
Abstract
The effects of glycosylation followed by limited hydrolysis on the structural and functional properties of soybean protein isolate (SPI) were investigated. SPI was glycosylated by transglutaminase (TGase) in the presence of oligochitosan to generate a glycosylated soybean protein isolate (GSPI) and then hydrolyzed using Alcalase to obtain hydrolysates with degree of hydrolysis (DH) of 1%, 2% and 4%. Structural properties were characterized using estimate of molecular weight distribution and circular dichroism (CD) spectra. Electrophoresis analysis revealed that GSPI contains peptide polymers, and its hydrolysates exhibit broad molecular weight distributions. CD analysis indicated more flexible structure for GSPI compared to SPI, and the following limited hydrolysis of GSPI induces even more flexible structure on its hydrolysates. GSPI showed significant increase in the apparent viscosity at the shear rate of 0.1–10/s. Emulsion stability of GSPI also increased by 33.9% than that of SPI. Hydrolysates from GSPI, especially for DH of 4%, showed significantly improved emulsifying activity index, increasing by 39.2% compared to SPI. The hydrolysates also showed notably enhanced antioxidant activities, namely, DPPH radical-scavenging ability and reducing power. These results revealed that TGase-induced glycosylation and further limited hydrolysis may have potential for creating food ingredients with improved functional properties.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Soybean protein isolate (SPI) is a prime source of high-quality proteins and can be used as a substitute of animal proteins despite its deficiency in the essential amino acid methionine. SPIs have less cost of natural resources and some good functional properties [1]. There has been increasing interest in SPI as a nutritional and functional ingredient in processed foods. Functional properties of proteins depend on their molecular size, structure and specific amino acid sequence. Moreover, functional properties of SPI can be enhanced by various methods, such as enzymatic hydrolysis, glycosylation and physical modifications [2,3,4].
Enzymatic hydrolysis is an effective enzymatic modification method to improve the functional properties of SPIs. Enzymatic hydrolysis is relatively easy to control with mild reaction conditions and producing minimal by-products [5]. And the obtained hydrolysates show two major modifications: smaller molecular size and less secondary structure [6]. To obtain desirable functional properties in SPI hydrolysates, enzymatic hydrolysis is carried out under well controlled conditions to a specified degree of hydrolysis (DH). A limited hydrolysis (generally low DH) has been studied to improve the interfacial and foaming characteristics of soybean glycinin [7], produce higher gel hardness of SPI [8], and lead to a decrease in surface hydrophobicity of lentil proteins [9].
Protein glycosylation has also been shown as a promising way to modify the functional properties of proteins via covalently conjugating saccharide groups to the protein molecule. It may be possible to combine their individual properties and thus generate food ingredients with a wider range of functional properties. Maillard-type glycation is a well-known and widely used protein glycosylation. Recently, transglutaminase (TGase, EC 2.3.2.13) induced glycosylation has been used for modifying the functional properties of food proteins. Originally, TGase-induced protein cross-linking, through ɛ-(γ-glutamyl) lysine isopeptide bonds between glutamine and lysine residues of the proteins [10], was used to focus on improving the emulsifying [11] and elastic [12] properties of food proteins. Until recently, TGase can also be found to conjugate the compound (i.e. acyl acceptor) into the glutamine residues (i.e. acyl donor) of the proteins in the presence of a primary amine compound [10], and has been successfully applied to glycosylate of food proteins. TGase induced glycosylation of casein or fish gelatin has been carried out in the presence of monosaccharide (glucosamine), and conferred the product improved emulsifying properties [13], or an improved antioxidant and antimicrobial activity [14].
In a recent study, Song and Zhao [15] demonstrated that TGase can be used to induce the conjugation of an oligosaccharide (oligochitosan) into SPI molecules [15]. The prepared product, a glycosylated and cross-linked soybean protein isolate, exhibited some desirable properties, especially with an improved water binding capacity and emulsion stability of SPI [15]. However, no information is currently available in the literature on the combined contributions of TGase induced glycosylation and hydrolysis with Alcalase on the structural and functional properties of SPI.
The objective of the present work is to characterize the secondary structure, emulsifying, antioxidant and viscosity properties of TGase-induced glycosylation of SPI followed by Alcalase hydrolysis, providing useful information regarding its potential applications as protein ingredients.
Materials and methods
Materials and chemicals
SPI was extracted as previously described [15] using defatted soybean flour purchased from Harbin Binxian Yuwang Vegetable Protein Co., Ltd. (Harbin, Heilongjiang, China), and protein content of 90.3% (w/w) determined by the Kjeldahl method. Oligochitosan with a declared deacetylation degree of about 90% and an average molecular weight of 1 kDa was purchased from Zhejiang Golden-Shell Biochemical Co. (Hangzhou, Zhejiang, China). TGase was purchased from Jiangsu Yiming Fine Chemical Industry Co., Ltd. (Qinxing, Jiangsu, China) with a declared activity of 100 U/g. Alcalase (2.94 × 102 U/mg) was obtained from Novozymes (Bagsvaerd, Denmark). Acrylamide and glycine were purchased from Amresco, Inc. (Solon, OH, USA). Bis-acrylamide and 1,1-diphenyl-2-picryl-hydrazyl (DPPH), pepsin P7000 and trypsin T7409 were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). The highly purified water used was prepared with Milli-Q PLUS (Millipore Corporation, New York, NY, USA). Other chemicals used were of analytical grade.
Preparation of a glycosylated soybean protein isolate (GSPI) and Alcalase catalyzed protein hydrolysates
GSPI was prepared as our previously reported method [15]. The stock SPI dispersion (heated to 90 °C for 10 min, and then cooled to 37 °C) was mixed with the oligochitosan solution with a 1:3 molar ratio to make a final soybean protein content of 4% (w/v) with a pH value of 7.5. TGase was added into the mixed solution at the level of 10 U/g protein. The reaction was conducted at 37 °C for 3 h and then deactivated the enzyme at 85 °C for 15 min. The mixture was treated with isoelectric precipitation and then centrifuged (4000×g at 4 °C) for 15 min. The collected precipitates were washed twice by water of pH 4.5 and lyophilized to obtain GSPI.
A GSPI dispersion (3.5%, w/v) was adjusted to pH 8.5 with 2 mol/L NaOH and incubated at 60 °C for about 10 min. Then, the protease Alcalase was added, at the ratio of enzyme to substrate of 0.1% (w/w). The mixtures were incubated at 60 °C to start the enzymatic hydrolysis reaction. The pH value of the mixture maintained constant during hydrolysis by addition of 1 mol/L NaOH using pH-stat method. Degree of hydrolysis (DH) was used as an indication of enzymatic hydrolysis, and DH was determined according to the method of Adler-Nissen [16]
where B is the amount of alkali consumed (mL), Nb is the normality of alkali, Mp is the mass of the substrate (protein in grams, % N × 6.25), 1/α is the calibration factors for pH-stat, and htot is the number of peptide bonds, that is, 7.8 mequiv/g protein [16]. When DH values had been reached (DHs of 1%, 2% and 4%), the hydrolysis reaction was terminated by heating at 85 °C for 15 min with occasional mixing. The samples were cooled and the pH values of samples were then adjusted to 7.0 in distilled water, and then lyophilized. Crude protein content of the prepared powder was determined by the Kjeldahl method.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE)
SDS–PAGE was performed using 12% separating gels and 4% stacking gels. Electrophoresis was carried out at 120 V for separation. After the electrophoresis, the gels were removed, fixed, then stained with the staining solution (Coosmasie brilliant blue 0.05%, ethanol 25%, acetic acid 10%) for protein. Protein staining was destained in methanol–water–acetic acid solution (5:5:1, v/v).
Far-UV circular dichroism (CD)
Circular dichroism spectra of five samples (herein SPI, GSPI without hydrolysis, GSPI with DH 1%, 2%, and 4% samples) from 190 to 240 nm were carried out to estimate the secondary structure using a Jasco J-815 CD spectrometer (Jasco Corporation, Tokyo, Japan). A 0.1 cm path length cuvette was used to contain the sample solution of 0.25 mg/mL (in 10 mM phosphate buffer). The data of secondary structures (α-helix, β-sheet, β-turn and random coil) was calculated using the protein secondary structure estimation software.
Evaluation of antioxidant properties
Determination of DPPH radical scavenging ability
The scavenging effect of five samples on DPPH free radical was measured according the procedure reported method [17], with minor modifications. Two milliliters of the samples (1.5 mg/mL) were mixed with 2.0 mL of DPPH in anhydrous ethanol and mixed vigorously. After incubating in dark for 30 min at room temperature, the absorbance of the resulting solutions was measured at 517 nm with the UV–visible spectrophotometer (model UV-5100, Shanghai Metash Instrument Co. Ltd., China). The control and blank were conducted in the same manner, where control used anhydrous ethanol instead of sample solution and blank used anhydrous ethanol rather than DPPH solution. Ascorbic acid (0.01 mg/mL) was used as a positive control. DPPH radical scavenging activity of the peptides was calculated according to the following equation:
where As, Ab and Ac represent the absorbance of sample, blank and control, respectively.
Reducing power
The reducing power of five samples was measured as per the reported method [18] with minor modifications. Two milliliters of samples (2.5 mg/mL) were mixed with 2.0 mL of 0.2 mol/L phosphate buffer (pH 6.6) and 2.0 mL of 1% (w/v) potassium ferricyanide. The mixture was incubated at 50 °C for 20 min. An aliquot (2.0 mL) of 10% trichloroacetic acid (TCA) were added to stop the reaction, followed by centrifugation at 4000×g for 10 min. The upper layer of 2.0 mL was mixed with 2.0 mL deionized water and 0.4 mL of 0.1% FeCl3. After incubating at 50 °C for 10 min, the absorbance of the resulting solution was measured at 700 nm. Deionized water was used as blank instead of sample solution. A stronger absorbance of the mixture indicated an increasing reducing power. Ascorbic acid (0.01 mg/mL) was used as a positive control.
Hydroxyl radical scavenging ability
The scavenging activity of five samples on hydroxyl radical was assessed as per the method described earlier [19] with minor modification. Two milliliters of samples (2.5 mg/mL) were mixed with FeSO4 (6.0 mmol/L) and H2O2 (6.0 mmol/L) and the mixture was incubated for 10 min. An aliquot (2.0 mL) of salicylic acid (6.0 mmol/L) was added and the mixture was incubated for 30 min. The absorbance of the resulting solution was measured at 510 nm. Distilled water was used for the blank (instead of salicylic acid solution) and the control (instead of sample solution). Ascorbic acid (0.01 mg/mL) was used as a positive control. Scavenging the hydroxyl radical activity of the peptides was calculated according to the following equation:
where As, Ab and Ac represent the absorbance of sample, blank and control, respectively.
Determination of functional properties
Emulsifying properties
Emulsifying activity index (EAI) and emulsion stability index (ESI) of 5 samples were determined by the turbidimetric method [20]. Emulsions were prepared by using 75 mL of protein solution of 1 mg/mL at pH 7.0 and 25 mL of refined soybean oil, homogenized at 12,000 rpm for 1 min (Model JS25, Electromechanical equipment manufacturing Co., Ltd., Jiangsu, China). Emulsion (50 µL) was pipetted from the bottom of the container into 5 mL of 0.1% (w/v) sodium dodecyl sulfate (SDS) solution immediately (0 min) and 10 min after homogenization. Absorbance of the deaerated emulsion (A0 or A10) was measured at 500 nm (UV-5100 spectrophotometer, Shanghai Metash Instruments Co., Ltd., Shanghai, China). EAI and ESI were calculated as followed:
where C is the concentration of aqueous phase protein (10 kg/L), Φ is the volumetric fraction of oil (0.25) and the value of the dilution equals to 101.
In vitro digestibility
In vitro digestibility of five samples was evaluated according to the earlier reported methods by Marciniak-Darmochwall and Kostyra [21] and Yin et al. [22]. Pepsin or pepsin–trypsin was used to hydrolyze five samples to simulate digestion in vitro. For one step hydrolysis, protein dispersions of five samples (1%, w/v) were hydrolyzed by pepsin at pH 2.0 with the enzyme addition level of 20 mg/g protein. The hydrolysis was conducted at 37 °C for 2 h and then reaction was stopped by addition an equal volume of 20% (w/v) trichloroacetic acid (TCA). The mixture was then centrifuged (10,000×g for 20 min at 4 °C) for 15 min. For two step hydrolysis, the starting solutions were prepared and treated as one step hydrolysis. After 1-h incubation at 37 °C, the pepsin was deactivated by heating at 90 °C for 5 min and the samples were lyophilized. Thereafter, the obtained samples further hydrolyzed by trypsin (60 mg/g protein) at 37 °C for 1 h. The termination process and centrifugation treatment of two step hydrolysis were conducted in the same manner as one step hydrolysis. TCA-soluble nitrogen released into the supernatants was measured absorbance at 280 nm after proper dilution, and the data was used to express the digestibility of the samples.
Apparent viscosity
Protein dispersions of five samples (3.5%, w/w) were prepared in deionized water at pH 7.0. The dispersions were allowed to equilibrate overnight at 4 °C and stirred for 30 min at room temperature before analysis. Small-amplitude oscillatory shear tests were performed with Kinexus Pro + Super rotational rheometer (Malvern Instruments, Malvern, USA) using a cone-and-plate probe (60 mm diameter, 1° cone angle). The dispersion was loaded onto the rheometer and relaxed the stress for about 5 min to reach thermal equilibrium (25 °C) before the measurement. The shear rate was controlled between 0.1 and 10/s.
Statistical analysis
All experiments or analyses were carried out three times. All reported data were expressed as means or means ± standard deviations. Differences between the means of multiple groups were analyzed by one-way analysis of variance (ANOVA) with Duncan′s multiple range tests. SPSS 13.0 software (SPSS Inc., Chicago, IL, USA) and MS Excel 2003 software (Microsoft Corporation, Redmond, WA, USA) were used to analyze and report the results.
Results and discussion
Molecular weight distribution of GSPI and its low-DH Alcalase hydrolysates by SDS–PAGE
Electrophoresis analysis was used to visually identify and estimate the molecular weight (size) distribution of the protein. Figure 1 shows the SDS–PAGE profile of GSPI and its low-DH Alcalase hydrolysates. SDS–PAGE analysis showed that the GSPI contains protein polymers (Fig. 1, Lane 2), which does not enter the stacking or separating gel. This observation is consistent with our earlier study [15]. It is also seen that the followed hydrolysis of GSPI with Alcalase resulted in a rapid disappearance of bands corresponding to protein polymers even through at low levels of hydrolysis (Fig. 1, Lane 3–5) and along with the appearance of the substantially broad band from the top of the stacking gel to bottom of the separating gel. During the course of hydrolysis, as the DH increases from 1 to 4%, the content of larger peptides was decreased, corresponding to the appearance of new peptides in the lower range of molecular weight. Hydrolysates contain a molecular weight distribution of peptides ranging from large (> 100 kDa) to small (< 10 kDa) molecular size. These results indicated that enzymatic treatment with Alcalase resulted in the hydrolysis of most of the GSPI proteins, producing hydrolysates with broad molecular weight distributions.

SDS–PAGE profiles of GSPI generated from SPI by TGase in presence of oligochitosan and its Alcalase hydrolysates with DH of 1%–4%. Lane M represents standard protein markers whereas lane 1–5 represent SPI, GSPI, GSPI hydrolysate with 1% DH, GSPI hydrolysate with 2% DH, and GSPI hydrolysate with 4% DH, respectively
Secondary structural characteristics of GSPI and its low-DH Alcalase hydrolysates
A deeper understanding of secondary structure of GSPI and its hydrolysates was obtained using far-UV CD spectra (Table 1). SPI consists mainly of globulins, whose structure is characterized in an ordered secondary structure [23]. Compared to SPI, the prepared GSPI has less α-helix and β-structures (Table 1). After hydrolysis treatment, the secondary structure of GSPI has significantly changed. With the increase of DH in the range of 1%–4%, the content of α-helical and β-structure for GSPI decreases, whereas random coil structure of GSPI increases. For the hydrolysate with DH of 4%, the β-sheet structure in the GSPI hydrolysates drops from 46.4 to 26.4%, whereas the contents of random coil and β-turn structure show a clear rise from 39.6 to 55.3% and from 10.0 to 18.2%, respectively (Table 1). It has been reported that β-sheet structures are relatively stable, whereas α-helix, β-turn, and random coil structures are relatively flexible and open [24]. This indicates that enzyme hydrolysis confers GSPI a more flexible secondary structure. Similar results are found in the previous study on the secondary structure of sunflower protein isolate hydrolysates, which becomes a more flexible form after treatment with Alcalase [25].
Antioxidants activity of GSPI and its hydrolysates
DPPH radical scavenging ability
DPPH analysis is based on the reduction of alcoholic DPPH solution in the presence of antioxidant (H-donating) due to the formation of DPPH-H (non-radical form) by the reaction [26]. DPPH radical scavenging ability of GSPI and its hydrolysates are shown in Fig. 2a. GSPI exhibits higher DPPH radical scavenging ability (35.1%) than that of SPI (27.7%), similar to the Maillard-type glycation that improves the antioxidant activity of egg white lysozyme [27]. The DPPH radical scavenging ability of GSPI is also enhanced by the following hydrolysis, and further enhanced with the increase of DH. Hydrolysates with DH of 4% show the strongest DPPH radical scavenging ability (59.0%). Conjugated hydrophilic saccharide into GSPI could donate more electron donors, which could react with free radicals to convert them to stable products and terminate the radical chain reactions. In addition, smaller molecular weight of the peptides obtained during Alcalase hydrolysis also contribute to the improved antioxidant activity [28]. These results indicated both glycosylation and the following hydrolysis are beneficial to improve the DPPH radical scavenging ability of SPI.

Antioxidant activity (at pH 7.0) of GSPI generated from SPI by TGase in presence of oligochitosan and its Alcalase hydrolysates with DH of 1%–4%: a DPPH radical scavenging ability, b reducing power, and c hydroxyl radical scavenging ability
Reducing power
Reducing power assay is an alternative way to describe the antioxidant activity of compounds, which depends on the changes of absorbance during the reaction of a ferric tripyridyltriazine complex to the ferrous form by an antioxidant at a low pH. The increase in absorbance directly correlates to better ability to donate electron by antioxidants present in the reaction mixture [29]. The results of reducing power of GSPI and its hydrolysates are presented in Fig. 2b. GSPI shows higher antioxidant activity (0.282) as compared to that of SPI (0.240). GSPI hydrolysates show enhanced reducing power (0.344–0.353), and there is no significant difference in the three hydrolysates. These results reveal that GSPI and its hydrolysates can possess the properties as effective electron donors.
Hydroxyl radical scavenging ability
Hydroxyl radical is the most reactive free radical among the oxygen free radicals, which can cause severe damage to adjacent biomolecules [29]. It is meaningful to scavenge the hydroxyl radical to prevent putrefaction of food and occurrence of various diseases. Hydroxyl radical scavenging ability was used to estimate the scavenging activity of free hydroxyl radicals like hydrogen peroxide in the presence of antioxidants. Figure 2c represents the hydroxyl radical scavenging ability of GSPI and its hydrolysates. GSPI exhibits higher inhibitory activity (41.6%) against hydroxyl radicals as compared to that of SPI (36.2%). The hydroxyl radical scavenging activitiy values of three GSPI hydrolysates (in the order of GSPI-1%, GSPI-2% and GSPI-4%) are 30.1%, 29.7% and 38.4%, respectively. GSPI hydrolysates with DH of 4% show the most effective inhibitory hydroxyl radical activity (38.4%) in hydrolysates, which is similar to SPI (36.2%). These results suggest that glycosylation of SPI has positive effect on the hydroxyl radical scavenging activity. Increasing DH of GSPI can also have the potential to increase the hydroxyl radical scavenging activity.
Functional and physicochemical characteristic of GSPI and its hydrolysates
Emulsifying properties
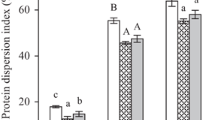
Emulsifying activity index (EAI) and emulsion stability index (ESI) of GSPI and its hydrolysates are shown in Fig. 3a. EAI value of GSPI decreases in comparison with SPI. After hydrolysis, a notable increase in EAI is observed, and the highest value (~ 66 m2/g protein) is exhibited for GSPI hydrolysates with DH of 4% that is higher than that of SPI (~ 48 m2/g protein). These results suggest that the following hydrolysis treatment of GSPI improves EAI. The reason may be that the hydrolysis makes the products to reach an optimal molecular weight distribution [9]. It is consistent with the previous study that the limited hydrolysis improves emulsifying properties of arachin protein [30] and whey protein [31]. For emulsion stability, GSPI exhibits significantly higher ESI than that of SPI, while its hydrolysates show a decreased ESI (p < 0.05). The higher ESI in GSPI is due to that saccharide molecules adsorbed in the layer can slow down the processes of droplet rise and cream separation [32, 33]. While a decreased ESI in hydrolysates of GSPI can be caused by small peptides that are less efficient in reducing the interfacial tension, because they cannot unfold and reorient at the interface [34]. These results show that the glycosylation significantly improves the ESI of SPI, while the following hydrolysis significantly enhances the EAI of SPI.

Emulsifying property (a) and in vitro digestibility (b) of GSPI generated from SPI by TGase in presence of oligochitosan and its Alcalase hydrolysates with DH of 1%–4% as well as apparent viscosity (c) of their aqueous dispersions
In vitro digestibility
In vitro protein digestibility of GSPI and its hydrolysates is reflected by the absorbance at 280 nm of the TCA-soluble nitrogen released during the enzymatic digestion as shown in Fig. 3b. The absorbance of GSPI digestion products is lower in both pepsin (one step hydrolysis) and pepsin–trypsin (two steps hydrolysis) in comparison with SPI (0.313 vs. 0.365 and 0.520 vs. 0.588, respectively). These results indicated that GSPI is more resistant to the hydrolysis by pepsin or trypsin than SPI. Even though binding of saccharides into protein molecules (resulting in partial unfolding structures) is beneficial to the hydrolysis of the conjugates [35], cross-linking of food proteins can result in reduced tryptic digestibility due to a reduction in chain flexibility and formation of isopeptide bonds [36]. Hydrolysates from GSPI possess shorter chain and more flexible structure, leading to a low resistance to pepsin or trypsin attack [37]. As expected, GSPI hydrolysates are strongly hydrolyzed during pepsin or pepsin–trypsin hydrolysis, and in vitro digestibility of the hydrolysates increases with increasing DH.
Apparent viscosity
Flow behaviors of aqueous dispersions of GSPI and its hydrolysates are shown in Fig. 3c. GSPI exhibits significantly higher apparent viscosity as compared to SPI. This can be attributed to that TGase induced cross-linking of protein molecules increases the molecular weights and molecular volume of the SPI. Water hydration of the attached saccharides (oligochitosan) also contributes to the increase of the apparent viscosity of GSPI [38]. In general, low apparent viscosity is observed in proteins when their molecular mass is reduced by proteolysis [39]. As shown in Fig. 3c, apparent viscosity of suspension of hydrolysates from GSPI significantly decreases. In addition, the apparent viscosity of all suspensions decrease with increasing shearing rate, i.e. all five SPI samples behave shear thinning behavior during shear treatment. A high viscosity is desirable for food processing like thickening soup and production of sausage, and a low viscosity has been considered beneficial in many manufacturing operations.
GSPI and hydrolysates from GSPI exhibit some functional properties that are different from SPI, especially on the improved rheological property and antioxidant activities. These improved properties indicate their potential use as protein ingredients for food processing. For example, GSPI might be applicable to some processed foods for thickening agent and stabiliser. The hydrolysates from GSPI might be as a good candidate for producing many functional foods, such as antioxidant peptide. More practical impact of GSPI and its hydrolysates on quality attributes of the formulated foods is still intriguing and needs to be further investigated in future.
Conclusions
A glycosylated soybean protein (GSPI) generated by TGase in the presence of oligochitosan exhibits significantly increased apparent viscosity and emulsion stability. Low degree of hydrolysis (DH) Alcalase hydrolysates from GSPI significantly improves emulsifying, in vitro digestibility and antioxidant properties at neural pH and generates products with a broad molecular weight distribution and flexible structure. Thus, this demonstrates that desirable functional properties of SPI can be obtained by TGase-induced glycosylation and further limited hydrolysis.
References
V.P. Ruíz-Henestrosa, C. Carrera Sánchez, M. del Mar Yust, J. Pedroche, F. Millán, J.M. Rodríguez Patino, Limited enzymatic hydrolysis can improve the interfacial and foaming characteristics of β-conglycinin. J. Agric. Food Chem. 55, 1536–1545 (2007)
D. Panyam, A. Kilara, Enhancing the functionality of food proteins by enzymatic modification. Trends Food Sci. Technol. 7, 120–125 (1996)
L. Mirmoghtadaie, S.S. Aliabadi, S.M. Hosseini, Recent approaches in physical modification of protein functionality. Food Chem. 199, 619–627 (2016)
J.H. Liu, Q.M. Ru, Y.T. Ding, Glycation a promising method for food protein modification: physicochemical properties and structure, a review. Food Res. Int. 49, 170–183 (2012)
A. Mannheim, M. Cheryan, Enzyme-modified proteins from corn gluten meal: preparation and functional properties. J. Am. Oil Chem. Soc. 69, 1163 (1992)
K. Bandyopadhyay, S. Ghosh, Preparation and characterization of papain-modified sesame (Sesamum indicum L.) protein isolates. J. Agric. Food Chem. 50, 6854–6857 (2002)
V.P. Ruíz-Henestrosa, C.C. Sánchez, J.J. Pedroche, F. Millán, J.M.R. Patino, Improving the functional properties of soy glycinin by enzymatic treatment. Adsorption and foaming characteristics. Food Hydrocol. 23, 377–386 (2009)
Y. Hou, X.H. Zhao, Limited hydrolysis of two soybean protein products with trypsin or neutrase and the impacts on their solubility, gelation and fat absorption capacity. Biotechnol. 10, 190–196 (2011)
N.A. Avramenko, N.H. Low, M.T. Nickerson, The effects of limited enzymatic hydrolysis on the physicochemical and emulsifying properties of a lentil protein isolate. Food Res. Int. 51, 162–169 (2013)
M. Kieliszek, A. Misiewicz, Microbial transglutaminase and its application in the food industry. A review. Folia Microbiol. (Praha) 59, 241–250 (2014)
C. Qiu, X. Hu, L. Li, X. Yang, M. Zhao, J. Ren, Effect of transglutaminase cross-linking on the conformational and emulsifying properties of peanut arachin and conarachin fractions. Eur. Food Res. Technol. 243, 913–920 (2017)
M. Jin, Q. Zhong, Transglutaminase cross-linking to enhance elastic properties of soy protein hydrogels with intercalated montmorillonite nanoclay. J. Food Eng. 115, 33–40 (2013)
S.J. Jiang, X.H. Zhao, Transglutaminase-induced cross-linking and glucosamine conjugation of casein and some functional properties of the modified product. Int. Dairy J. 21, 198–205 (2011)
P.K. Hong, D. Gottardi, M. Ndagijimana, M. Betti, Glycation and transglutaminase mediated glycosylation of fish gelatin peptides with glucosamine enhance bioactivity. Food Chem. 142, 285–293 (2014)
C.L. Song, X.H. Zhao, Structure and property modification of an oligochitosan-glycosylated and crosslinked soybean protein generated by microbial transglutaminase. Food Chem. 163, 114–119 (2014)
J. Adler-Nissen, Limited enzymic degradation of proteins: a new approach in the industrial application of hydrolases. J. Chem. Tech. Biotechnol. 32, 138–156 (1982)
Y. Xia, F. Bamdad, M. Gänzle, L. Chen, Fractionation and characterization of antioxidant peptides derived from barley glutelin by enzymatic hydrolysis. Food Chem. 134, 1509–1518 (2012)
R.R. Lu, P. Qian, Z. Sun, X.H. Zhou, T.P. Chen, J.F. He, H. Zhang, J. Wu, Hempseed protein derived antioxidative peptides: purification, identification and protection from hydrogen peroxide-induced apoptosis in PC12 cells. Food Chem. 123, 1210–1218 (2010)
B. Halliwell, Superoxide-dependent formation of hydroxyl radicals in the presence of iron chelates: is it a mechanism for hydroxyl radical production in biochemical systems? FEBS Lett. 92, 321–326 (1978)
K.N. Pearce, J.E. Kinsella, Emulsifying properties of proteins: evaluation of a turbidimetric technique. J. Agric. Food Chem. 26, 716–723 (1978)
K. Marciniak-Darmochwal, H. Kostyra, Influence of nonenzymatic glycosylation (glycation) of pea proteins (Pisum sativum) on their susceptibility to enzymatic hydrolysis. J. Food Biochem. 33, 506–521 (2009)
S.W. Yin, C.H. Tang, Q.B. Wen, X.Q. Yang, D.B. Yuan, The relationships between physicochemical properties and conformational features of succinylated and acetylated kidney bean (Phaseolus vulgaris L.) protein isolates. Food Res. Int. 43, 730–738 (2010)
C.Y. Zhu, H.F. Liu, M. Fu, X.H. Zhao, Structure and property changes of soybean protein isolates resulted from the glycation and cross-linking by transglutaminase and a degraded chitosan. CyTA-J. Food 14, 138–144 (2016)
Y.H. Yong, S. Yamaguchi, Y. Matsumura, Effects of enzymatic deamidation by protein-glutaminase on structure and functional properties of wheat gluten. J. Agric. Food Chem. 54, 6034–6040 (2006)
J. Ren, C.L. Song, P. Wang, S. Li, N.K. Kopparapu, X.Q. Zheng, Modification of structural and functional properties of sunflower 11S globulin hydrolysates. Czech J. Food Sci. 33, 474–478 (2015)
M.Y. Shon, T.H. Kim, N.J. Sung, Antioxidants and free radical scavenging activity of Phellinus baumii (Phellinus of Hymenochaetaceae) extracts. Food Chem. 82, 593–597 (2003)
P. Liu, M. Huang, S. Song, K. Hayat, X. Zhang, S. Xia, C. Jia, Sensory characteristics and antioxidant activities of Maillard reaction products from soy protein hydrolysates with different molecular weight distribution. Food Bioprocess Technol. 5, 1775–1789 (2012)
P.J. Park, W.K. Jung, K.S. Nam, F. Shahidi, S.K. Kim, Purification and characterization of antioxidative peptides from protein hydrolysate of lecithin-free egg yolk. J. Am. Oil Chem. Soc. 78, 651–656 (2001)
X.Q. Zheng, J.T. Wang, X.L. Liu, Y. Sun, Y.J. Zheng, X.J. Wang, Y. Liu, Effect of hydrolysis time on the physicochemical and functional properties of corn glutelin by Protamex hydrolysis. Food Chem. 172, 407–415 (2015)
K. Govindaraju, H. Srinivas, Studies on the effects of enzymatic hydrolysis on functional and physico-chemical properties of arachin. LWT 39, 54–62 (2006)
S. Severin, W.S. Xia, Enzymatic hydrolysis of whey proteins by two different proteases and their effect on the functional properties of resulting protein hydrolysates. J. Food Biochem. 30, 77–97 (2006)
A. Kato, Industrial applications of Maillard-type protein-polysaccharide conjugates. Food Sci. Technol. Res. 8, 193–199 (2002)
N. Diftis, C. Biliaderis, V. Kiosseoglou, Rheological properties and stability of model salad dressing emulsions prepared with a dry-heated soybean protein isolate-dextran mixture. Food Hydrocol. 19, 1025–1031 (2005)
H.G. Kristinsson, B.A. Rasco, Fish protein hydrolysates: production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 40, 43–81 (2000)
M.E. Thomas, J. Scher, S. Desobry-Banon, S. Desobry, Milk powders ageing: effect on physical and functional properties. Crit. Rev. Food Sci. Nutr. 44, 297–322 (2004)
S. Oh, G.L. Catignani, H.E. Swaisgood, Characteristics of an immobilized form of transglutaminase: a possible increase in substrate specificity by selective interaction with a protein spacer. J. Agric. Food Chem. 41, 1337–1342 (1993)
S. Roufik, S.F. Gauthier, S.L. Turgeon, In vitro digestibility of bioactive peptides derived from bovine β-lactoglobulin. Int. Dairy J. 16, 294–302 (2006)
C. Branca, S. Magazù, G. Maisano, F. Migliardo, P. Migliardo, G. Romeo, α, α-Trehalose/water solutions. 5. Hydration and viscosity in dilute and semidilute disaccharide solutions. J. Phys. Chem. B 105, 10140–10145 (2001)
C.E. Kotlar, A.G. Ponce, S.I. Roura, Improvement of functional and antimicrobial properties of brewery byproduct hydrolysed enzymatically. LWT 50, 378–385 (2013)
Acknowledgements
This study was funded by the National Natural Science Foundation of China (Project No. 31401639) and Youth Innovative Talent Program of Heilongjiang Province of China (UNPYSCT-2015094).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Rights and permissions
About this article

Cite this article
Song, CL., Ren, J., Chen, JP. et al. Effect of glycosylation and limited hydrolysis on structural and functional properties of soybean protein isolate. Food Measure 12, 2946–2954 (2018). https://doi.org/10.1007/s11694-018-9910-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11694-018-9910-5