidative enzyme activity (Fig. 2). The activity of both SOD and CAT was higher in all competent tissues, with the exception of SOD activity in faba bean explants. However, the differences between R and NR wheat calli were smaller than those observed for the explants.
Figure 3 shows the influence of cold and heat pre-treatment on H2O2 production in regeneration-competent and non-regenerating calli during 28 days of culture on the regeneration medium. Independently of pre-treatment, the R tissue of both species (Fig. 3a, b) showed enhanced accumulation of H2O2 in comparison to NR calli (Fig. 3c, d) during the whole culture period. The time course of H2O2 revealed that regenerating cultures in control conditions and after the cold pre-treatment accumulated more H2O2 at the beginning of the subculture, up to 4–7 days, at which point the first shoots were observed (Fig. 3a, b). Culture prolongation accompanied by shoot emergence caused a reduction of H2O2. The exception was faba bean tissue under control conditions (Fig. 3b). However, it should be noted that a large increase in H2O2 at the end of culture could be a result of the subculture lasting too long, and the resulting callus darkening. Pre-treatment at 4°C prevented callus darkening and H2O2 accumulation in the faba bean tissue, probably as a consequence of earlier adaptation to stress (Fig. 3b). H2O2 increase was not observed at the beginning of the subculture in regeneration-competent wheat and faba bean calli that had undergone 30°C pre-treatment (conditions inhibiting regeneration) (Fig. 3a, b). However, an increase in the H2O2 level similar to that seen in control regenerating tissue was caused by the prolongation of faba bean culture after heat pre-treatment. Non-regenerating tissue did not produce temperature-dependent diversity in the course of H2O2 (Fig. 3c, d).

Effects of cold (4°C) and heat (30°C) pre-treatment in comparison with the control condition (25°C) on the endogenous hydrogen peroxide concentration (a–d), superoxide dismutase activity (e, h) and catalase (i–l) in subsequent days of culture on the regeneration medium (0–28). H2O2, SOD and CAT in regenerating calli initiated from wheat (a, e, i) and faba bean (b, f, j) immature embryos; H2O2, SOD and CAT in non-regenerating calli initiated from wheat mature embryos (c, g, k) and faba bean epicotyls (d, h, l). All data are mean ± SE from 15 (n = 15) independent samples, shown with error bars corresponding to the standard deviation. Statistical significances are indicated with different letters according to Duncan’s test (P < 0.05) performed for different days of culture within selected condition (control, cold and heat)
The content of total hydrogen peroxide corresponded to antioxidant enzyme activity. Control conditions and cold pre-treatment induced an increase in the total SOD activity at the beginning of R calli subculture (Fig. 3e, f): the highest SOD activity was observed in wheat cultures pre-treated at 4°C in the second day. Heat did not produce such enzyme response. The increase in SOD activity consistent with the time of subculture was detected in most of non-regenerating tissue (Fig. 3g, h).
The time course of CAT activity exhibited a negative relationship with cellular H2O2 levels in conditions favouring regeneration (control, cold pre-treatment) (Fig. 3i, j). Low CAT values were observed at the beginning of R culture, and increased values after the seventh day (which corresponded to the first shoot formation). The heat treatment did not influence CAT activity in regenerating wheat cultures, while CAT activity was very high in faba bean R culture up to the seventh day. CAT activity in non-regenerating tissues was relatively stable and increased only in the latest stage of culture (Fig. 3k, l).
It is not just the absolute concentration of H2O2 that may determine the regeneration potential of tissue; its cellular localisation might be responsible for controlling reactions that are important for regeneration processes. To investigate this, we determined SOD isoenzymes by native gel electrophoresis (Fig. 4). In extracts from wheat tissue, three SOD isoenzymes were detected (1 band of MnSOD izoenzyme and 2 bands of Cu/ZnSOD isoenzymes), whereas in faba bean samples up to ten different bands of SOD izoenzymes could be distinguished (1–4 bands of MnSOD, 1 band of FeSOD, and 1–5 bands of Cu/ZnSOD). The most characteristic changes were observed for MnSOD isoforms for both species; MnSOD activity was enhanced in tissue competent to regenerate (explants, calli), in regenerating tissue in control conditions and after cold pre-treatment. Conditions inhibiting regeneration (heat pre-treatment) induced MnSOD in regeneration-recalcitrant tissue, and new isoforms with higher mobility were detected in the faba bean tissue.

Changes in SOD isoform patterns in immature wheat and faba bean embryos, in mature wheat embryos and faba bean epicotyls, in the corresponding callus before the temperature treatment, and in calli cultured on the regeneration media 3 h and 7 days after cold (4°C) and heat (30°C) pre-treatments in comparison with the control (25°C). R explants and calli potentially capable of regeneration, NR explants and callus non-capable of regeneration. The figure shows representative examples from three independent experiments
The chloroplastic isoform (FeSOD) was present only in faba bean non-competent explants (epicotyls) and R calli after cold pre-treatment. These results could be correlated with the presence of chloroplasts in the epicotyls (green tissue) and a lack of chloroplasts in other explants (the embryos did not develop mature chloroplasts). The presence of FeSOD could also be a consequence of chloroplast presence in R calli after cold pre-treatment (the fastest shoot formation on calli surfaces subcultured on the regeneration medium in comparison with the other cultures).
Cu/ZnSOD activity in R and NR tissue was ambiguous. In wheat tissue, the Cu/ZnSOD pattern did not change significantly. Differences were detected only in cultures after heat pre-treatment, where Cu/ZnSOD activity was higher in non-regenerating cultures. In faba bean explants, five bands of Cu/ZnSOD were observed in competent tissue and three in non-competent tissue. Additional bands were also detected in regenerating calli 3 h after cold pre-treatment.
Northern blot
The response of the antioxidative enzymatic system during the regeneration processes was also studied by manganese superoxide dysmutase and catalase gene expression. Molecular probes of MnSOD showed a transcript of about 1,000 bp (Fig. 5). In competent explants and the corresponding calli (before subculture on the regeneration medium) of both species, a higher level of MnSOD expression was observed in contrast to the very weak signal in the non-competent tissue. A similar pattern was detected for calli on regeneration medium directly after temperature pre-treatment (3 h), with the exception of faba bean culture after heat pre-treatment. The highest signal was observed after cold pre-treatment. On the seventh day of wheat subculture, a decrease in signal intensity for R calli was observed for all conditions, whereas in faba bean culture the MnSOD gene was still induced in the regenerating tissues under control conditions and as a result of exposure to cold.

Northern blot analysis of mRNA levels, showing MnSOD and CAT expression. Transcript levels were analysed in wheat and faba bean explants competent (R) and non-competent (NR) to regenerate and in the corresponding calli, before the temperature treatment as well as in callus cultured on the regeneration media 3 h and 7 days after cold (4°C) and heat (30°C) pre-treatment in comparison with the control (25°C). The figure shows representative examples from three independent experiments
A transcript of the length of about 1,800 bp was identified by the radiolabelled molecular probe of catalase (Fig. 5). In wheat explants and calli, a higher CAT expression was observed in the NR tissues than in the R tissues. In faba bean cultures, similar changes were observed only for explants. For calli, a comparable intensity of signal was detected. The temperature pre-treatment affected the transcript level ambiguously. Higher expression was detected 3 h and 7 days after the cold pre-treatment in the R cultures of both species, compared to the NR cultures. Control conditions and heat pre-treatment increased the transcript level in wheat R tissue 3 h after subculture, and moreover, in wheat NR tissue on the seventh day of subculture. The opposite effects were observed for faba bean under control conditions and after heat pre-treatment.
Influence of medium supplements
To strengthen evidence of the hydrogen peroxide effect on regeneration ability, the modification of oxidative stress in regeneration-competent wheat culture was determined. Regeneration media were supplemented with substances, which enhance oxidative stress (H2O2 and FeEDTA) and inhibitors which affect the antioxidative enzyme system (DDC, SOD inhibitor; AT, CAT inhibitor). The presence of hydrogen peroxide in the medium greatly promoted the embryogenic potential and led to a significant shortening of the regeneration period (Fig. 6). Elevated shoot production was specific to H2O2 treatment; because the addition of a transition metal (Fe) that also causes oxidative stress in plant cells (Sinha et al. 1997) blocked the regeneration ability of the callus culture. Experiments with antioxidant enzyme inhibitors provided additional confirmation of this result. Shoot regeneration potential was repressed by the SOD inhibitor (DDC) and increased by the CAT inhibitor (AT). However, the addition of AT to the medium negatively influenced the quality of the regenerated shoots; some plantlets were albinotic.

Effects of medium supplements—hydrogen peroxide (200 mM) and FeEDTA (50 mM) as well as SOD inhibitor (DDC 0.1 mM) and CAT inhibitor (AT 0.02 mM)—on the regeneration efficiency of regenerating wheat culture. The figure presents representative examples of regeneration from at least five replicates (Petri dishes)
Discussion
The immature embryos of various species, including wheat and faba bean, have been regarded as the most suitable explants for the induction of morphogenic callus cultures and the regeneration of whole plants via indirect somatic embryogenesis and/or organogenesis (Benkirane et al. 2000; Tamás et al. 2004; Schryer et al. 2005; Szechynska-Hebda et al. 2007; Rose et al. 2010). Other explants have been shown to be more or less recalcitrant (Imani et al. 2001). The type of explants used in our study also determined the regeneration abilities in callus culture. Calli originating from wheat mature embryos and faba bean epicotyls had no regeneration capability, in contrast to the high regeneration efficiency seen in calli obtained from the immature embryos of both species.
The assumption underlying this study was the complex metabolic modifications that explants undergo during their isolation in and adaptation to in vitro conditions, which may include an alteration in the oxygen balance with an overproduction of active forms of oxygen. An excess of reactive oxygen in a cell may influence its regeneration capacity. It has been suggested that enhanced ROS production in initial explants can have an effect, from plant recalcitrance to regeneration (Benson and Roubelakis-Angelakis 1994; de Marco and Roubelakis-Angelakis 1996a; b; Zavattieri et al. 2010; Zhang et al. 2010; Rai et al. 2011); but despite this we conclude that mild oxidative stress and hydrogen peroxide-dependent metabolic pathways are crucial for in vitro shoot formation via somatic embryogenesis and organogenesis. During our experiments, a higher cellular H2O2 content was already observed in the immature embryos of both species, and it occurred despite an improved response of the antioxidative enzymes. In contrast, mature wheat embryos and faba bean epicotyls were characterised by low H2O2 levels as well as low activity of ROS-scavenging enzymes. This suggests the dependence of the oxidative state of cells on the age of the explants, which was previously also reported by Gara et al. (2003) and Gupta (2011). A similar relationship was also observed in calli; elevated H2O2 production and high enzymatic activity were observed in the small cells of competent calli with dense cytoplasm and compact nuclei. This microscopic appearance of cells suggests a morphologically younger age in comparison to the large, differentiated, highly vacuolated cells of non-competent calli (Fig. 1a, g compared to Fig. 1d, j). Maintenance of small cell volumes was previously believed to be one of the conditions needed to enable the onset of embryogenesis/organogenesis (Pellegrineschi et al. 2004) and to represent a dedifferentiated cell state with the potency to initiate a new developmental programme (Nomura and Komamine 1995; Pasternak et al. 2002). The conclusion from our results is that the cells of mature explants are able to divide and to initiate the callus in a similar way to the younger explants, but the “memory” of the origin being forwarded to the new generation of cells and the switch of the developmental programme to regeneration are dependent on the competence “inherited” from the cells of the initial explants. We cannot rule out the possibility that the “memory” of explant capacity could also determine the responses of regenerating and non-regenerating calli to various temperature conditions. Immature embryo-derived tissues in most cases showed higher sensitivity to cold pre-treatment in comparison with the non-regenerating calli. Regeneration efficiency, production of hydrogen peroxide and enzyme activity were all greatly enhanced, especially during the first week after 4°C pre-treatment. These results, taken as a whole, suggest that the factor responsible for maintaining regeneration potential in cells could be their oxidative/antioxidative capacity.
The study of regeneration efficiency on media with stress-enhancing supplements provides additional proof for this hypothesis and indicates that H2O2 is the most probable factor influencing cell morphogenesis. The regenerating tissues could tolerate high concentrations of exogenous H2O2 and even responded with a higher number of produced shoots. H2O2 treatment also influenced plantlet development and allowed a much earlier completion of plant ontogenesis (data not shown). The exogenous H2O2 was probably able to diffuse through the plasma membrane, reinforce internal resources and additionally improve cell response. Cell susceptibility to ROS appears to be specific to hydrogen peroxide; regeneration potential was blocked in the presence of another stress inducer—FeEDTA. Fe(2+) enhances ROS production (·OH, ·OOH) via the Fenton reaction with parallel consumption of H2O2. Additionally, FeEDTA increased the activity of the hydrogen peroxide scavengers (Pasternak et al. 2002) and caused a rapid accumulation of ascorbate peroxidase mRNA (Vansuyt et al. 1997; Yoshimura et al. 2000). Thus, the Fenton reactions and activation of H2O2-removing enzymes could have lowered the level of cellular H2O2 in our cultures on the medium supplemented with FeEDTA, and subsequently repressed the regeneration potential.
Moreover, the role of the enzymatic control of endogenous H2O2 during the determination of cell regeneration ability was shown during experiments with blockers of antioxidant enzymes. The application of a SOD-blocking agent (resulting in lower H2O2 production) decreased the regeneration potential, whereas the addition of a CAT-blocking agent (resulting in lower H2O2 consumption) increased the ability of the tissue to undergo embryogenesis. It follows that a higher hydrogen peroxide content specifically facilitated realisation of the developmental programme. However, CAT activity blocking also had the undesirable effect of producing albinism in some of the regenerated plantlets. It is likely that catalase is involved in plastid development and promotes the formation of green plants. This finding is consistent with results obtained for some maize mutants exhibiting a chlorophyll-less phenotype (Hu et al. 1998). The activity of catalase, a heme-requiring enzyme, was reduced by up to 100% in homozygotes when compared with the level detected in the wild-type, and a lack of catalase also possibly caused heme, and consequently, chlorophyll deficiency.
The effect of CAT blockers on green plant regeneration suggests that not only is absolute H2O2 concentration an important factor affecting cell response. An analysis of SOD isoenzyme patterns supports the thesis that subcellular ROS distribution and the place of its action may be crucial for regeneration processes. Higher MnSOD activity in initial tissue competent to regenerate and the calli derived from them indicate that mitochondria-originated H2O2 (MnSOD is mainly localised in the mitochondria) could be involved in the modification of regeneration potential. The amount of H2O2 produced in plant mitochondria is usually much less than that of chloroplasts when exposed to light. However, oxidative metabolism is the main source of ROS in non-green tissues (Vertucci 1989; Rhoads et al. 2006; Ślesak et al. 2007), and up to 2–4% of the oxygen metabolised by mitochondria could be converted to oxygen radicals (Morré et al. 2000) due to a less than fully efficient flow of electrons through the mitochondrial electron transport chain (Morré et al. 2000; Gille and Nohl 2001; Möller 2001). The predominance of respiration over photosynthesis also concerned our culture and was confirmed by direct respiration measurements (unpublished data) and the limited number of cells containing mature chloroplasts in competent explants and in regenerating calli at the beginning of culture (microscopic data and lack of FeSOD band on Fig. 4).
Additionally, the fact that the highest MnSOD activity was seen in callus cultures after cold pre-treatment indicates that mitochondria are an important source of superoxide, and subsequently, hydrogen peroxide at low temperature, which is in agreement with data obtained by Purvis et al. (1995) and Jevremović et al. (2010). This could be the result of more intensive production of ROS in the enhanced flow of electrons, as low temperatures caused enhanced respiration in the regenerating calli (unpublished data).
H2O2 does not act only in the mitochondria. Our data suggested the involvement of cytoplasmic isoforms of SOD in ROS control during regeneration processes. Competent explants, calli and tissue in conditions promoting regeneration showed higher Cu/ZnSOD activity in comparison to the non-competent tissue. These isoforms could contribute to H2O2 production in cytoplasm. Cytoplasmic H2O2 can act as a trigger for signal transduction or the oxidation of other molecules by H2O2-induced reactions (Desikan et al. 2003; Zhang et al. 2010). Finally, H2O2 might change gene expression and modify the activity of the transcription factors (Apel and Hirt 2004). Using cDNA microarray technology to carry out a transcriptomic analysis, Desikan et al. (2001) confirmed that the expression of some genes is up-regulated by H2O2. The H2O2-induced transcripts encoded proteins with functions such as metabolism, energy, protein destination and transport, cellular organisation and biogenesis and transcription (Desikan et al. 2001). Moreover, H2O2 regulated the expression of genes encoding antioxidant enzymes and modulators of H2O2 production (Neill et al. 2002; Geisler et al. 2006). In the present study, the gene expression of the enzyme controlling hydrogen peroxide levels was also enhanced in explants and the callus cultures capable of regeneration. Its expression was specifically modified by temperature treatment. However, it is difficult to state definitively whether the observed higher expression is a result of induced oxidative stress or whether the events underlying the regulation of cell division and further differentiation processes are under the tight control of enzymes, which are continuously highly expressed.
In conclusion, we point out the existence of a clear relationship between the tissue regeneration efficiency of wheat and faba bean cultures and H2O2 accumulation. However, we also note that the stress per se (the conditions enhancing H2O2) could not be the key factor in determining the morphological response. It has previously been reported that cold can induce the switch of the developmental programme from pollen development to in vitro androgenesis (Hou et al. 1993; Touraev et al. 1997; Obert et al. 2004; Xynias et al. 2001). In our cultures, cold pre-treatment and exogenous H2O2 strongly promoted regeneration efficiency in competent tissue, but did not break the internal barriers against induction of the regeneration process in non-competent tissue. Thus, we rather suggest that the competent culture might retain the “memory” of the explants and retain the capacity of the initial cells. Greater susceptibility and flexibility of the oxidative/antioxidative state of cells is proposed as the factor responsible for this retention of the potential for induction of the regeneration process. In particular, H2O2 is the best candidate for cellular signals that can regulate plant regeneration. However, the primary place of H2O2 action still requires further detailed study.
Abbreviations
- CAT:
-
Catalase [EC 1.11.1.6]
- NR:
-
Regeneration-recalcitrant tissue
- R:
-
Tissue potentially competent to regenerate, or regenerating tissue
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase [EC 1.15.1.1.]
References
Aebi H (1984) Catalase in vitro. Meth Enzymol 105:121–125
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–399
Asada K (1992) Ascorbate peroxidase: a hydrogen peroxide-scavenging enzyme in plants. Physiol Plant 85:235–241
Beauchamp CO, Fridovich I (1971) Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal Biochem 44:276–287
Benkirane H, Sabounji K, Chlyah A (2000) Somatic embryogenesis and plant regeneration from fragments of immature inflorescences and coleoptiles of durum wheat. Plant Cell Tiss Org Cult 61:107–113
Benson EE, Roubelakis-Angelakis KA (1994) Oxidative stress in recalcitrant tissue cultures of grapevine. Free Rad Biol Med 16:355–362
Bradford M (1976) A rapid and sensitive method for the quantitation of microprogram quantitaties of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Chiappetta A, Michelotti V, Fambrini M, Bruno L, Salvini M, Petrarulo M, Azmi A, van Onckelen H, Pugliesi C, Bitonti M (2006) Zeatin accumulation and misexpression of a class I knox gene are intimately linked in the epiphyllous response of the interspecific hybrid EMB-2 (Helianthus annuus × H. tuberosus). Planta 223:917–931
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Cui K, Xing G, Liu X, Wang Y (1999) Effect of hydrogen peroxide on somatic embryogenesis of Lycium barbarum L. Plant Sci 146:9–16
Cutler A, Saleem M, Wang H (1991) Cereal protoplast recalcitrance. In Vitro Cell Dev Biol 27:104–111
Dąbrowska G, Kaczmarek K, Skrzypek E, Szechyńska-Hebda M (2007) cDNA cloning and expression gene encoding manganese superoxide dismutase of Vicia faba ssp. minor in regenerated and non-regenerated explants. Adv Agric Sci Probl Issues 523:83–92
Daimon Y, Takabe K, Tasaka M (2003) The CUP-SHAPED COTYLEDON genes promote adventitious shoot formation on calli. Plant Cell Physiol 44:113–121
de Marco A, Roubelakis-Angelakis KA (1996a) Hydrogen peroxide plays a bivalent role in the regeneration of protoplasts. J Plant Physiol 149:109–114
de Marco A, Roubelakis-Angelakis KA (1996b) The complexity of enzymic control of hydrogen peroxide concentration may affect the regeneration potential of plant protoplasts. Plant Physiol 11:137–145
Desikan R, Mackerness AHS, Hancock JT, Neill SJ (2001) Regulation of the Arabidopsis transcriptome by oxidative stress. Plant Physiol 127:159–172
Desikan R, Hancock JT, Neill SJ (2003) Oxidative stress signalling. In: Hirt K, Shinozaki K (eds) Plant responses to abiotic stress, topics in current genetics, vol 4. Springer, Berlin, pp 129–149
Dodeman VL, Ducreux G (1996) Total protein pattern expression during induction and development of carrot somatic embryos. Plant Sci 120:57–69
Dong JZ, Dunstan DI (2000) Molecular biology of somatic embryogenesis in conifers. In: Jain SM, Minocha SC (eds) Molecular biology of woody plants, vol. 1. Kluwer Academic Publishers, Dordrecht, pp 51–87
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide and glutathione-associated mechanisms of acclimatory stress tolerance and signaling. Physiol Plant 100:241–254
Gara L, Pinto MC, Moliterni VMC, D’Egidio MG (2003) Redox regulation and storage processes during maturation in kernels of Triticum durum. J Exp Bot 54:249–258
Geisler M, Kleczkowski LA, Karpinski S (2006) A universal algorithm for genome-wide in silico identification of biologically significant gene promoter putative cis-regulatory-elements; identification of new elements for reactive oxygen species and sucrose signaling in Arabidopsis. Plant J 45:384–398
Gille L, Nohl H (2001) The ubiquinol/bc1 redox couple regulates mitochondrial oxygen radical formation. Arch Biochem Biophys 45:34–38
Gupta SD (2011) Role of free radicals and antioxidants in in vitro morphogenesis. In: Gupta SD (ed) Reactive oxygen species and antioxidants in higher plants. Science Publishers, USA, pp 229–247
Hake S, Smith HMS, Holtan H, Magnani E, Mele G, Ramirez J (2004) The role of knox genes in plant development. Annu Rev Cell Dev Biol 20:125–151
Halperin W (1995) In vitro embryogenesis: some historical issues and unresolved problems. In: Thorpe TA (ed) In vitro embryogenesis in plants. Kluwer Academic Publishers, Dordrecht, pp 1–16
Henzler T, Steudle E (2000) Transport and metabolic degradation of hydrogen peroxide in Chara corallina: model calculations and measurements with the pressure probe suggest transport of H2O2 across water channels. J Exp Bot 51:2053–2066
Hjortswang HI, Larsson AS, Bharathan G, Bozhkov PV, von Arnold S, Vahala T (2002) Knotted1-like homeobox genes of gymnosperm, Norway spruce, expressed during somatic embryogenesis. Plant Physiol Biochem 40:837–843
Hou L, Ullrich SE, Kleinhofs A, Stiff CM (1993) Improvement of anther culture methods for doubled haploid production in barley breeding. Plant Cell Rep 12:334–338
Hu G, Yalpani N, Briggs SP, Johal GS (1998) A porphyrin pathway impairment is responsible for the phenotype of a dominant disease lesion mimic mutant of maize. Plant Cell 10:1095–1105
Imani J, Tran Thi L, Langen G, Arnholdt-Schmitt B, Roy S, Lein C, Kumar A, Neumann KH (2001) Somatic embryogenesis and DNA organization of genomes from selected Daucus species. Plant Cell Rep 20:537–541
Inzé D, Montagu MV (1995) Oxidative stress in plants. Curr Opin Biotechnol 6:153–158
Ishikawa T, Takeda T, Shigeoka S, Hirayama O, Mitsunaga T (1993) Hydrogen peroxide generation in organelles of Euglena gracilis. Phytochemistry 33:1297–1299
Jevremović S, Petrić M, Zivković S, Trifunović M, Subotić A (2010) Superoxide dismutase activity and isoenzyme profiles in bulbs of snake’s head fritillary in response to cold treatment. Arch Biol Sci 62:553–558
Jiménez VM, Bangerth F (2001) Endogenous hormone concentrations and embryogenic callus development in wheat. Plant Cell Tiss Org Cult 67:37–46
Kairong C, Gengsheng X, Xinmin L, Gengmei X, Yafu W (1999) Effect of hydrogen peroxide on somatic embryogenesis of Lycium barbanum L. Plant Sci 146:9–16
Karpinski S, Reynolds H, Karpinska B, Wingsle G, Creissen G, Mullineaux P (1999) Systemic signalling and acclimatisation in response to excess excitation energy in Arabidopsis. Science 284:654–657
Krikorian AD, Simola LK (1999) Totipotency, somatic embryogenesis, and Harry Waris (1893–1973). Physiol Plant 105:348–355
Loschiavo F, Pitto L, Giuliano G, Torti G, Nuti-Ronchi V, Marazziti D, Vergara R, Orselli S, Terzi M (1989) DNA methylation of embryogenic carrot cell cultures and its variation as caused by mutation, differentiation, hormones and hypomethylating drugs. Theor Appl Genet 77:325–331
McCord JM, Friodovich I (1969) Superoxide dismutase. An enzyme function for erythrocuperein (hemocuperein). J Biol Chem 244:6049–6055
Mehlhorn H, Lelandais M, Korth HG, Foyer CH (1996) Ascorbate is the natural substrate for plant peroxidases. FEBS Lett 378:203–206
Merkle SA, Parrott WA, Flinn BS (1995) Morphogenic aspects of somatic embryogenesis. In: Thorpe TA (ed) In vitro embryogenesis in plants. Kluwer Academic Publishers, Dordrecht, pp 155–203
Mittler R, Zilinskas B (1991) Molecular cloning and nucleotide sequence analysis of a cDNA encoding pea cytosolic ascorbate peroxidase. FEBS Lett 289:257–259
Möller IM (2001) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu Rev Plant Physiol Plant Mol Biol 52:561–591
Morré DM, Lenaz G, Morré DJ (2000) Surface oxidase and oxidative stress propagation in aging. J Exp Biol 203:1513–1521
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Neill S, Desikan R, Hancock J (2002) Hydrogen peroxide signalling. Curr Opin Plant Biol 5:388–395
Nomura K, Komamine A (1995) Physiological and biological aspects of somatic embryogenesis. In: Thorpe TA (ed) In vitro embryogenesis in plants. Kluwer Academic Publishers, Dordrecht, pp 249–266
Obert B, Dedicova B, Hricova A, Samaj J, Pretova A (2004) Flax anther culture: effect of genotype, cold treatment and media. Plant Cell Tiss Org Cult 79:233–238
Okuda T, Masuda Y, Yamanaka A, Sagisaka S (1991) Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol 97:1265–1267
Padmanabhan K, Cantliffe DJ, Koch KE (2001) Auxin-regulated gene expression and embryogenic competence in callus cultures of sweet potato, Ipomoea batatas (L.). Plant Cell Rep 20:187–192
Papadakis AK, Roubelakis-Angelakis A (1999) The generation of active oxygen species differs in tobacco and grapevine mesophyll protoplasts. Plant Physiol 121:197–205
Papadakis AK, ChI Siminis, Roubelakis-Angelakis KA (2001) Reduced activity of antioxidant machinery is correlated with suppression of totipotency in plant protoplasts. Plant Physiol 126:434–444
Pasternak TP, Prinsen E, Ayaydin F, Miskolczi P, Potters G, Asard H, van Onckelen HA, Dudits D, Feher A (2002) The role of auxin, pH, and stress in the activation of embryogenic cell division in leaf protoplast-derived cells of Alfalfa. Plant Physiol 129:1807–1819
Pellegrineschi A, Brito RM, McLean S, Hoisington D (2004) Effect of 2, 4-dichlorophenoxyacetic acid and NaCl on the establishment of callus and plant regeneration in durum and bread wheat. Plant Cell Tiss Org Cult 77:245–250
Pellinen RI, Minna-Sisko K, Tauriainen AA, Palva ET, Kangasjärvi J (2002) Hydrogen peroxide activates cell death and defense gene expression in birch. Plant Physiol 130:549–560
Prasad TK, Anderson MD, Stewart CR (1994) Acclimation, hydrogen peroxide, and abscisic acid protect mitochondria against irreversible chilling injury in maize seedlings. Plant Physiol 105:619–627
Purvis AC, Shewfelt RL, Gegogeine JW (1995) Superoxide production by mitochondria isolated from green bell pepper fruit. Physiol Plant 94:743–749
Qiusheng Z, Bao J, Likun L, Xianhua X (2005) Effects of antioxidants on the plant regeneration and GUS expressive frequency of peanut (Arachis hypogaea) explants by Agrobacterium tumefaciens. Plant Cell Tiss Org Cult 81:83–90
Rai MK, Kalia RK, Singh R, Gangola MP, Dhawan AK (2011) Developing stress tolerant plants through in vitro selection—an overview of the recent progress. Env Exp Bot 71:89–98
Rao KS (1996) Embryogenesis in flowering plants—recent approaches and prospects. J Biosci 21:827–841
Reilly K, Han Y, Tohme J, Beeching JR (2001) Isolation and characterisation of a cassava catalase expressed during post-harvest physiological deterioration. Biochim Biophys Acta 1518:317–323
Rhoads DM, Umbach AL, Subbaiah CC, Siedow JN (2006) Mitochondrial reactive oxygen species. Contribution to oxidative stress and interorganellar signalling. Plant Physiol 141:357–366
Rose RJ, Mantiri FR, Kurdyukov S, Chen SK, Wang XD, Nolan KE, Sheahan MB (2010) Developmental biology of somatic embryogenesis. Plant Dev Biol Biotech Perspect 1:3–26
Roubelakis-Angelakis KA (1993) An assessment of possible factors contributing to recalcitrance of plant protoplasts. In: Roubelakis-Angelakis KA, Tranh TVK (eds) Morphogenesis in plants: molecular approaches. Plenum Publishing, New York, pp 201–219
Sambrook J, Fritsh EF, Maniatis T (1989) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor
Scandalios JG (1993) Oxygen stress and superoxide dismutases. Plant Physiol 100:7–12
Schmidt EDL, Guzzo F, Toonen MAJ, De Vries SC (1997) A leucine-rich repeat containing receptor-like kinase marks somatic plant cell cultures. Development 124:2049–2062
Schrader S, Kaldenhoff R, Richter G (1997) Expression of novel genes during somatic embryogenesis of suspension cultured carrot cells (Daucus carota). J Plant Physiol 150:63–68
Schryer PA, Lu Q, Vandenberg A, Bett KE (2005) Rapid regeneration of Phaseolus angustissimus and P. vulgaris from very young zygotic embryos. Plant Cell Tiss Org Cult 83:67–74
Sinha S, Gupta M, Chandra P (1997) Oxidative stress induced by iron in Hydrilla verticillata (l.f.) Royle: response of antioxidants. Ecotoxicol Environ Saf 38:286–291
Ślesak I, Libik M, Karpinska B, Karpinski S, Miszalski Z (2007) The role of hydrogen peroxide in regulation of plant metabolism and cellular signalling in response to environmental stresses. Acta Biochem Pol 54:39–50
Somleva MN, Schmidt EDL, De Vries SC (2000) Embryogenic cells in Dactylis glomerata L. (Poaceae) explants identified by cell tracking and by SERK expression. Plant Cell Rep 19:718–726
Szechynska-Hebda M, Skrzypek E, Dabrowska G, Koscielniak J, Filek M, Wedzony M (2007) The role of oxidative stress induced by growth regulators in the regeneration process of wheat. Acta Physiol Plant 29:327–337
Szechyńska-Hebda M, Kruk J, Gorecka M, Karpinska B, Karpiński S (2010) Evidence for light wavelength-specific systemic photoelectrophysiological signalling and cellular light memory of excess light episode in Arabidopsis. Plant Cell 22:1–18
Tamás C, Szűcs P, Rakszegi M, Tamás L, Bedő Z (2004) Effect of combined changes in culture medium and incubation conditions on the regeneration from immature embryos of elite varieties of winter wheat. Plant Cell Tiss Org Cult 79:39–44
Toonen MAJ, De Vries SC (1996) Initiation of somatic embryos from single cells. In: Wang TL, Cuming A (eds) Embryogenesis: the generation of a plant. Bios Scientific Publishers, Oxford, pp 173–189
Touraev A, Vicente O, Heberle-Bors E (1997) Initiation of microspore embryogenesis by stress. Trends Plant Sci 2:297–302
Vansuyt G, Lopez F, Inzé D, Briat JF, Fourcroy P (1997) Iron triggers a rapid induction of ascorbate peroxidase gene expression in Brassica napus. FEBS Lett 410:195–200
Veljovic-Jovanovic S, Noctor G, Foyer CH (2002) Are leaf hydrogen peroxide concentrations commonly overestimated? The potential influence of artefactual interference by tissue phenolics and ascorbate. Plant Physiol Biochem 40:501–507
Vertucci CW (1989) The kinetics of seed imbibitions: controlling factors and relevance to seedling vigor. In: Stanwood PC, McDonald MB (eds) Seed moisture, CSSA Special Publication 14. Crop Science Society of America, Inc., Madison, pp 93–115
Vranová E, Inzé D, Van Breusegem F (2002) Signal transduction during oxidative stress. J Exp Bot 53:1227–1236
Willekens H, Chamnongpol S, Davey M, Schraudner M, Langebartels C, van Montagu M, Inzé D, van Camp W (1997) Catalase is a sink for H2O2 and indispensable for stress defence in C3 plants. EMBO J 16:4806–4816
Wu G, Wilen RW, Robertson AJ, Gusta LV (1999) Isolation, chromosomal localization, and differential expression of mitochondrial manganese superoxide dismutase, and chloroplastic copper/zinc superoxide dismutase genes in wheat. Plant Physiol 120:513–520
Xynias IN, Zamani IA, Gouli-Vavdinoudi E (2001) Effect of cold pre-treatment and incubation temperature on bread wheat (Triticum aestivum L.) anther culture. Cereal Res Com 29:331–338
Yoshimura K, Yabuta Y, Ishikawa T, Shigeoka S (2000) Expression of spinach ascorbate peroxidase. Plant Physiol 123:223–233
Zavattieri MA, Frederico AM, Lima M, Sabino R, Arnhold-Schmitt B (2010) Induction of somatic embryogenesis as an example of stress-related plant reactions. Electr J Biotech 13:1–9. doi:10.2225/vol13-issue1-fulltext-4
Zhang S, Wong L, Meng L, Lemaux PG (2002) Similarity of expression patterns of knotted1 and zmlec1 during somatic and zygotic embryogenesis in maize (Zea mays L.). Planta 215:191–194
Zhang S, Han S, Yang W, Wei H, Zhang M, Qi Li (2010) Changes in H2O2 content and antioxidant enzyme gene expression during the somatic embryogenesis of Larix leptolepis. Plant Cell Tiss Org Cult 100:21–29
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W. Filek.
Rights and permissions
About this article
Cite this article
Szechyńska-Hebda, M., Skrzypek, E., Dąbrowska, G. et al. The effect of endogenous hydrogen peroxide induced by cold treatment in the improvement of tissue regeneration efficiency. Acta Physiol Plant 34, 547–560 (2012). https://doi.org/10.1007/s11738-011-0852-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-011-0852-3